The Female-Specific W Chromosomes of Birds Have Conserved Gene Contents but Are Not Feminized


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genomic Dataset
2.2. Identification of the Sex-Linked Sequences in Paleognathae
2.3. Demarcation of the Paleognathous Evolutionary Strata
2.4. Gene Expression Analyses
2.5. Gene Content Analysis
2.6. HINT1W Copy Number Estimation
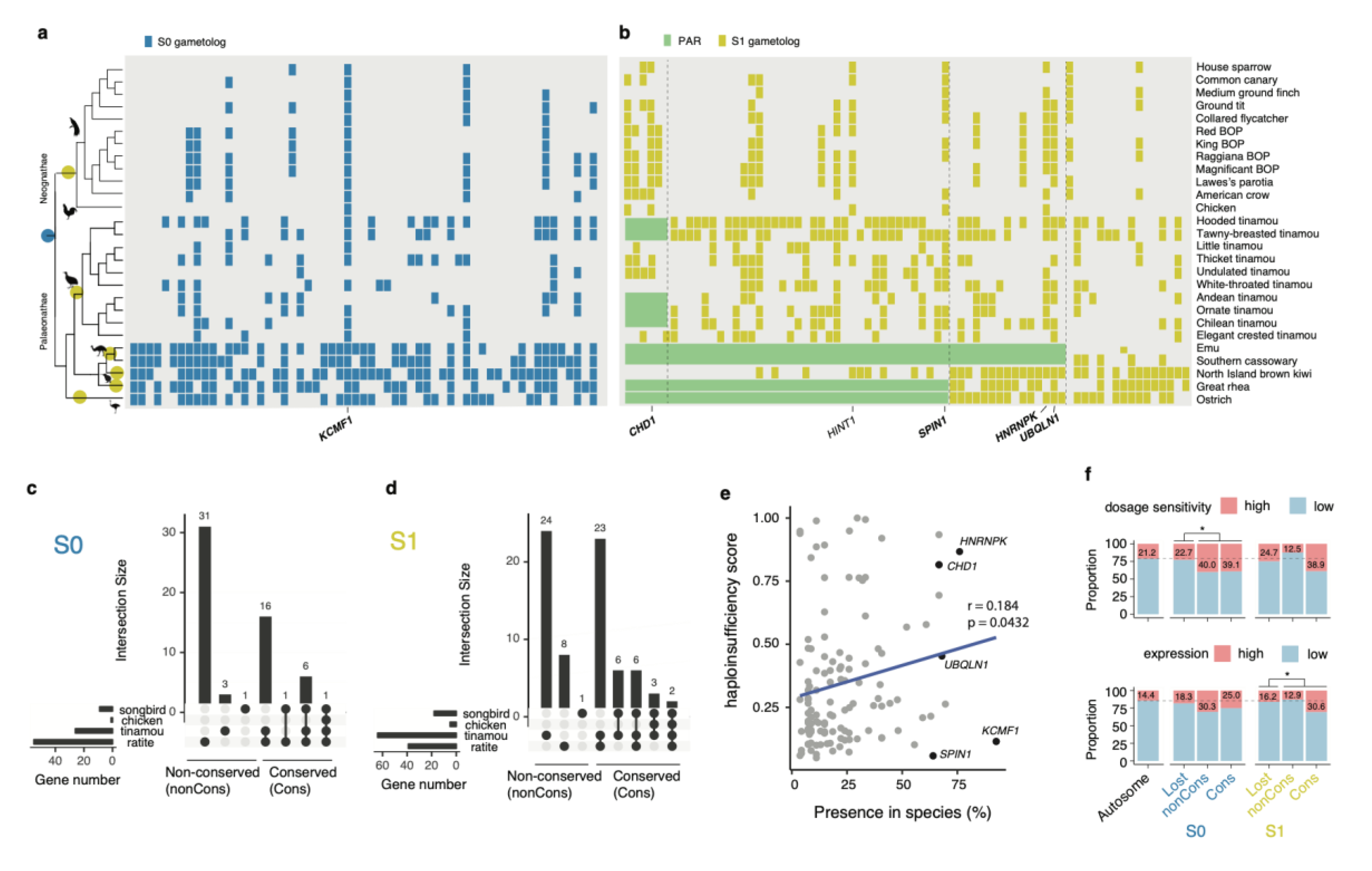
3. Results
3.1. Independent Sex Chromosome Differentiation in Palaeognathous Birds
3.2. Demarcation of the Evolutionary Strata of Sex Chromosomes
3.3. Deep Conservation of the Gene Content of the Avian W Chromosomes
3.4. Genes on the W Chromosomes Have High Expression and High Dosage Sensitivity
3.5. Little Evidence of Female-Specific Positive Selection on the Bird W Chromosomes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bachtrog, D. Y-chromosome evolution: Emerging insights into processes of Y-chromosome degeneration. Nat. Rev. Genet. 2013, 14, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, W.R. Genetic Hitchhiking and the Evolution of Reduced Genetic Activity of the Y Sex Chromosome. Genetics 1987, 116, 161–167. [Google Scholar] [PubMed]
- Charlesworth, B. Model for evolution of Y chromosomes and dosage compensation. Proc. Natl. Acad. Sci. USA 1978, 75, 5618–5622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, W.R. Evolution of the Y Sex Chromosome in Animals. Bioscience 1996, 46, 331–343. [Google Scholar] [CrossRef] [Green Version]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.-L.; Hahn, M.W.; Kitano, J.; Mayrose, V.B.; Ming, R.; et al. Sex Determination: Why So Many Ways of Doing It? PLoS Boil. 2014, 12, e1001899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortez, D.; Marin, R.; Toledo-Flores, D.; Froidevaux, L.; Liechti, A.; Waters, P.D.; Grutzner, F.; Kaessmann, H. Origins and functional evolution of Y chromosomes across mammals. Nature 2014, 508, 488–493. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhang, J.; Bachtrog, R.; An, N.; Huang, Q.; Jarvis, E.D.; Gilbert, M.T.P.; Zhang, G. Complex evolutionary trajectories of sex chromosomes across bird taxa. Science 2014, 346, 1246338. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Auer, G.; Peona, V.; Suh, A.; Deng, Y.; Feng, S.; Zhang, G.; Blom, M.P.K.; Christidis, L.; Prost, S.; et al. Dynamic evolutionary history and gene content of sex chromosomes across diverse songbirds. Nat. Ecol. Evol. 2019, 3, 834–844. [Google Scholar] [CrossRef]
- Rice, W.R. The Accumulation of Sexually Antagonistic Genes as a Selective Agent Promoting the Evolution of Reduced Recombination between Primitive Sex Chromosomes. Evolution 1987, 41, 911–914. [Google Scholar] [CrossRef]
- Mank, J.E. Population genetics of sexual conflict in the genomic era. Nat. Rev. Genet. 2017, 18, 721–730. [Google Scholar] [CrossRef]
