Molecular Data Reveal Multiple Lineages in Piranhas of the Genus Pygocentrus (Teleostei, Characiformes)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Extraction, Amplification and Sequencing
2.3. Species Delimitation Analyses
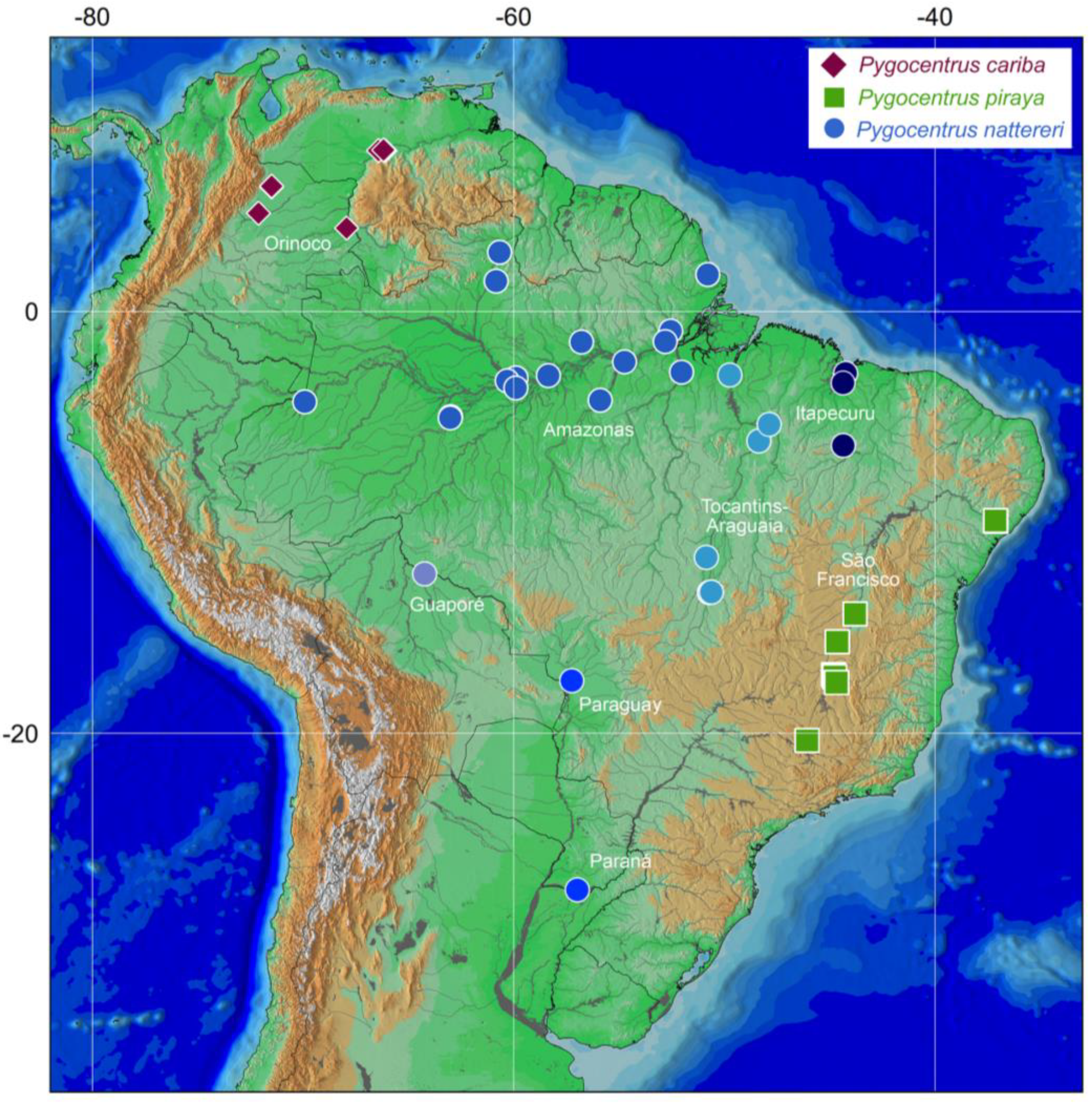
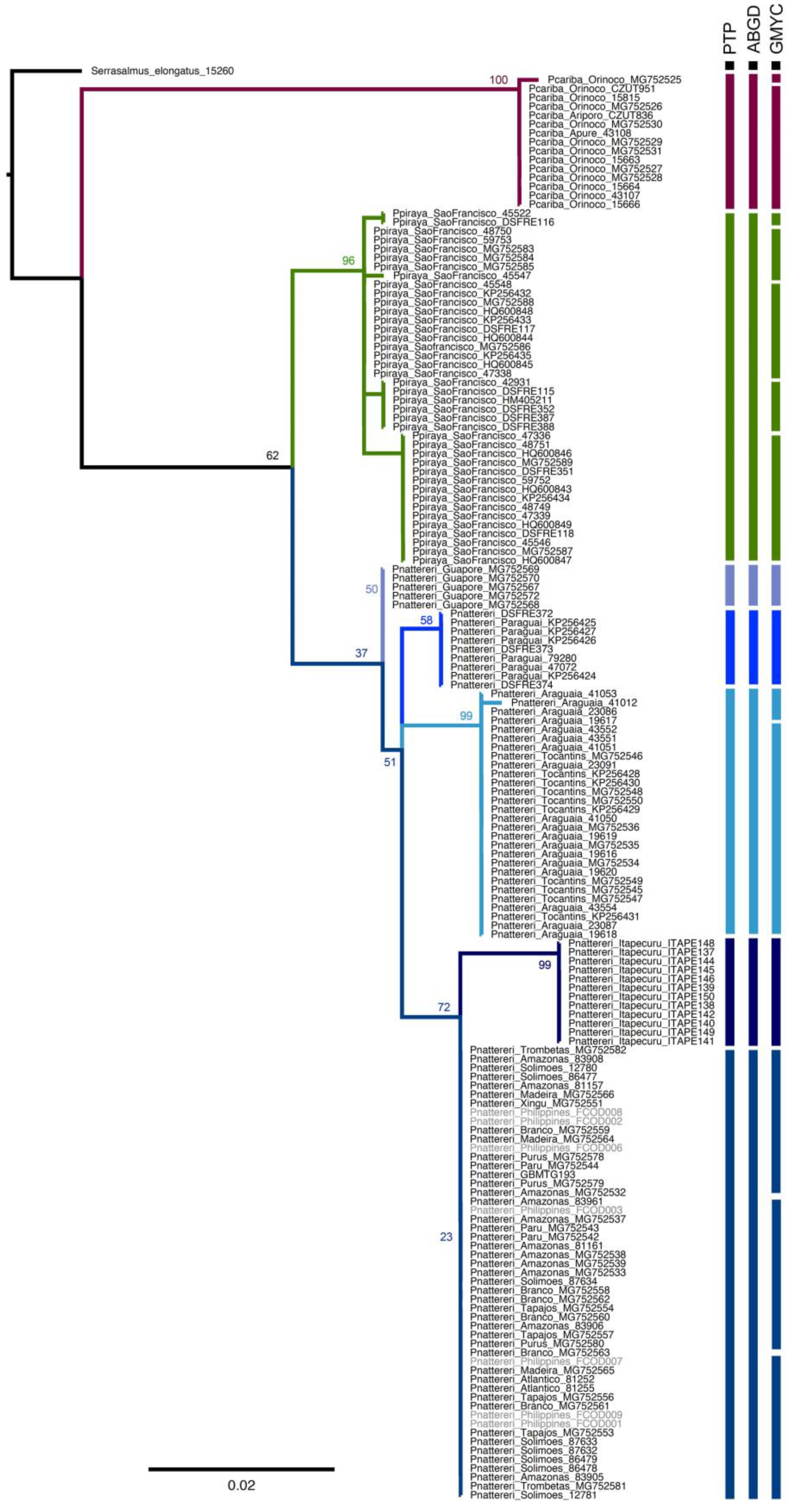
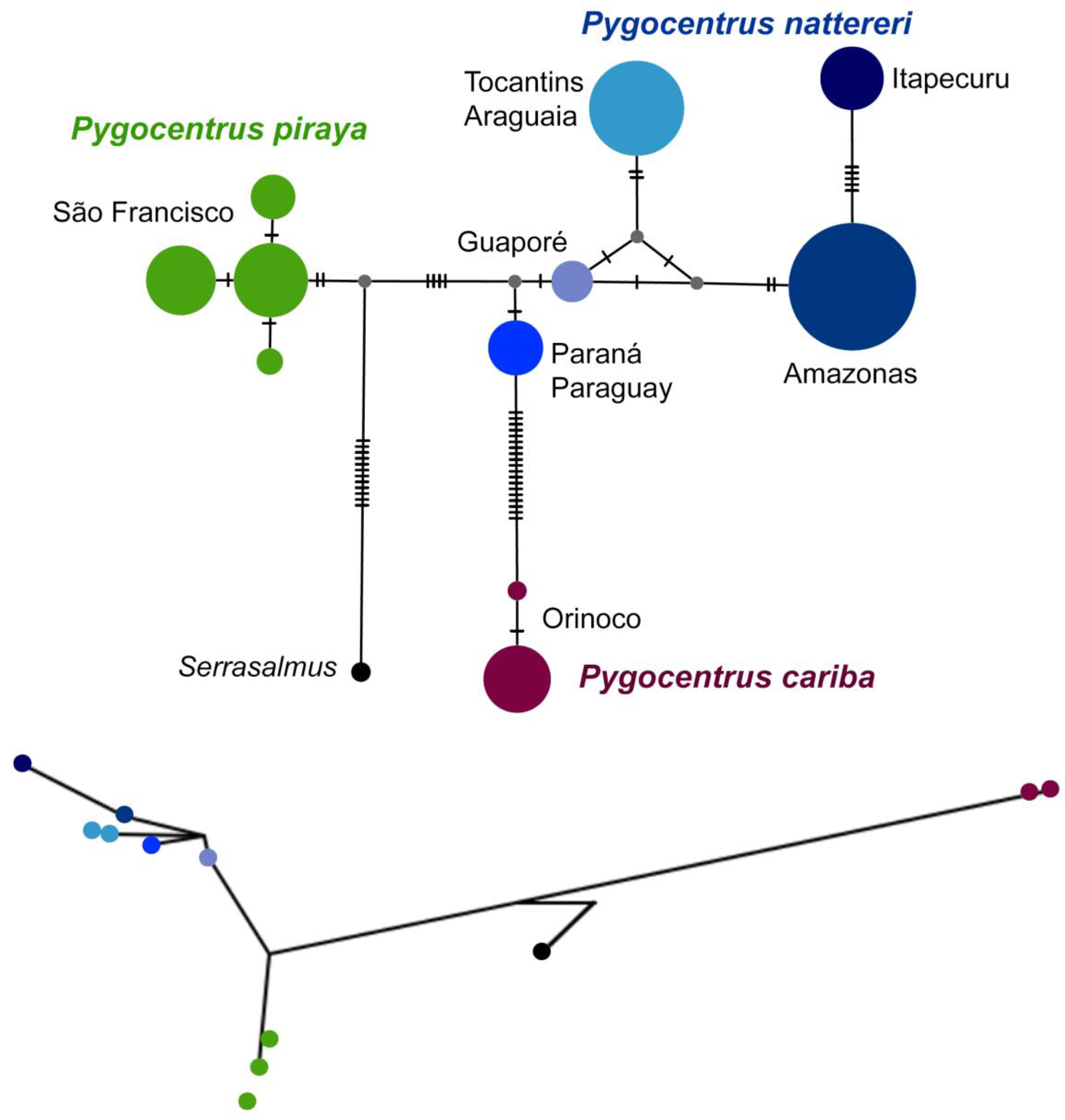
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Haplotype | Taxon | Voucher | Specimen | Locality, Basin | City, State | Country | Accession n. |
|---|---|---|---|---|---|---|---|
| 1 | P. cariba | LBP 2229 | 15663 | Río Orinoco basin | Caicara del Orinoco, Bolívar | Venezuela | PYGO003-18 |
| 1 | P. cariba | LBP 2229 | 15664 | Río Orinoco basin | Caicara del Orinoco, Bolívar | Venezuela | PYGO004-18 |
| 1 | P. cariba | LBP 2229 | 15666 | Río Orinoco basin | Caicara del Orinoco, Bolívar | Venezuela | PYGO005-18 |
| 1 | P. cariba | LBP 2290 | 15815 | Río Orinoco basin | Caicara del Orinoco, Bolívar | Venezuela | PYGO006-18 |
| 1 | P. cariba | LBP 10225 | 43107 | Río Apure, Orinoco basin | Cabruta, Guarico | Venezuela | PYGO001-18 |
| 1 | P. cariba | LBP 10225 | 43108 | Río Apure, Orinoco basin | Cabruta, Guarico | Venezuela | PYGO002-18 |
| 1 | P. cariba | CZUT-IC 12810 | 836 | Río Ariporo, Orinoco basin | - | Colombia | PYGO054-19 |
| 1 | P. cariba | CZUT-IC 11395 | 951 | Caño Materro, Orinoco basin | - | Colombia | PYGO052-19 |
| 2 | P. cariba | UFAM 13525 | 13525 | Río Orinoco basin | Guainía | Colombia | MG752525 |
| 1 | P. cariba | UFAM 13526 | 13526 | Río Orinoco basin | Guainía | Colombia | MG752526 |
| 1 | P. cariba | UFAM 13529 | 13529 | Río Orinoco basin | Guainía | Colombia | MG752527 |
| 1 | P. cariba | UFAM 13741 | 13741 | Río Orinoco basin | Guainía | Colombia | MG752528 |
| 1 | P. cariba | UFAM 13743 | 13743 | Río Orinoco basin | Guainía | Colombia | MG752529 |
| 1 | P. cariba | UFAM 13744 | 13744 | Río Orinoco basin | Guainía | Colombia | MG752530 |
| 1 | P. cariba | UFAM 13745 | 13745 | Río Orinoco basin | Guainía | Colombia | MG752531 |
| 6 | P. nattereri | INPA 41663 | 102036 | Rio Purus, Amazon basin | Amazonas | Brazil | MG752578 |
| 6 | P. nattereri | INPA 41689 | 102084 | Rio Purus, Amazon basin | Amazonas | Brazil | MG752579 |
| 6 | P. nattereri | INPA 41689 | 102085 | Rio Purus, Amazon basin | Amazonas | Brazil | MG752580 |
| 6 | P. nattereri | INPA 50418 | 105742 | Rio Trombetas, Amazon basin | Pará | Brazil | MG752581 |
| 6 | P. nattereri | INPA 50175 | 105817 | Rio Trombetas, Amazon basin | Pará | Brazil | MG752582 |
| 6 | P. nattereri | LBP 1697 | 12780 | Lago do Vanico, Amazon basin | Carero, Amazonas | Brazil | PYGO031-18 |
| 6 | P. nattereri | LBP 1697 | 12781 | Lago do Vanico, Amazon basin | Carero, Amazonas | Brazil | PYGO032-18 |
| 9 | P. nattereri | LBP 2978 | 19616 | Rio Araguaia, Amazon basin | Cocalinho, Mato Grosso | Brazil | PYGO014-18 |
