Variable Number Tandem Repeats in the Mitochondrial DNA of Lentinula edodes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Mating between Dikaryons and Monokaryons
2.3. Analysis of Mitochondrial DNA Sequences
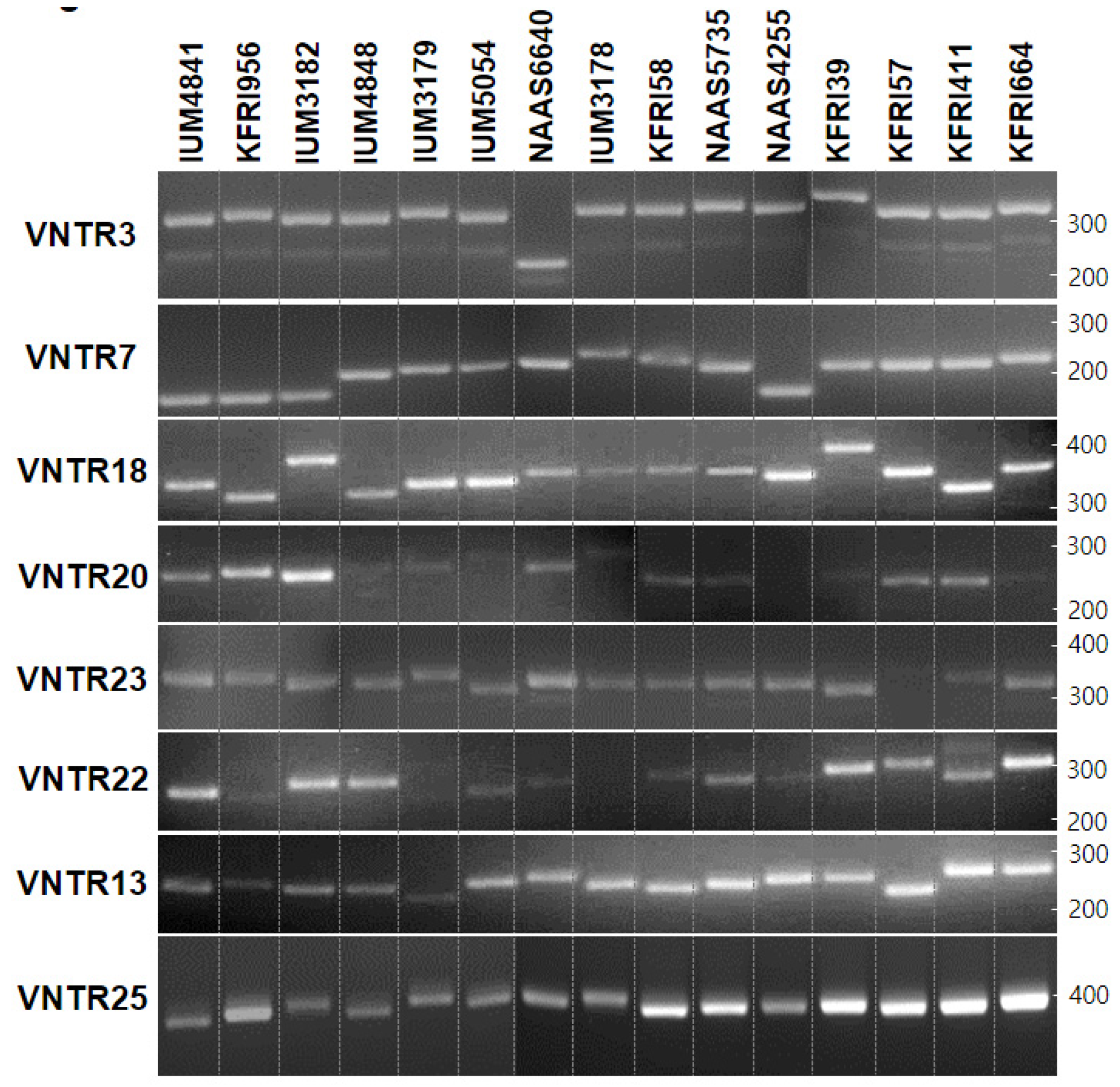
2.4. PCR Analyses Targeting VNTRs
3. Results
3.1. VNTRs in the Mitochondrial DNA
3.2. VNTRs in the Verification of Mitochondria in Lentinula edodes
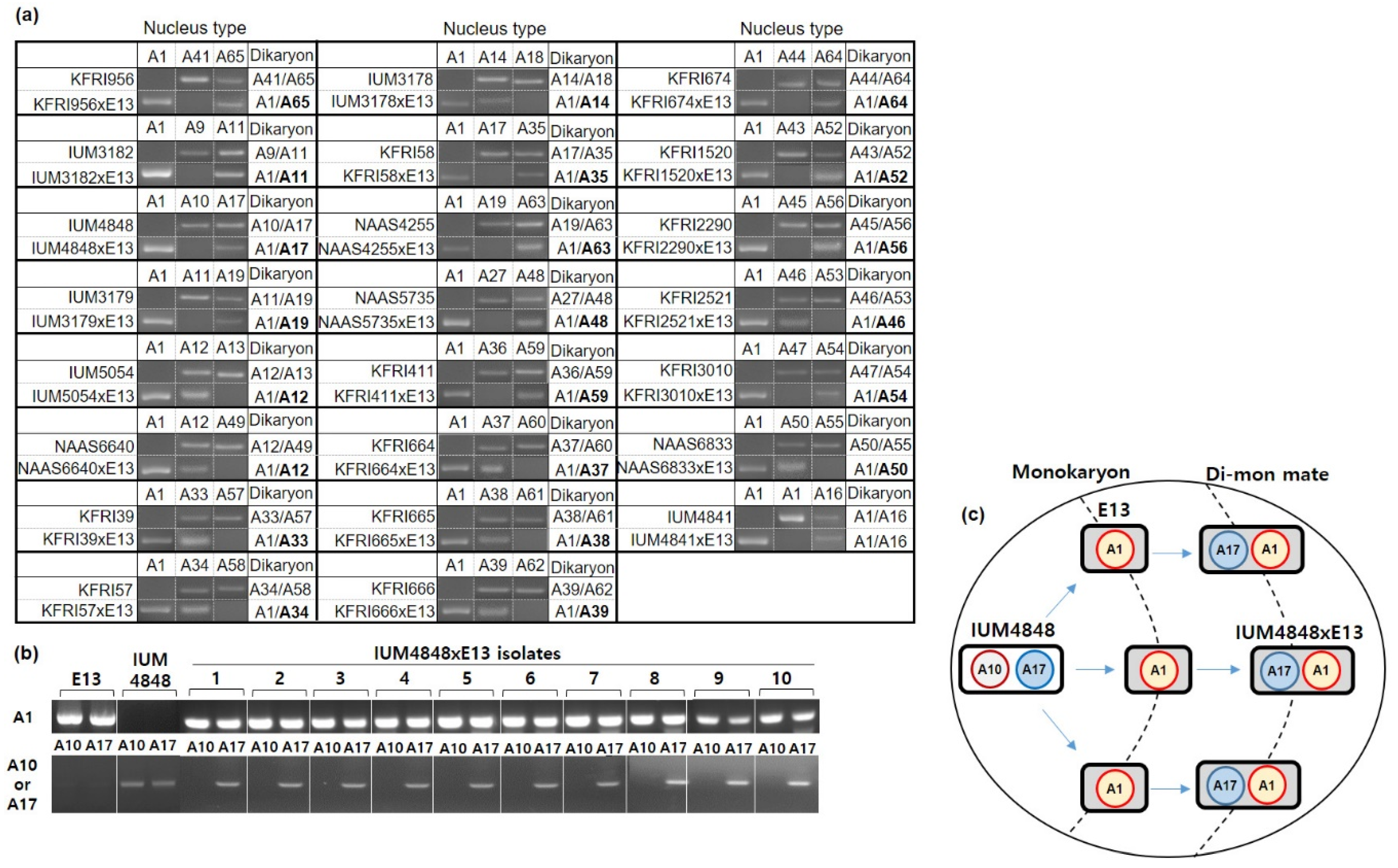
3.3. Unidirectional Movement of the Nucleus in Dikaryon–Monokaryon Mating
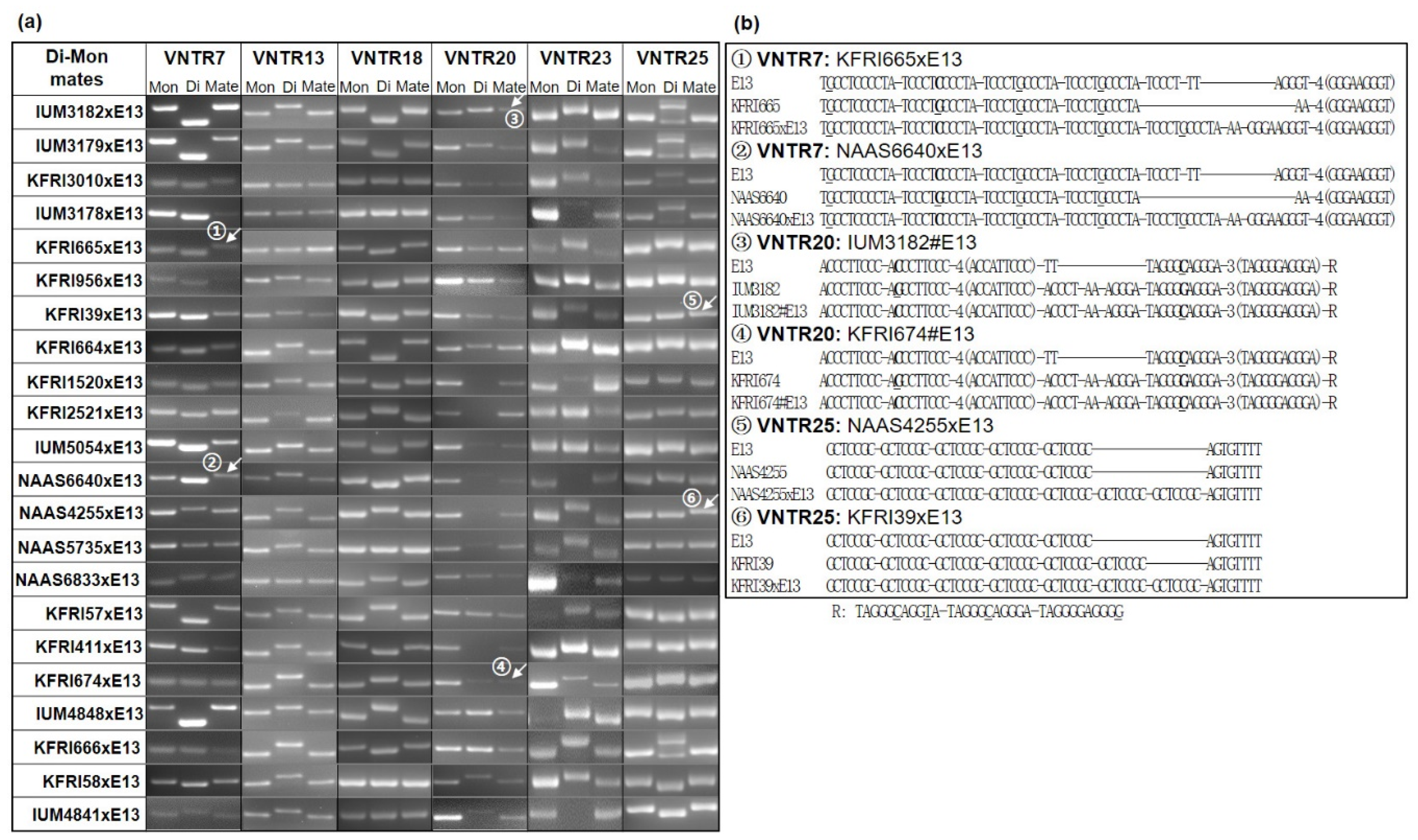
3.4. Mitochondrial Inheritance as a Marker of Cytoplasm Selection in Di–Mon Mating
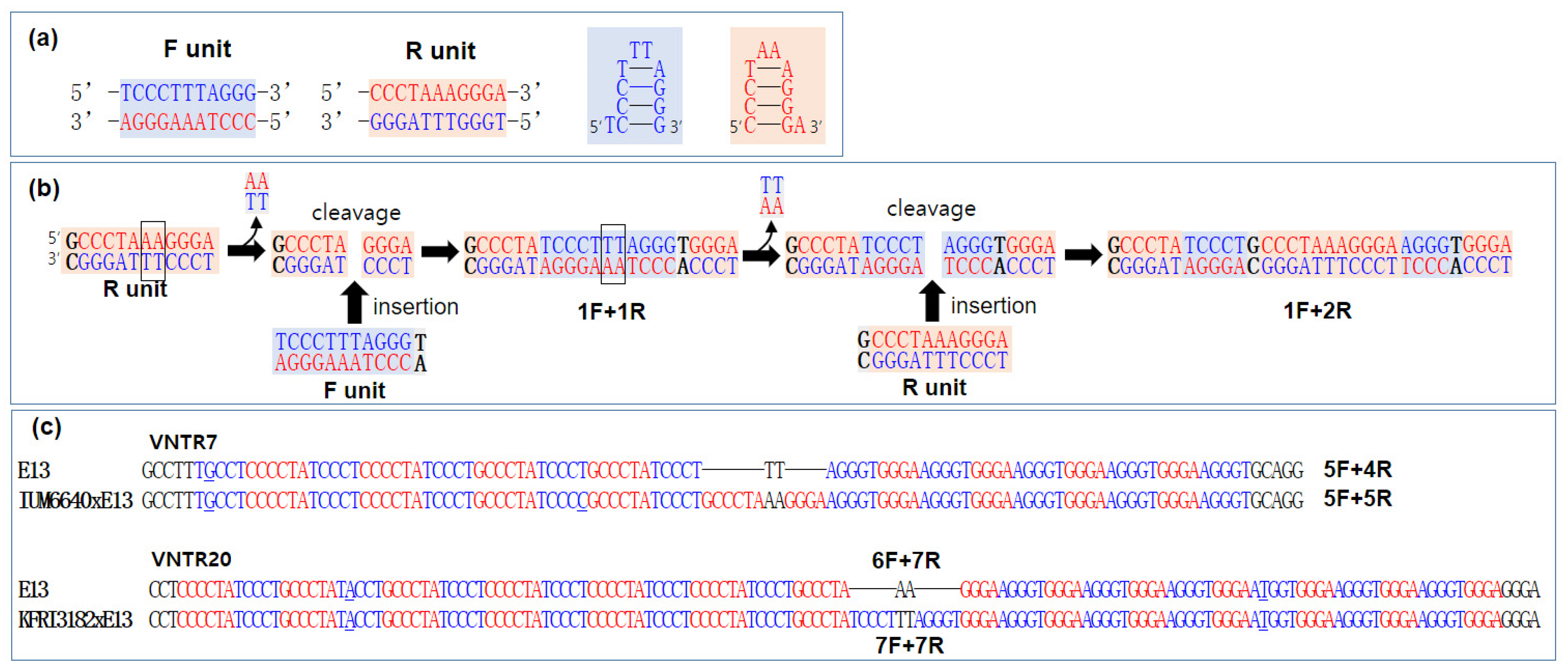
3.5. Elongation of Repeat Units in VNTRs and Implications for mtDNA Evolution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chan, D.C. Mitochondria: Dynamic organelles in disease, aging, and development. Cell 2006, 125, 1241–1252. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Sánchez, R.; Rodríguez-Enríquez, S.; Marín-Hernández, A.; Saavedra, E. Energy metabolism in tumor cells. FEBS J. 2007, 274, 1393–1418. [Google Scholar] [CrossRef] [PubMed]
- Pattappa, G.; Heywood, H.K.; de Bruijn, J.D.; Lee, D.A. The metabolism of human mesenchymal stem cells during proliferation and differentiation. J. Cell. Physiol. 2011, 226, 2562–2570. [Google Scholar] [CrossRef] [PubMed]
- Sweet, S.; Singh, G. Changes in mitochondrial mass, membrane potential, and cellular adenosine triphosphate content during the cell cycle of human leukemic (HL-60) cells. J. Cell. Physiol. 1999, 180, 91–96. [Google Scholar] [CrossRef]
