1. Introduction
The emergence and spread of parasites resistant to antimalarial drugs and mosquitoes resistant to insecticides threaten the recent gains in malaria control and challenge the goal for malaria elimination in the Greater Mekong Subregion (GMS) of Southeast Asia [
1]. Antimalarial drug resistance in
P. falciparum tends to emerge in low-transmission settings, particularly in Southeast Asia and South America, before expanding to high-transmission settings in sub-Saharan Africa [
2]. Resistance to chloroquine (CQ) and later to sulfadoxine-pyrimethamine (SP) has been responsible for the increased mortality in African children [
3,
4]. Artemisinin (ART)-resistant
P. falciparum parasites were first reported in Cambodia and subsequently in all countries of the GMS [
5]. This has accelerated resistance development in parasites to artemisinin combination therapy (ACT) partner drugs, resulting in increased treatment failure rates with dihydroartemisinin-piperaquine (DP) in Cambodia and with artesunate-mefloquine (AS-MQ) in Cambodia and on the Thai-Myanmar border [
6,
7]. The timely monitoring of antimalarial resistance spread is essential for maintaining the recent progress in malaria control and for achieving the goal of regional malaria elimination.
As a high malaria-burden country within the GMS, Myanmar uses several first-line treatments for
P. falciparum infections, including artemether-lumefantrine, AS-MQ, and DP. However, delayed clearance has been observed in all the three first-line ACTs [
8]. Similar to other malaria-endemic areas, Myanmar experiences a large proportion of asymptomatic malaria infections, as demonstrated by active case surveillance of malaria infections among healthy populations [
9,
10]. In the Shwegyin township of southern Myanmar, mutations in genes associated with drug resistance including those with ART resistance were identified in asymptomatic infections [
11]. Asymptomatic or sub-clinical malaria infections seldom cause acute disease, but they are capable of infecting mosquitoes, thus serving as silent reservoirs for continued malaria transmission. Therefore, prevalent antimalarial drug resistant strains in asymptomatic infections may underlie the rapid spread of drug resistance. Given the limited data on drug resistance in asymptomatic infections, ongoing molecular epidemiological studies of drug resistance are needed in these parasite reservoirs.
Molecular epidemiology studies provide information for detecting the emergence and tracking the spread of antimalarial drug resistance. Several mutations in the
P. falciparum dihydrofolate reductase (
pfdhfr) and
P. falciparum dihydropteroate synthase (
pfdhps) genes (e.g., triple mutations at codons 51, 59 and 108 of
pfdhfr and double mutations at codons 437 and 540 of
pfdhps) are associated with SP treatment failures [
12]. The
P. falciparum chloroquine resistance transporter (
pfcrt) K76T mutation and
P. falciparum multidrug resistance protein 1 (
pfmdr1) N86Y mutation have been linked to CQ and amodiaquine resistance [
13]. Mutations in the
P. falciparum multidrug resistance-associated protein 1 (
pfmrp1), such as H191Y and S437A, were reported to be associated with resistance to CQ and quinine (QN) in vitro [
14]. Point mutations in the propeller domain of Kelch protein 13 (
k13) were collectively correlated with clinical ART resistance [
15,
16]. Molecular surveillance showed that
k13 mutations associated with ART resistance were restricted to certain areas of the GMS, with C580Y being the predominant mutation in the Thai-Cambodian region and F446I in China and northern Myanmar [
17,
18,
19,
20].
Here, we profiled mutations in antimalarial drug resistance genes in asymptomatic P. falciparum infections identified from cross-sectional studies in three townships of Myanmar. The results will provide updated information on the drug resistance status of P. falciparum malaria in multiple sentinel sites in Myanmar, which is necessary for developing strategies to eliminate P. falciparum infections in this country.
4. Discussion
As the GMS is moving towards malaria elimination, Myanmar requires immediate attention because of the high malaria burden and its geographical connection with South Asia, through which drug resistance could spread rapidly. According to WHO reports, the latest malaria drug resistance map reveals that both treatment failure and resistant genotypes are prevalent in Myanmar, including our three study sites [
31]. Irrespective of the ACTs used, from 2010 to 2018, treatment failure among
P. falciparum patients in Kachin State was higher than that in Chin State. Our study found a higher prevalence of
k13 and
pfmdr1 mutations in Kachin State (east) than in Chin State (west) which is possibly contributing to the treatment failure in Kachin State. Asymptomatic
Plasmodium infections, as silent reservoirs of malaria parasites, play an important role in continued malaria transmission. Critically, these asymptomatic infections may carry genetic variations conferring drug resistance, thus facilitating the spread of drug-resistant parasites [
11]. Therefore, this study aimed to provide molecular epidemiology information about drug-resistant
P. falciparum in asymptomatic infections at three sentinel sites located in southwestern, northern and northeastern Myanmar, each with a different malaria epidemiology.