| 9 | P. nattereri | LBP 2978 | 19617 | Rio Araguaia, Amazon basin | Cocalinho, Mato Grosso | Brazil | PYGO015-18 |
| 9 | P. nattereri | LBP 2978 | 19618 | Rio Araguaia, Amazon basin | Cocalinho, Mato Grosso | Brazil | PYGO016-18 |
| 9 | P. nattereri | LBP 2978 | 19619 | Rio Araguaia, Amazon basin | Cocalinho, Mato Grosso | Brazil | PYGO017-8 |
| 9 | P. nattereri | LBP 2978 | 19620 | Rio Araguaia, Amazon basin | Cocalinho, Mato Grosso | Brazil | PYGO018-18 |
| 9 | P. nattereri | LBP 4000 | 23086 | Rio Araguaia, Amazon basin | S. Félix Araguaia, Mato Grosso | Brazil | PYGO011-18 |
| 9 | P. nattereri | LBP 4000 | 23087 | Rio Araguaia, Amazon basin | S. Félix Araguaia, Mato Grosso | Brazil | PYGO012-18 |
| 9 | P. nattereri | LBP 4000 | 23091 | Rio Araguaia, Amazon basin | S. Félix Araguaia, Mato Grosso | Brazil | PYGO013-18 |
| 10 | P. nattereri | LBP 12641 | 47072 | Rio Cuiabá, Paraguay basin | Corumbá, Mato Grosso do Sul | Brazil | PYGO022-18 |
| 9 | P. nattereri | LBP 12693 | 43551 | Rio Araguaia, Amazon basin | Cocalinho, Mato Grosso | Brazil | PYGO019-18 |
| 9 | P. nattereri | LBP 12693 | 43552 | Rio Araguaia, Amazon basin | Cocalinho, Mato Grosso | Brazil | PYGO020-18 |
| 9 | P. nattereri | LBP 12693 | 43554 | Rio Araguaia, Amazon basin | Cocalinho, Mato Grosso | Brazil | PYGO021-18 |
| 9 | P. nattereri | LBP 12738 | 41012 | Rio Araguaia, Amazon basin | Cocalinho, Mato Grosso | Brazil | PYGO007-18 |
| 9 | P. nattereri | LBP 12738 | 41050 | Rio Araguaia, Amazon basin | Cocalinho, Mato Grosso | Brazil | PYGO008-18 |
| 9 | P. nattereri | LBP 12738 | 41051 | Rio Araguaia, Amazon basin | Cocalinho, Mato Grosso | Brazil | PYGO009-18 |
| 9 | P. nattereri | LBP 12738 | 41053 | Rio Araguaia, Amazon basin | Cocalinho, Mato Grosso | Brazil | PYGO010-18 |
| 10 | P. nattereri | LBP 19961 | 79280 | Rio Paraná, lower Paraná basin | Coratei | Paraguay | PYGO023-18 |
| 6 | P. nattereri | LBP 21836 | 83905 | Rio Negro, Amazon basin | Manaus, Amazonas | Brazil | PYGO024-18 |
| 6 | P. nattereri | LBP 21836 | 83906 | Rio Negro, Amazon basin | Manaus, Amazonas | Brazil | PYGO025-18 |
| 6 | P. nattereri | LBP 21836 | 83908 | Rio Negro, Amazon basin | Manaus, Amazonas | Brazil | PYGO026-18 |
| 6 | P. nattereri | LBP 21836 | 83961 | Rio Negro, Amazon basin | Manaus, Amazonas | Brazil | PYGO027-18 |
| 6 | P. nattereri | LBP 22328 | 86477 | Rio Solimões, Amazon basin | Tabatinga, Amazonas | Brazil | PYGO028-18 |
| 6 | P. nattereri | LBP 22328 | 86478 | Rio Solimões, Amazon basin | Tabatinga, Amazonas | Brazil | PYGO029-18 |
| 6 | P. nattereri | LBP 22328 | 86479 | Rio Solimões, Amazon basin | Tabatinga, Amazonas | Brazil | PYGO030-18 |
| 6 | P. nattereri | LBP 20651 | 81252 | Lago Pracuúba, Atlantic drainage | Pracuúba, Amapá | Brazil | PYGO033-18 |