- Sullivan, L.B.; Gui, D.Y.; Hosios, A.M.; Bush, L.N.; Freinkman, E.; Vander Heiden, M.G. Supporting aspartate biosynthesis is an essential function of respiration in proliferating cells. Cell 2015, 162, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Diebold, L.P.; Gil, H.J.; Gao, P.; Martinez, C.A.; Weinberg, S.E.; Chandel, N.S. Mitochondrial complex III is necessary for endothelial cell proliferation during angiogenesis. Nat. Metab. 2019, 1, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Andersson, S.G.; Karlberg, O.; Canback, B.; Kurland, C.G. On the origin of mitochondria: A genomics perspective. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2003, 358, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Giles, R.E.; Blanc, H.; Cann, H.M.; Wallace, D.C. Maternal inheritance of human mitochondrial DNA. Proc. Nat. Acad. Sci. USA 1980, 77, 6715–6719. [Google Scholar] [CrossRef] [PubMed]
- Hauswirth, W.W.; Laipis, P.J. Mitochondrial DNA polymorphism in a maternal lineage of Holstein cows. Proc. Nat. Acad. Sci. USA 1982, 79, 4686–4690. [Google Scholar] [CrossRef]
- Sato, M.; Sato, K. Maternal inheritance of mitochondrial DNA: degradation of paternal mitochondria by allogeneic organelle autophagy, allophagy. Autophagy 2012, 8, 424–425. [Google Scholar] [CrossRef]
- Mogensen, H.L. The hows and whys of cytoplasmic inheritance in seed plants. Am. J. Bot. 1996, 83, 383–404. [Google Scholar] [CrossRef]
- Nunnari, J.; Marshall, W.F.; Straight, A.; Murray, A.; Sedat, J.W.; Walter, P. Mitochondrial transmission during mating in Saccharomyces cerevisiae is determined by mitochondrial fusion and fission and the intramitochondrial segregation of mitochondrial DNA. Mol. Biol. Cell 1997, 8, 1233–1242. [Google Scholar] [CrossRef] [PubMed]