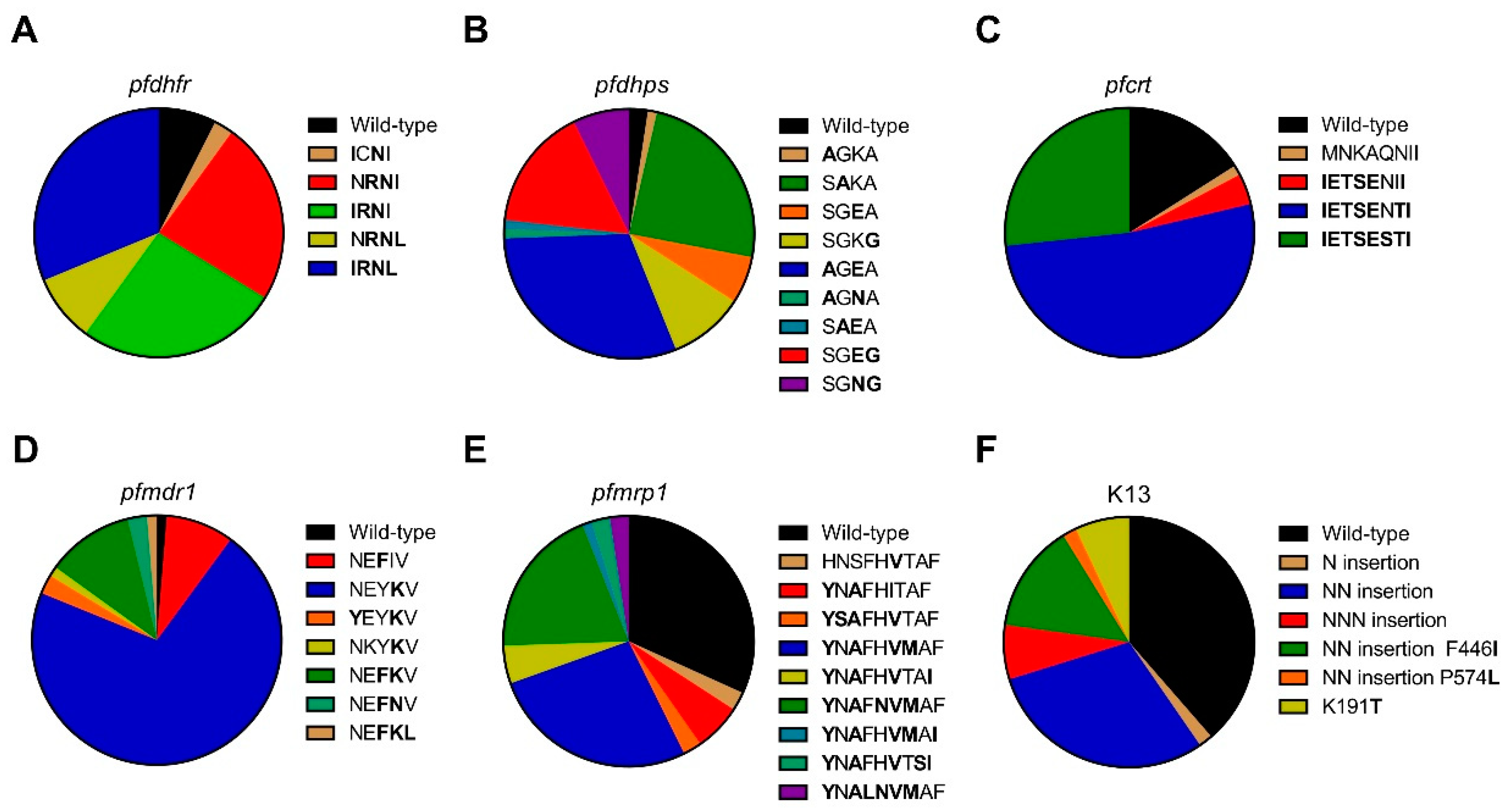
The drug combination SP, commercially known as Fansidar, has been used as an antimalarial chemotherapy in Southeast Asia and Africa. The mechanism of resistance to sulfadoxine has been associated with five point mutations at the S436A/F, G437A, K540E, A581G, and A613T/S codons of the
pfdhps gene. Mutations at 437 and 540 were strongly associated with SP treatment failure, whereas the 436, 581, and 613 mutations confer some degrees of resistance [
32]. In this study, A613T/S was not observed, but K540N was prevalent, consistent with a previous study from northeast Myanmar [
33]. A581G, G437A, and K540E were the predominant mutations in Laiza, Banmauk, and Paletwa, respectively. In population studies, mutations at codon 59 of the
pfdhfr gene and codon 540 of the
pfdhps gene are strongly predictive of SP treatment failure [
34,
35]. The prevalence of this mutation combination in Laiza and Paletwa was 100% and 83.3%, respectively, and was slightly lower in Banmauk (34.1%), suggesting that strong selection pressure against SP was emerging in western and northeastern Myanmar. Similarly, triple mutations at codons 108, 51, and 59 of
pfdhfr and double mutations at codons 437 and 540 of
pfdhps are associated with SP treatment failure [
12]. Here, one isolate was found to have this quintuple mutation combination in Banmauk, and 12 isolates (nine in Laiza, two in Banmauk and one in Paletwa) harbored the sextuple mutations with
pfdhfr (N51I, C59R, S108N, and I164L) and
pfdhps (K540N/E and A581G), suggesting that
P. falciparum from these asymptomatic carriers would be highly resistant to SP.
Mutations in the
pfcrt gene, especially the K76T mutation, are major determinants of CQ resistance [
36,
37]. In addition,
pfcrt mutations also influence
P. falciparum susceptibility to mefloquine (MQ), halofantrine (HF), and ART [
38]. The K76T mutation is associated with different sets of mutations at different codons, most commonly C72S, M74I, N75E, A220S, Q271E, N326S, I356T, and R371I [
39]. Consistent with a previous study conducted in Thailand [
25], C72S was not found. Except for N326S, other mutations reached high prevalence in all three sites. Notably, almost all parasite isolates had the CIVIET haplotype around codons 72-76, which is the typical CQ-resistant haplotype in Southeast Asia. The stable and highly prevalent of
pfcrt mutations may be the result of continued use of CQ as the first-line treatment for
P. vivax infections in this region [
40,
41]. In addition to CQ resistance,