| 6 | P. nattereri | LBP 20651 | 81255 | Lago Pracuúba, Atlantic drainage | Pracuúba, Amapá | Brazil | PYGO034-18 |
| 6 | P. nattereri | LBP 20977 | 81157 | Rio Jari, Amazon basin | Almeirim, Pará | Brazil | PYGO035-18 |
| 6 | P. nattereri | LBP 20977 | 81161 | Rio Jari, Amazon basin | Almeirim, Pará | Brazil | PYGO036-18 |
| 6 | P. nattereri | LBP 22815 | 87632 | Rio Solimões, Amazon basin | Iranduba, Amazonas | Brazil | PYGO051-19 |
| 6 | P. nattereri | LBP 22815 | 87633 | Rio Solimões, Amazon basin | Iranduba, Amazonas | Brazil | PYGO050-19 |
| 6 | P. nattereri | LBP 22815 | 87634 | Rio Solimões, Amazon basin | Iranduba, Amazonas | Brazil | PYGO049-19 |
| 10 | P. nattereri | NtrMS01 | - | Rio Paraguay basin | Unknown | Brazil | KP256424 |
| 10 | P. nattereri | NtrMS02 | - | Rio Paraguay basin | Unknown | Brazil | KP256425 |
| 10 | P. nattereri | NtrMS10 | - | Rio Paraguay basin | Unknown | Brazil | KP256426 |
| 10 | P. nattereri | NtrMS11 | - | Rio Paraguay basin | Unknown | Brazil | KP256427 |
| 9 | P. nattereri | NtrTO19 | - | Rio Tocantins basin | Unknown | Brazil | KP256428 |
| 9 | P. nattereri | NtrTO21 | - | Rio Tocantins basin | Unknown | Brazil | KP256429 |
| 9 | P. nattereri | NtrTO24 | - | Rio Tocantins basin | Unknown | Brazil | KP256430 |
| 9 | P. nattereri | NtrTO30 | - | Rio Tocantins basin | Unknown | Brazil | KP256431 |
| 10 | P. nattereri | OL-0485 | - | Unknown | Unknown | Unknown | DSFRE372-08 |
| 10 | P. nattereri | OL-0486 | - | Unknown | Unknown | Unknown | DSFRE373-08 |
| 10 | P. nattereri | OL-0487 | - | Unknown | Unknown | Unknown | DSFRE374-08 |
| 6 | P. nattereri | P1A | - | Introduced specimens | Metro Manila | Philippines | FCOD001-15 |
| 6 | P. nattereri | P1B | - | Introduced specimens | Metro Manila | Philippines | FCOD002-15 |
| 6 | P. nattereri | P1C | - | Introduced specimens | Metro Manila | Philippines | FCOD003-15 |
| 6 | P. nattereri | P2C | - | Introduced specimens | Metro Manila | Philippines | FCOD006-15 |
| 6 | P. nattereri | P3A | - | Introduced specimens | Metro Manila | Philippines | FCOD007-15 |
| 6 | P. nattereri | P3B | - | Introduced specimens | Metro Manila | Philippines | FCOD008-15 |
| 6 | P. nattereri | P3C | - | Introduced specimens | Metro Manila | Philippines | FCOD009-15 |
| 6 | P. nattereri | NC_015840 | - | Unknown | Unknown | Unknown | NC015840 |
| 8 | P. nattereri | UEMA 104549 | - | Rio Itapecuru basin | Rosário, Maranhão | Brazil | ITAPE137-15 |
| 8 | P. nattereri | UEMA 104549 | - | Rio Itapecuru basin | Rosário, Maranhão | Brazil | ITAPE138-15 |
| 8 | P. nattereri | UEMA 104549 | - | Rio Itapecuru basin | Rosário, Maranhão | Brazil | ITAPE139-15 |
| 8 | P. nattereri | UEMA 104549 | - | Rio Itapecuru basin | Rosário, Maranhão | Brazil | ITAPE140-15 |
| 8 | P. nattereri | UEMA 104549 | - | Rio Itapecuru basin | Rosário, Maranhão | Brazil | ITAPE141-15 |