- Berger, K.H.; Yaffe, M.P. Mitochondrial DNA inheritance in Saccharomyces cerevisiae. Trends Microbiol. 2000, 8, 508–513. [Google Scholar] [CrossRef]
- Dujon, B.; Slonimski, P.P.; Weill, L. Mitochondrial genetics IX: A model for recombination and segregation of mitochondrial genomes in Saccharomyces cerevisiae. Genetics 1974, 78, 415–437. [Google Scholar]
- Xu, J.; Ali, R.Y.; Gregory, D.A.; Amick, D.; Lambert, S.E.; Yoell, H.J.; Vigalys, R.J.; Mitchell, T.G. Uniparental mitochondrial transmission in sexual crosses in Cryptococcus neoformans. Curr. Microbiol. 2000, 40, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Xu, J. Mitochondria are inherited from the MATa parent in crosses of the basidiomycete fungus Cryptococcus neoformans. Genetics 2003, 163, 1315–1325. [Google Scholar]
- Fedler, M.; Luh, K.S.; Stelter, K.; Nieto-Jacobo, F.; Basse, C.W. The a2 mating-type locus genes lga2 and rga2 direct uniparental mitochondrial DNA (mtDNA) inheritance and constrain mtDNA recombination during sexual development of Ustilago maydis. Genetics 2009, 181, 847–860. [Google Scholar] [CrossRef]
- Yan, Z.; Hull, C.; Sun, S.; Heitman, J.; Xu, J. The mating type-specific homeodomain genes SXI1a and SXI2a coordinately control uniparental mitochondrial inheritance in Cryptococcus neoformans. Curr. Genet. 2007, 3, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Gyawali, R.; Lin, X. Prezygotic and postzygotic control of uniparental mitochondrial DNA inheritance in Cryptococcus neoformans. mBio 2013, 4, 00112–00113. [Google Scholar] [CrossRef]