- Mank, J.E.; Hosken, D.J.; Wedell, N. Conflict on the Sex Chromosomes: Cause, Effect, and Complexity. Cold Spring Harb. Perspect. Boil. 2014, 6, a017715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soh, Y.Q.S.; Alföldi, J.; Pyntikova, T.; Brown, L.G.; Graves, T.; Minx, P.J.; Fulton, R.S.; Kremitzki, C.; Koutseva, N.; Mueller, J.L.; et al. Sequencing the mouse Y chromosome reveals convergent gene acquisition and amplification on both sex chromosomes. Cell 2014, 159, 800–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Bachtrog, R. Sex-Specific Adaptation Drives Early Sex Chromosome Evolution in Drosophila. Science 2012, 337, 341–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachtrog, D. The Y Chromosome as a Battleground for Intragenomic Conflict. Trends Genet. 2020, 36, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Koerich, L.B.; Wang, X.; Clark, A.G.; De Carvalho, A.B. Low conservation of gene content in the Drosophila Y chromosome. Nature 2008, 456, 949–951. [Google Scholar] [CrossRef] [Green Version]
- De Carvalho, A.B. Origin and evolution of the Drosophila Y chromosome. Curr. Opin. Genet. Dev. 2002, 12, 664–668. [Google Scholar] [CrossRef]
- De Carvalho, A.B.; Dobo, B.A.; Vibranovski, M.D.; Clark, A.G. Identification of five new genes on the Y chromosome of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2001, 98, 13225–13230. [Google Scholar] [CrossRef] [Green Version]
- Bachtrog, D.; Mahajan, S.; Bracewell, R. Massive gene amplification on a recently formed Drosophila Y chromosome. Nat. Ecol. Evol. 2019, 3, 1587–1597. [Google Scholar] [CrossRef]
- Larson, E.L.; Kopania, E.E.K.; Good, J.M. Spermatogenesis and the Evolution of Mammalian Sex Chromosomes. Trends Genet. 2018, 34, 722–732. [Google Scholar] [CrossRef]
- Ellison, C.E.; Bachtrog, D. Recurrent gene co-amplification on Drosophila X and Y chromosomes. PLoS Genet. 2019, 15, e1008251. [Google Scholar] [CrossRef] [Green Version]
- Mahajan, S.; Bachtrog, D. Convergent evolution of Y chromosome gene content in flies. Nat. Commun. 2017, 8, 785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albritton, S.E.; Kranz, A.-L.; Rao, P.; Kramer, M.J.; Dieterich, C.; Ercan, S. Sex-Biased Gene Expression and Evolution of the X Chromosome in Nematodes. Genetics 2014, 197, 865–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, S.L.; Bonduriansky, R.; Chenoweth, S.F. The genomic distribution of sex-biased genes in drosophila serrata: X chromosome demasculinization, feminization, and hyperexpression in both sexes. Genome Boil. Evol. 2013, 5, 1986–1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sturgill, D.; Zhang, Y.; Parisi, M.; Oliver, B. Demasculinization of X chromosomes in the Drosophila genus. Nature 2007, 450, 238–241. [Google Scholar] [CrossRef] [Green Version]
- Vicoso, B.; Bachtrog, D. Numerous transitions of sex chromosomes in Diptera. PLoS Boil. 2015, 13, e1002078. [Google Scholar] [CrossRef] [Green Version]
- Bachtrog, D. A dynamic view of sex chromosome evolution. Curr. Opin. Genet. Dev. 2006, 16, 578–585. [Google Scholar] [CrossRef]
- Reinius, B.; Johansson, M.; Radomska, K.J.; Morrow, E.H.; Pandey, G.K.; Kanduri, C.; Sandberg, R.; Williams, R.W.; Jazin, E. Abundance of female-biased and paucity of male-biased somatically expressed genes on the mouse X-chromosome. BMC Genom. 2012, 13, 607. [Google Scholar] [CrossRef] [Green Version]