| 8 | P. nattereri | UEMA 104549 | - | Rio Itapecuru basin | Rosário, Maranhão | Brazil | ITAPE142-15 |
| 8 | P. nattereri | UEMA 104549 | - | Rio Itapecuru basin | Itapecuru Mirim, Maranhão | Brazil | ITAPE144-15 |
| 8 | P. nattereri | UEMA 104549 | - | Rio Itapecuru basin | Itapecuru Mirim, Maranhão | Brazil | ITAPE145-15 |
| 8 | P. nattereri | UEMA 104549 | - | Rio Itapecuru basin | Itapecuru Mirim, Maranhão | Brazil | ITAPE146-15 |
| 8 | P. nattereri | UEMA 104550 | - | Rio Itapecuru basin | Mirador, Maranhão | Brazil | ITAPE148-15 |
| 8 | P. nattereri | UEMA 104550 | - | Rio Itapecuru basin | Mirador, Maranhão | Brazil | ITAPE149-15 |
| 8 | P. nattereri | UEMA 104550 | - | Rio Itapecuru basin | Mirador, Maranhão | Brazil | ITAPE150-15 |
| 6 | P. nattereri | UFAM 643 | 643 | Rio Amazonas basin | Pará | Brazil | MG752532 |
| 6 | P. nattereri | UFAM 644 | 644 | Rio Amazonas basin | Pará | Brazil | MG752533 |
| 9 | P. nattereri | UFAM 2931 | 2931 | Rio Araguaia basin | Pará | Brazil | MG752534 |
| 9 | P. nattereri | UFAM 2932 | 2932 | Rio Araguaia basin | Pará | Brazil | MG752535 |
| 9 | P. nattereri | UFAM 2933 | 2933 | Rio Araguaia basin | Pará | Brazil | MG752536 |
| 6 | P. nattereri | UFAM 3579 | 3579 | Rio Amazonas basin | Amazonas | Brazil | MG752537 |
| 6 | P. nattereri | UFAM 3580 | 3580 | Rio Amazonas basin | Amazonas | Brazil | MG752538 |
| 6 | P. nattereri | UFAM 3581 | 3581 | Rio Amazonas basin | Amazonas | Brazil | MG752539 |
| 6 | P. nattereri | UFAM 3829 | 3829 | Rio Paru, Amazon basin | Pará | Brazil | MG752542 |
| 6 | P. nattereri | UFAM 3830 | 3830 | Rio Paru, Amazon basin | Pará | Brazil | MG752543 |
| 6 | P. nattereri | UFAM 3831 | 3831 | Rio Paru, Amazon basin | Pará | Brazil | MG752544 |
| 9 | P. nattereri | UFAM 3903 | 3903 | Rio Tocantins basin | Pará | Brazil | MG752545 |
| 9 | P. nattereri | UFAM 3904 | 3904 | Rio Tocantins basin | Pará | Brazil | MG752546 |
| 9 | P. nattereri | UFAM 3905 | 3905 | Rio Tocantins basin | Pará | Brazil | MG752547 |
| 9 | P. nattereri | UFAM 4556 | 4556 | Rio Tocantins basin | Pará | Brazil | MG752548 |
| 9 | P. nattereri | UFAM 4557 | 4557 | Rio Tocantins basin | Pará | Brazil | MG752549 |
| 9 | P. nattereri | UFAM 4558 | 4558 | Rio Tocantins basin | Pará | Brazil | MG752550 |
| 6 | P. nattereri | UFAM 5553 | 5553 | Rio Xingu, Amazon basin | Pará | Brazil | MG752551 |
| 6 | P. nattereri | UFAM 11497 | 11497 | Rio Tapajós, Amazon basin | Pará | Brazil | MG752553 |
| 6 | P. nattereri | UFAM 11498 | 11498 | Rio Tapajós, Amazon basin | Pará | Brazil | MG752554 |
| 6 | P. nattereri | UFAM 11500 | 11500 | Rio Tapajós, Amazon basin | Pará | Brazil | MG752556 |
| 6 | P. nattereri | UFAM 11501 | 11501 | Rio Tapajós, Amazon basin | Pará | Brazil | MG752557 |
| 6 | P. nattereri | UFAM 12603 | 12603 | Rio Branco, Amazon basin | Roraima | Brazil | MG752558 |