- Wang, Z.X.; Wilson, A.; Xu, J. Mitochondrial DNA inheritance in the human fungal pathogen Cryptococcus gattii. Fungal Genet. Biol. 2015, 75, 1–10. [Google Scholar] [CrossRef]
- Kües, U. Life history and developmental processes in the basidiomycete Coprinus cinereus. Microbiol. Mol. Biol. Rev. 2000, 64, 316–353. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Fukumasa-Nakai, Y. Mitochondrial DNA inheritance in sexual crosses of Pleurotus ostreatus. Curr. Genet. 1996, 30, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Specht, C.A.; Novotny, C.P.; Ullrich, R.C. Mitochondrial DNA of Schizophyllum commune: Restriction map, genetic map, and mode of inheritance. Curr. Genet. 1992, 22, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Hintz, W.; Anderson, J.B.; Horgen, P.A. Nuclear migration and mitochondrial inheritance in the mushroom Agaricus bitorquis. Genetics 1988, 119, 35–41. [Google Scholar] [PubMed]
- Bastide, P.Y.; Horgen, P.A. Mitochondrial inheritance and the detection of non-parental mitochondrial DNA haplotypes in crosses of Agaricus bisporus homokaryons. Fungal Genet. Biol. 2003, 38, 333–342. [Google Scholar] [CrossRef]
- Baptista-Ferreira, J.L.C.; Economou, A.; Casselton, L.A. Mitochondrial genetics of Coprinus: Recombination of mitochondrial genomes. Curr. Genet. 1983, 7, 405–407. [Google Scholar] [CrossRef] [PubMed]
- May, G.; Taylor, J.W. Patterns of mating and mitochondrial DNA inheritance in the agaric basidiomycete Coprinus cinereus. Genetics 1988, 118, 213–220. [Google Scholar]
- Ellingboe, A.H.; Raper, J.R. Somatic recombination in Schizophyllum commune. Genetics 1962, 47, 85–98. [Google Scholar] [PubMed]