pfcrt has gained recognition as a multidrug resistance transporter, which influences parasites’ susceptibilities to multiple first-line antimalarial drugs [
42,
43]. Emerging mutations in
pfcrt, H97Y, F145I, M343L, and G353V, also have been linked to piperaquine (PPQ) resistance [
44,
45], but these mutations were not identified in this study. Thus, the high prevalence of
pfcrt mutations in asymptomatic
P. falciparum parasites may be linked to failures of multiple ACTs in Myanmar.
The ATP-binding cassette (ABC) transporters, including
pfmdr1 and
pfmrp1, are potentially involved in resistance to multiple antimalarial drugs [
46,
47]. Several point mutations in
pfmdr1 are associated with changed sensitivities to CQ, MQ, QN, HF, and ART [
38,
48,
49,
50]. In the presence of
pfcrt K76T, point mutations in
pfmdr1, primarily at codon 86 [
13,
14] and additionally at positions 184, 1034, 1042, and 1246 [
51], can increase resistance of the parasites to CQ. Decreased susceptibility to lumefantrine (LMF) has also been linked to polymorphism in
pfcrt and
pfmdr1 [
52,
53]. In this study, only two
P. falciparum isolates from the western township Paletwa had the
pfcrt K76T and
pfmdr1 N86Y mutations, but the combination of
pfcrt K76T and
pfmdr1 Y184F had much higher prevalence in Laiza (58.3%) than Banmauk (18.0%) and Paletwa (12.5%). The
pfmdr1 Y184F was not detected in an earlier study [
54], where I185K was reported as a novel mutation. The authors speculated that the decrease in parasite’s susceptibility to the drug was associated with substitution of mutation at codon Y184F for I185K. In our study, the
pfmdr1 I185K mutation was highly prevalent (~90% in all samples) and reached fixation in Banmauk and Paletwa. It would be worthwhile to establish whether this mutation is linked to altered drug sensitivity through genetic studies given the high prevalence of this mutation. Since disruption of
pfmrp1 in a CQ-resistant
P. falciparum isolate rendered this parasite more sensitive to CQ, QN, PPQ, primaquine, and ART, PfMRP1 is proposed to mediate resistance of the parasite to multiple antimalarial drugs [
55]. Association studies suggest that mutations at codons Y191H, A437S, H785N, I876V and F1390I in
pfmrp1 contribute to decreased sensitivity to CQ, PPQ, LMF, artesunate (ATS) and dihydroartemisinin (DHA) as well as reduce both in vivo and in vitro susceptibility to ACT [
28,
33,
56]. Our results suggest that
P. falciparum isolates from asymptomatic infections in these areas might be resistant to CQ, PPQ, LMF, ATS, DHA, and ACT.