| 6 | P. nattereri | UFAM 12626 | 12626 | Rio Branco, Amazon basin | Roraima | Brazil | MG752559 |
| 6 | P. nattereri | UFAM 14105 | 14105 | Rio Branco, Amazon basin | Roraima | Brazil | MG752560 |
| 6 | P. nattereri | UFAM 14106 | 14106 | Rio Branco, Amazon basin | Roraima | Brazil | MG752561 |
| 6 | P. nattereri | UFAM 14107 | 14107 | Rio Branco, Amazon basin | Roraima | Brazil | MG752562 |
| 6 | P. nattereri | UFAM 14108 | 14108 | Rio Branco, Amazon basin | Roraima | Brazil | MG752563 |
| 6 | P. nattereri | UFAM 15261 | 15261 | Rio Madeira, Amazon basin | Amazonas | Brazil | MG752564 |
| 6 | P. nattereri | UFAM 15262 | 15262 | Rio Madeira, Amazon basin | Amazonas | Brazil | MG752565 |
| 6 | P. nattereri | UFAM 15263 | 15263 | Rio Madeira, Amazon basin | Amazonas | Brazil | MG752566 |
| 11 | P. nattereri | UFAM 15272 | 15272 | Rio Guaporé, Amazon basin | Rondônia | Brazil | MG752567 |
| 11 | P. nattereri | UFAM 15273 | 15273 | Rio Guaporé, Amazon basin | Rondônia | Brazil | MG752568 |
| 11 | P. nattereri | UFAM 15274 | 15274 | Rio Guaporé, Amazon basin | Rondônia | Brazil | MG752569 |
| 11 | P. nattereri | UFAM 15276 | 15276 | Rio Guaporé, Amazon basin | Rondônia | Brazil | MG752570 |
| 11 | P. nattereri | UFAM 15278 | 15278 | Rio Guaporé, Amazon basin | Rondônia | Brazil | MG752572 |
| 4 | P. piraya | INPA 56741 | 15283 | Rio São Francisco basin | Minas Gerais/Bahia | Brazil | MG752583 |
| 4 | P. piraya | INPA 56741 | 15284 | Rio São Francisco basin | Minas Gerais/Bahia | Brazil | MG752584 |
| 4 | P. piraya | INPA 56741 | 15285 | Rio São Francisco basin | Minas Gerais/Bahia | Brazil | MG752585 |
| 3 | P. piraya | LBP 11286 | 48749 | Rio São Francisco basin | Gararu, Sergipe | Brazil | PYGO037-18 |
| 4 | P. piraya | LBP 11286 | 48750 | Rio São Francisco basin | Gararu, Sergipe | Brazil | PYGO038-18 |
| 3 | P. piraya | LBP 11286 | 48751 | Rio São Francisco basin | Gararu, Sergipe | Brazil | PYGO039-18 |
| 5 | P. piraya | LBP 11300 | 42931 | Rio São Francisco basin | S. Roque Minas, Minas Gerais | Brazil | PYGO046-18 |
| 7 | P. piraya | LBP 11336 | 45522 | Rio São Francisco basin | S. Roque Minas, Minas Gerais | Brazil | PYGO047-18 |
| 3 | P. piraya | LBP 11337 | 45546 | Rio São Francisco basin | Pirapora, Minas Gerais | Brazil | PYGO055-19 |
| 4 | P. piraya | LBP 11337 | 45547 | Rio São Francisco basin | Pirapora, Minas Gerais | Brazil | PYGO057-19 |
| 4 | P. piraya | LBP 11337 | 45548 | Rio São Francisco basin | Pirapora, Minas Gerais | Brazil | PYGO056-19 |
| 3 | P. piraya | LBP 21613 | 47336 | Rio São Francisco basin | Pirapora, Minas Gerais | Brazil | PYGO040-18 |
| 3 | P. piraya | LBP 21613 | 47337 | Rio São Francisco basin | Pirapora, Minas Gerais | Brazil | PYGO041-18 |
| 4 | P. piraya | LBP 21613 | 47338 | Rio São Francisco basin | Pirapora, Minas Gerais | Brazil | PYGO042-18 |