- Nieuwenhuis, B.P.S.; Debets, A.J.M.; Aanen, D.K. Sexual selection in mushroom-forming basidiomycetes. Proc. R. Soc. B 2011, 278, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Ha, B.; Kim, S.; Ro, H.S. Activation of the mating pheromone response pathway of Lentinula edodes by synthetic pheromones. Mycobiology 2018, 46, 407–415. [Google Scholar] [CrossRef]
- Ha, B.; Lee, S.; Kim, S.; Kim, M.; Moon, Y.J.; Song, Y.; Ro, H.S. Nucleus-selective expression of laccase genes in the dikaryotic dtrain of Lentinula edodes. Mycobiology 2017, 45, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Ha, B.; Kim, S.; Kim, M.; Moon, Y.J.; Song, Y.; Ryu, J.S.; Ryu, H.; Ro, H.S. Diversity of A mating type in Lentinula edodes and mating type preference in domesticated strains. J. Microbiol. 2018, 56, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Ha, B.; Moon, Y.J.; Song, Y.; Kim, S.; Kim, M.; Yoon, C.W.; Ro, H.S. Molecular analysis of B mating type diversity in Lentinula edodes. Sci. Hortic. 2019, 243, 55–63. [Google Scholar] [CrossRef]
- Lunt, D.H.; Whipple, L.E.; Hyman, B.C. Mitochondrial DNA variable number tandem repeats (VNTRs): Utility and problems in molecular ecology. Mol. Ecol. 1998, 7, 1441–1455. [Google Scholar] [CrossRef] [PubMed]
- Shim, D.; Park, S.G.; Kim, K.; Bae, W.; Lee, G.W.; Ha, B.S.; Ro, H.S.; Kim, M.; Ryoo, R.; Rhee, S.K.; et al. Whole genome de novo sequencing and genome annotation of the world popular cultivated edible mushroom, Lentinula edodes. J. Biotechnol. 2016, 223, 24–25. [Google Scholar] [CrossRef] [PubMed]
- Benson, G. Tandem repeats finder: a program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basse, C.W. Mitochondrial inheritance in fungi. Curr. Opin. Microbiol. 2010, 13, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.; Xu, J. Mitochondrial inheritance: diverse patterns and mechanisms with an emphasis on fungi. Mycology 2012, 3, 158–166. [Google Scholar]