ART resistance in
P. falciparum has emerged in western Cambodia and its potential spread to Africa threatens global malaria control [
57]. ART resistance is characterized by delayed parasite clearance, which corresponds to the decreased susceptibility of ring-stage parasites [
58]. Non-synonymous SNPs in K13 were found to be strongly associated with resistance to ART [
16]. To date, more than 200 non-synonymous mutations in the
k13 gene have been reported. N458Y, Y493H, R539T, I543T, and C580Y are validated ART resistance mutations, and a number of mutations are candidates for ART resistance, including P574L [
8]. In Africa, non-synonymous
k13 mutations are relatively rare [
59]. Recently,
k13 mutations have emerged independently in multiple locations in Southeast Asia, including Myanmar [
17]. C580Y, Y493H, R539T, and I543T
k13 mutations were prevalent in the Thai-Myanmar border, whereas F446I was the most prevalent mutant allele of
k13 in the northern Myanmar and China-Myanmar border, and P574L next to C580Y were reported among isolates collected from migrant goldmine workers in southern Myanmar [
19,
60,
61,
62]. In our study, the validated mutations of
k13 resistance were not found, while F446I, P574L, and K191T mutations in the
k13 propeller domain were observed among parasites from asymptomatic infections. The K191T has not been reported in previous studies. F446I mutation was observed with high prevalence only at Laiza along China-Myanmar border, which is consistent with previous studies. F446I mutation in
k13 has been found to be associated with delayed parasite clearance [
61], and a recent study showed that F446I was associated with the increased ring survival rates by genetically introducing the mutation into the
k13 gene [
63]. Consistent with previous studies of
P. falciparum from a symptomatic population, the absence of
k13 propeller mutations in Paletwa suggest that ART resistant parasites may not have spread to the western Myanmar region [
18]. Moreover, we found the N, NN, and NNN insertion in the N terminus of
k13 protein through analysis of the full-length
k13 sequences. A previous study reported that an NN insertion in the N terminus of
k13 protein was associated with increased in vitro ring-stage survival in the presence of DHA, but the possibility of the NN insert acting cooperatively with F446I in conferring ART resistance was not ruled out [
33]. In view of the high prevalence (45.6%) of the NN insertion among
P. falciparum from asymptomatic populations in Myanmar, it would be valuable to determine whether it independently affects ART resistance.