| 3 | P. piraya | LBP 21613 | 47339 | Rio São Francisco basin | Pirapora, Minas Gerais | Brazil | PYGO043-18 |
| 3 | P. piraya | LBP 21612 | 59752 | Rio São Francisco basin | Jenipatuba, Minas Gerais | Brazil | PYGO044-18 |
| 4 | P. piraya | LBP 21612 | 59753 | Rio São Francisco basin | Jenipatuba, Minas Gerais | Brazil | PYGO045-18 |
| 3 | P. piraya | DCC502 | - | Rio Pandeiros, São Francisco basin | Januária, Minas Gerais | Brazil | HQ600843 |
| 4 | P. piraya | DCC503 | - | Rio Pandeiros, São Francisco basin | Januária, Minas Gerais | Brazil | HQ600844 |
| 4 | P. piraya | DCC499 | - | Rio Pandeiros, São Francisco basin | Januária, Minas Gerais | Brazil | HQ600845 |
| 3 | P. piraya | DCC501 | - | Rio Pandeiros, São Francisco basin | Januária, Minas Gerais | Brazil | HQ600846 |
| 3 | P. piraya | DCC532 | - | Rio Pandeiros, São Francisco basin | Januária, Minas Gerais | Brazil | HQ600847 |
| 4 | P. piraya | DCC500 | - | Rio Pandeiros, São Francisco basin | Januária, Minas Gerais | Brazil | HQ600848 |
| 3 | P. piraya | DCC306 | - | Rio Pandeiros, São Francisco basin | Januária, Minas Gerais | Brazil | HQ600849 |
| 5 | P. piraya | DCC1043 | - | Rio São Francisco basin | Várzea da Palma, Minas Gerais | Brazil | HM405211 |
| 4 | P. piraya | PrySF499 | - | Rio São Francisco basin | Unknown | Brazil | KP256432 |
| 4 | P. piraya | PrySF500 | - | Rio São Francisco basin | Unknown | Brazil | KP256433 |
| 3 | P. piraya | PrySF501 | - | Rio São Francisco basin | Unknown | Brazil | KP256434 |
| 4 | P. piraya | PrySF503 | - | Rio São Francisco basin | Unknown | Brazil | KP256435 |
| 5 | P. piraya | - | - | Unknown | Unknown | Unknown | DSFRE115-08 |
| 7 | P. piraya | - | - | Unknown | Unknown | Unknown | DSFRE116-08 |
| 4 | P. piraya | - | - | Unknown | Unknown | Unknown | DSFRE117-08 |
| 3 | P. piraya | - | - | Unknown | Unknown | Unknown | DSFRE118-08 |
| 3 | P. piraya | - | - | Unknown | Unknown | Unknown | DSFRE351-08 |
| 5 | P. piraya | - | - | Unknown | Unknown | Unknown | DSFRE352-08 |
| 5 | P. piraya | - | - | Unknown | Unknown | Unknown | DSFRE387-08 |
| 5 | P. piraya | - | - | Unknown | Unknown | Unknown | DSFRE388-08 |
| 4 | P. piraya | UFAM 15286 | 15286 | Rio São Francisco basin | Minas Gerais/Bahia | Brazil | MG752586 |
| 3 | P. piraya | UFAM 15287 | 15287 | Rio São Francisco basin | Minas Gerais/Bahia | Brazil | MG752587 |
| 4 | P. piraya | UFAM 15288 | 15288 | Rio São Francisco basin | Minas Gerais/Bahia | Brazil | MG752588 |
| 3 | P. piraya | UFAM 15289 | 15289 | Rio São Francisco basin | Minas Gerais/Bahia | Brazil | MG752589 |
| 12 | S. elongatus | UFAM 15260 | 15260 | Rio Madeira basin | Amazonas | Brazil | MG752622 |
References
- CAS-Eschmeyer’s Catalog of Fishes-Species by Family. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 1 April 2019).