- Wang, Y.; Zeng, F.; Hon, C.C.; Zhang, Y.; Leung, F.C. The mitochondrial genome of the basidiomycete fungus Pleurotus ostreatus (oyster mushroom). FEMS Microbiol. Lett. 2008, 280, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, N.; Zhang, H.; Yang, X.J.; Huang, Y.; Lei, F. Extreme variation in patterns of tandem repeats in mitochondrial control region of yellow-browed tits (Sylviparus modestus, Paridae). Sci. Rep. 2015, 5, 13227. [Google Scholar] [CrossRef]
- Lunt, D.H.; Hyman, B.C. Animal mitochondrial DNA recombination. Nature 1997, 387, 247. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, S.; Kubo, T.; Mikami, T. Variable number of tandem repeat loci in the mitochondrial genomes of beets. Curr. Genet. 2000, 37, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Levinson, G.; Gutman, G.A. Slipped-strand mispairing: A major mechanism for DNA sequence evolution. Mol. Biol. Evol. 1987, 43, 203–221. [Google Scholar]
- Mundy, N.I.; Helbig, A.J. Origin and evolution of tandem repeats in the mitochondrial DNA control region of shrikes (Lanius spp.). J. Mol. Evol. 2004, 59, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Albert, B.; Sellem, C. Dynamics of the mitochondrial genome during Podospora anserina aging. Curr. Genet. 2002, 40, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Lobachev, K.S.; Shor, B.M.; Tran, H.T.; Taylor, W.; Keen, J.D.; Resnick, M.A.; Gordenin, D.A. Factors affecting inverted repeat stimulation of recombination and deletion in Saccharomyces cerevisiae. Genetics 1998, 148, 1507–1524. [Google Scholar] [PubMed]