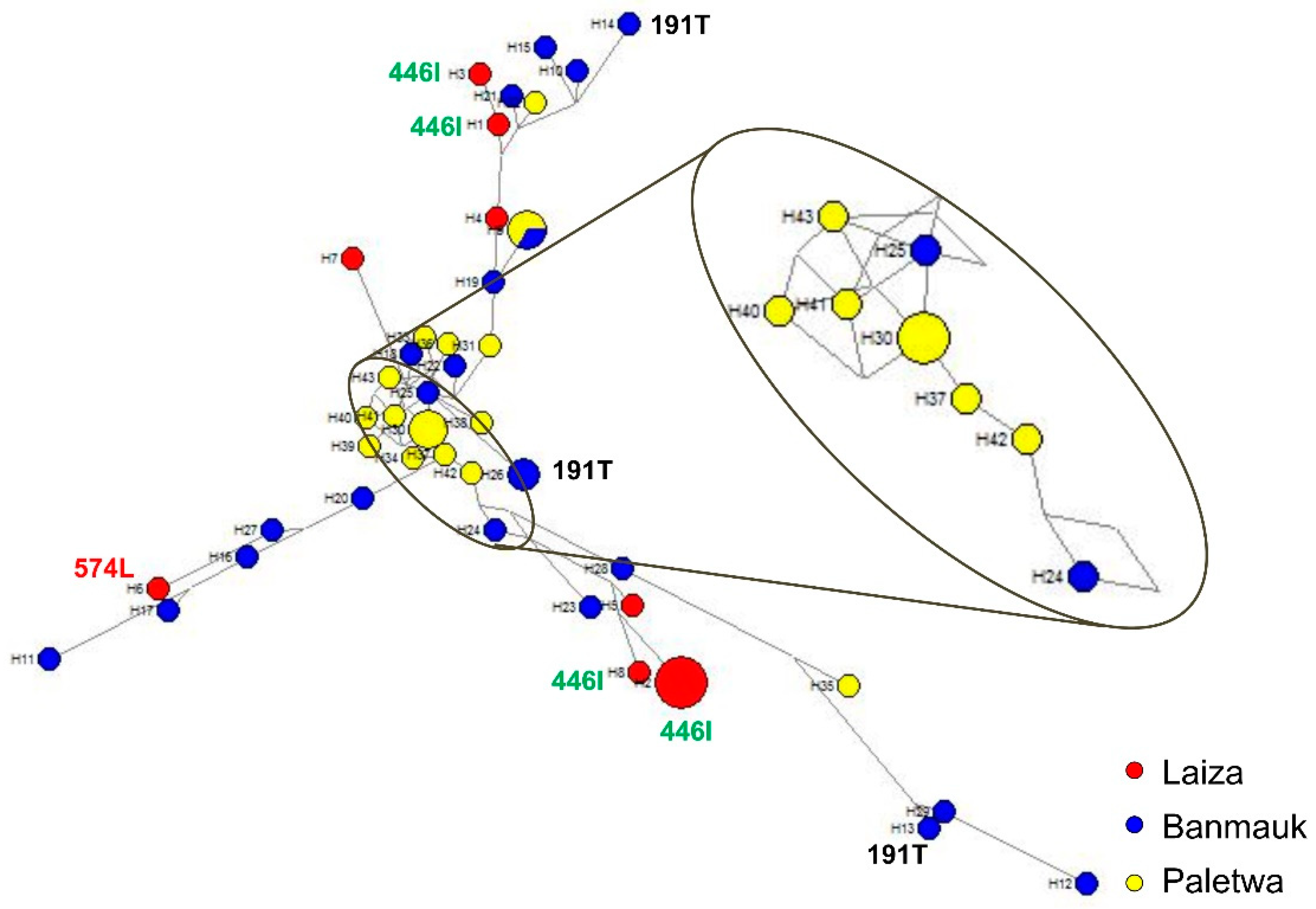
The generation of the haplotype network can be used to study the phylogenetic relationship of parasites, to determine the origins of drug resistance gene mutations from different geographical regions, and to assess the diversity of parasite populations. Consistent with the results from clinical P. falciparum isolates, parasites from asymptomatic individuals also displayed extraordinarily high diversity in Myanmar. The genotypes of the six genes associated with drug resistance indicated that the asymptomatic P. falciparum infections in Myanmar are resistant to multiple drugs. More importantly, 58.8% (20/34) haplotypes carried k13 mutations (N458Y, R539T, P574L, and F446I) associated with ART resistance in clinical isolates, whereas 62.5% (5/8) haplotypes of asymptomatic P. falciparum isolates carried F446I and P574L. Although only two k13 mutations related to ART resistance were detected in our study, the presence of such high haplotype diversity in the limited isolates of asymptomatic infection should be concerning. Despite reduced P. falciparum prevalence in recent years, the evidence suggests that ART resistant P. falciparum parasites appear independently and spread widely along the China-Myanmar border, which is a reminder that surveillance of P. falciparum should be strengthened both in Myanmar and China. Although k13 mutation associated with ART resistance was not found in the other two areas, the diversity of multiple drug resistance reflected by haplotype analysis should not be ignored.

 ,
, 



{kind=link}
{kind=link}
{kind=link}