- Thompson, A.W.; Betancur-R, R.; López-Fernández, H.; Ortí, G. A time-calibrated, multi-locus phylogeny of piranhas and pacus (Characiformes: Serrasalmidae) and a comparison of species tree methods. Mol. Phylogenet. Evol. 2014, 81, 242–257. [Google Scholar] [CrossRef]
- Machado-Allison, A. Estudios sobre la sistemática de la subfamilia Serrasalminae (Teleostei, Characidae). Parte II. Discusión sobre la condición monofilética de la subfamilia. Acta Biol. Venez. 1983, 11, 145–195. [Google Scholar]
- Cione, A.L.; Dahdul, W.M.; Lundberg, J.G.; Machado-Allison, A. Megapiranha paranensis, a new genus and species of Serrasalmidae (Characiformes, Teleostei) from the upper Miocene of Argentina. J. Vertebr. Paleontol. 2007, 29, 350–358. [Google Scholar] [CrossRef]
- Ortí, G.; Petry, P.; Porto, J.I.R.; Jégu, M.; Meyer, A. Patterns of nucleotide change in mitochondrial ribosomal RNA genes and the phylogeny of piranhas. J. Mol. Evol. 1996, 42, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Ortí, G.; Sivasundar, A.; Dietz, K.; Jégu, M. Phylogeny of the Serrasalmidae (Characiformes) based on mitochondrial DNA sequences. Genet. Mol. Biol. 2008, 31, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Britski, H.A.; Silimon, K.Z.S.; Lopes, B.S. Peixes do Pantanal-Manual de identificação, 2nd ed.; Embrapa: Brasília, Brazil, 2007. [Google Scholar]
- Santos, G.; Ferreira, E.; Zuanon, J. Peixes comerciais de Manaus; Ibama/AM, ProVárzea: Manaus, Brazil, 2006.
- Lin, Y.; Gao, Z.; Zhan, A. Introduction and use of non-native species for aquaculture in China: Status, risks and management solutions. Rev. Aquac. 2015, 7, 28–58. [Google Scholar] [CrossRef]
- Fink, W.L. Revision of the Piranha Genus Pygocentrus (Teleostei, Characiformes). Copeia 1993, 1993, 665. [Google Scholar] [CrossRef]
- Nico, L.G.; Jégu, M.; Andrade, M.C. Family Serrasalmidae-Piranhas and Pacus. In Field guide to the fishes of the Amazon, Orinoco, and Guianas; Van der Sleen, P., Albert, J., Eds.; D&N Publishing: Wiltshire, UK, 2017; pp. 172–196. [Google Scholar]
- Machado-Allison, A.; Fink, W.L. Notas sobre la sistemática del género Serrasalmus y géneros relacionados. Parte II: el género Pygocentrus en Venezuela. Acta Biol. Venez. 1991, 13, 109–135. [Google Scholar]
- Hubert, N.; Duponchelle, F.; Nuñez, J.; Garcia-Davila, C.; Paugy, D.; Renno, J.-F. Phylogeography of the piranha genera Serrasalmus and Pygocentrus: Implications for the diversification of the Neotropical ichthyofauna. Mol. Ecol. 2007, 16, 2115–2136. [Google Scholar] [CrossRef]
- Fink, W.L.; Zelditch, M.L. Shape Analysis and Taxonomic Status of Pygocentrus Piranhas (Ostariophysi, Characiformes) from the Paraguay and Paraná River Basins of South America. Copeia 1997, 1997, 179–182. [Google Scholar] [CrossRef]
- Machado, V.N.; Collins, R.A.; Ota, R.P.; Andrade, M.C.; Farias, I.P.; Hrbek, T. One thousand DNA barcodes of piranhas and pacus reveal geographic structure and unrecognised diversity in the Amazon. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Luz, L.A.; Reis, L.L.; Sampaio, I.; Barros, M.C.; Fraga, E. Genetic differentiation in the populations of red piranha, Pygocentrus nattereri Kner (1860) (Characiformes: Serrasalminae), from the river basins of northeastern Brazil. Braz. J. Biol. 2015, 75, 838–845. [Google Scholar] [CrossRef]
- Dos Santos, C.H.; de Sá Leitão, C.S.; Paula-Silva, M.D.N.; Almeida-Val, V.M.F. Genetic differentiation in red-bellied piranha populations (Pygocentrus nattereri, Kner, 1858) from the Solimões-Amazonas River. Ecol. Evol. 2016, 6, 4203–4213. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Ahmed, A.T.A. Studies on breeding and larval development of red bellied piranha, Pygocentrus nattereri kner, 1858 in bangladesh. Bangladesh J. Zool. 2007, 35, 193–203. [Google Scholar]
- Guerrero, R.I. Impacts of Introduced Freshwater Fishes in the Philippines (1905–2013): A Review and Recommendations. Philipp. J. Sci. 2014, 143, 49–59. [Google Scholar]
- Sarmiento, K.P.; Ventolero, M.F.H.; Santos, M.D. Identification of prohibited red piranha, Pygocentrus nattereri (Kner 1858) from confiscated juvenile fish in Manila, Philippines. Int. J. Philipp. Sci. Technol. 2016, 9, 7. [Google Scholar]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA barcoding Australia’s fish species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Xia, X. DAMBE7: New and Improved Tools for Data Analysis in Molecular Biology and Evolution. Mol. Biol. Evol. 2018, 35, 1550–1552. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. Partitionfinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinform. Oxf. Engl. 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Pattengale, N.D.; Alipour, M.; Bininda-Emonds, O.R.P.; Moret, B.M.E.; Stamatakis, A. How Many Bootstrap Replicates Are Necessary? In Proceedings of the Research in Computational Molecular Biology; Batzoglou, S., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 184–200. [Google Scholar]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinform. Oxf. Engl. 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, T.; Barraclough, T.G. Delimiting Species Using Single-Locus Data and the Generalized Mixed Yule Coalescent Approach: A Revised Method and Evaluation on Simulated Data Sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Mamlouk, A.M.; Martinetz, T.; Chang, S.; Wang, J.; Hilgenfeld, R. PhyloMap: An algorithm for visualizing relationships of large sequence data sets and its application to the influenza A virus genome. BMC Bioinform. 2011, 12, 248. [Google Scholar] [CrossRef] [PubMed]
- Géry, J. Poissons characoïdes des Guyanes. I. Géneralités. II. Famille des Serrasalmidae. Zool. Verh. Leiden 1972, 122, 1–250. [Google Scholar]
- Mayr, E. Animal species and their evolution; Harvard University Press: Cambridge, MA, USA; Oxford University Press: Oxford, UK, 1963. [Google Scholar]
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.L.; Meier, R.; Winker, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 2007, 22, 148–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaral, C.R.L.; Brito, P.M.; Silva, D.A.; Carvalho, E.F. A New Cryptic Species of South American Freshwater Pufferfish of the Genus Colomesus (Tetraodontidae), Based on Both Morphology and DNA Data. PloS ONE 2013, 8, e74397. [Google Scholar] [CrossRef] [PubMed]
- Costa-Silva, G.J.; Rodriguez, M.S.; Roxo, F.F.; Foresti, F.; Oliveira, C. Using Different Methods to Access the Difficult Task of Delimiting Species in a Complex Neotropical Hyperdiverse Group. PloS ONE 2015, 10, e0135075. [Google Scholar] [CrossRef]
- Cryptic species in the Neotropical fish genus Curimatopsis (Teleostei, Characiformes)-Melo-2016-Zoologica Scripta-Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/zsc.12178 (accessed on 9 May 2019).