- Aguileta, G.; de Vienne, D.M.; Ross, O.N.; Hood, M.E.; Giraud, T.; Petit, E.; Gabaldon, T. High variability of mitochondrial gene order among fungi. Genome Biol. Evol. 2014, 6, 451–465. [Google Scholar] [CrossRef]
- Moon, S.; Lee, H.Y.; Shim, D.; Kim, M.; Ka, K.H.; Ryoo, R.; Ko, H.K.; Koo, C.D.; Chung, J.W.; Ryu, H. Development and molecular characterization of novel polymorphic genomic DNA SSR markers in Lentinula edodes. Mycobiology 2017, 45, 105–109. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Number | Nuclear Type | Mitochondrial Type in the Di–Mon Matea | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Di | Mon | Mate | VNTR7 | VNTR13 | VNTR18 | VNTR20 | VNTR23 | VNTR25 | Overall | |
| IUM3182 | A9/A11 | A1 | A1/A11 | Mon | Mon | Mon | Monb | Mon | Mon | Mon |
| IUM3179 | A11/A19 | A1/A19 | Mon | Mon | Mon | Mon | Mon | Mon | Mon | |
| KFRI3010 | A47/A54 | A1/A54 | Mon | NDc | ND | ND | Mon | Mon | Mon | |
| IUM3178 | A14/A18 | A1/A14 | Mon | ND | ND | ND | Mon | Mon | Mon | |
| KFRI665 | A38/A61 | A1/A38 | UPd | ND | Mon | ND | Mon | Mon | Mon | |
| KFRI956 | A41/A65 | A1/A65 | Mon | Mon | Mon | ND | Mon | Mon | Mon | |
| KFRI39 | A33/A57 | A1/A33 | Mon | Mon | Mon | ND | Mon | UP | Mon | |
| KFRI664 | A37/A60 | A1/A37 | Mon | Mon | Mon | Mon | Mon | Mon | Mon | |
| KFRI1520 | A43/A52 | A1/A52 | Mon | Mon | Mon | Mon | Mon | Mon | Mon | |
| KFRI2521 | A46/A53 | A1/A46 | Mon | Mon | Mon | Mon | ND | Mon | Mon | |
| IUM5054 | A12/A13 | A1/A12 | Mon | Mon | Mon | Mon | ND | Mon | Mon | |
| NAAS6640 | A12/A49 | A1/A12 | UP | Mon | Mon | Mon | Mon | Mon | Mon | |