- Latini, A.O.; Petrere, M. Reduction of a native fish fauna by alien species: An example from Brazilian freshwater tropical lakes. Fish. Manag. Ecol. 2004, 11, 71–79. [Google Scholar] [CrossRef]
- Trindade, M.E.d.J.; Jucá-Chagas, R. Diet of two serrasalmin species, Pygocentrus piraya and Serrasalmus brandtii (Teleostei: Characidae), along a stretch of the rio de Contas, Bahia, Brazil. Neotrop. Ichthyol. 2008, 6, 645–650. [Google Scholar] [CrossRef]
- Simberloff, D. Biological invasions: What’s worth fighting and what can be won? Ecol. Eng. 2014, 65, 112–121. [Google Scholar] [CrossRef]
- Mateussi, N.T.B.; Pavanelli, C.S.; Oliveira, C. Molecular identification of cryptic diversity in species of cis-Andean Mylossoma (Characiformes: Serrasalmidae). Mitochondrial DNA Part DNA Mapp. Seq. Anal. 2017, 28, 778–780. [Google Scholar] [CrossRef]
- Mateussi, N.T.B.; Oliveira, C.; Pavanelli, C.S. Taxonomic revision of the Cis-Andean species of Mylossoma Eigenmann & Kennedy, 1903 (Teleostei: Characiformes: Serrasalmidae). Zootaxa 2018, 4387, 275–309. [Google Scholar] [PubMed]
- Andrade, M.C.; Machado, V.N.; Jégu, M.; Farias, I.P.; Giarrizzo, T. A new species of Tometes Valenciennes 1850 (Characiformes: Serrasalmidae) from Tocantins-Araguaia River Basin based on integrative analysis of molecular and morphological data. PloS ONE 2017, 12, e0170053. [Google Scholar] [CrossRef] [PubMed]




| P. cariba | P. piraya | P. nattereri Amaz | P. nattereri Itapec | P. nattereri Toc/Ara | P. nattereri Par/Par | P. nattereri Guap | S. elongatus | |
|---|---|---|---|---|---|---|---|---|
| P. cariba | 0.000 ± 0.000 | |||||||
| P. piraya | 0.059 ± 0.010 | 0.003 ± 0.001 | ||||||
| P. nattereri Amaz | 0.051 ± 0.010 | 0.025 ± 0.006 | 0.000 ± 0.000 | |||||
| P. nattereri Itapec | 0.056 ± 0.010 | 0.035 ± 0.008 | 0.009 ± 0.004 | 0.000 ± 0.000 | ||||
| P. nattereri Toc/Ara | 0.056 ± 0.010 | 0.027 ± 0.007 | 0.011 ± 0.004 | 0.017 ± 0.005 | 0.000 ± 0.000 | |||
| P. nattereri Par/Par | 0.045 ± 0.009 | 0.019 ± 0.005 | 0.009 ± 0.004 | 0.015 ± 0.005 | 0.011 ± 0.004 | 0.000 ± 0.000 | ||
| P. nattereri Guap | 0.050 ± 0.009 | 0.017 ± 0.005 | 0.007 ± 0.003 | 0.017 ± 0.005 | 0.009 ± 0.039 | 0.005 ± 0.003 | 0.000 ± 0.000 | |
| S. elongatus | 0.046 ± 0.009 | 0.037 ± 0.008 | 0.041 ± 0.009 | 0.048 ± 0.009 | 0.042 ± 0.009 | 0.037 ± 0.008 | 0.040 ± 0.008 | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mateussi, N.T.B.; Melo, B.F.; Foresti, F.; Oliveira, C. Molecular Data Reveal Multiple Lineages in Piranhas of the Genus Pygocentrus (Teleostei, Characiformes). Genes 2019, 10, 371. https://doi.org/10.3390/genes10050371
Mateussi NTB, Melo BF, Foresti F, Oliveira C. Molecular Data Reveal Multiple Lineages in Piranhas of the Genus Pygocentrus (Teleostei, Characiformes). Genes. 2019; 10(5):371. https://doi.org/10.3390/genes10050371
Chicago/Turabian StyleMateussi, Nadayca T.B., Bruno F. Melo, Fausto Foresti, and Claudio Oliveira. 2019. "Molecular Data Reveal Multiple Lineages in Piranhas of the Genus Pygocentrus (Teleostei, Characiformes)" Genes 10, no. 5: 371. https://doi.org/10.3390/genes10050371
APA StyleMateussi, N. T. B., Melo, B. F., Foresti, F., & Oliveira, C. (2019). Molecular Data Reveal Multiple Lineages in Piranhas of the Genus Pygocentrus (Teleostei, Characiformes). Genes, 10(5), 371. https://doi.org/10.3390/genes10050371