| NAAS4255 | A19/A63 | A1/A63 | Mon | Mon | Mon | Mon | Mon | UP | Mon | |
| NAAS5735 | A27/A48 | A1/A48 | ND | Mon | ND | Mon | Mon | ND | Mon | |
| NAAS6833 | A50/A55 | A1/A50 | Mon | ND | Mon | ND | Mon | ND | Mon | |
| KFRI57 | A34/A58 | A1/A34 | Mon | Mon | Mon | ND | Mon | ND | Mon | |
| KFRI411 | A36/A59 | A1/A59 | Mon | Mon | Mon | Mon | Mon | Mon | Mon | |
| KFRI674 | A44/A64 | A1/A64 | ND | Mon | Mon | Monb | Mon | Mon | Mon | |
| IUM4848 | A10/A17 | A1/A17 | Mon | Mon | Mon | ND | Mon | Mon | Mon | |
| KFRI666 | A39/A62 | A1/A39 | ND | Mon | Mon | ND | Mon | Mon | Mon | |
| KFRI58 | A17/A35 | A1/A35 | Mon | Mon | ND | Mon | Mon | Mon | Mon | |
| IUM4841 | A1/A16 | A1/A16 | Mon | Mon | ND | Mon | Mon | Mon | Mon | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Song, Y.; Ha, B.; Moon, Y.J.; Kim, M.; Ryu, H.; Ro, H.-S. Variable Number Tandem Repeats in the Mitochondrial DNA of Lentinula edodes. Genes 2019, 10, 542. https://doi.org/10.3390/genes10070542
Kim S, Song Y, Ha B, Moon YJ, Kim M, Ryu H, Ro H-S. Variable Number Tandem Repeats in the Mitochondrial DNA of Lentinula edodes. Genes. 2019; 10(7):542. https://doi.org/10.3390/genes10070542
Chicago/Turabian StyleKim, Sinil, Yelin Song, Byeongsuk Ha, Yoon Jung Moon, Minseek Kim, Hojin Ryu, and Hyeon-Su Ro. 2019. "Variable Number Tandem Repeats in the Mitochondrial DNA of Lentinula edodes" Genes 10, no. 7: 542. https://doi.org/10.3390/genes10070542
APA StyleKim, S., Song, Y., Ha, B., Moon, Y. J., Kim, M., Ryu, H., & Ro, H. -S. (2019). Variable Number Tandem Repeats in the Mitochondrial DNA of Lentinula edodes. Genes, 10(7), 542. https://doi.org/10.3390/genes10070542