- Khil, P.P.; A Smirnova, N.; Romanienko, P.J.; Camerini-Otero, R.D. The mouse X chromosome is enriched for sex-biased genes not subject to selection by meiotic sex chromosome inactivation. Nat. Genet. 2004, 36, 642–646. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.J.; McCarrey, J.R.; Yang, F.; Page, D.C. An abundance of X-linked genes expressed in spermatogonia. Nat. Genet. 2001, 27, 422–426. [Google Scholar] [CrossRef]
- Rice, W.R. Sex Chromosomes and the Evolution of Sexual Dimorphism. Evolution 1984, 38, 735–742. [Google Scholar] [CrossRef]
- Ellegren, H. Emergence of male-biased genes on the chicken Z-chromosome: Sex-chromosome contrasts between male and female heterogametic systems. Genome Res. 2011, 21, 2082–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mank, J.E.; Ellegren, H. Sex-linkage of sexually antagonistic genes is predicted by female, but not male, effects in birds. Evolution 2009, 63, 1464–1472. [Google Scholar] [CrossRef] [PubMed]
- Bellott, D.W.; Skaletsky, H.; Pyntikova, T.; Mardis, E.R.; Graves, T.; Kremitzki, C.; Brown, L.G.; Rozen, S.G.; Warren, W.C.; Wilson, R.K.; et al. Convergent evolution of chicken Z and human X chromosomes by expansion and gene acquisition. Nature 2010, 466, 612–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicoso, B.; Kaiser, V.B.; Bachtrog, D. Sex-biased gene expression at homomorphic sex chromosomes in emus and its implication for sex chromosome evolution. Proc. Natl. Acad. Sci. USA 2013, 110, 6453–6458. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Sin, S.Y.W.; Grayson, P.; Edwards, S.V.; Sackton, T.B. Evolutionary Dynamics of Sex Chromosomes of Paleognathous Birds. Genome Boil. Evol. 2019, 11, 2376–2390. [Google Scholar] [CrossRef] [Green Version]
- Moghadam, H.K.; Pointer, M.A.; Wright, A.E.; Berlin, S.; Mank, J.E. W chromosome expression responds to female-specific selection. Proc. Natl. Acad. Sci. USA 2012, 109, 8207–8211. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, M.; Binder, M.D.; A Smith, C.; Andrews, J.; Reed, K.; Smith, M.; Millar, C.; Lambert, D.; Sinclair, A.H. ASW: A gene with conserved avian W-linkage and female specific expression in chick embryonic gonad. Dev. Genes Evol. 2000, 210, 243–249. [Google Scholar] [CrossRef]
- Hori, T.; Asakawa, S.; Itoh, Y.; Shimizu, N.; Mizuno, S. Wpkci, Encoding an Altered Form of PKCI, Is Conserved Widely on the Avian W Chromosome and Expressed in Early Female Embryos: Implication of Its Role in Female Sex Determination. Mol. Boil. Cell 2000, 11, 3645–3660. [Google Scholar] [CrossRef] [Green Version]
- Bellott, D.W.; Skaletsky, H.; Cho, T.-J.; Brown, L.; Locke, D.P.; Chen, N.; Galkina, S.; Pyntikova, T.; Koutseva, N.; Graves, T.; et al. Avian W and mammalian Y chromosomes convergently retained dosage-sensitive regulators. Nat. Genet. 2017, 49, 387–394. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zhang, J.; Xu, X.; Witt, C.C.; Deng, Y.; Chen, G.; Meng, G.; Feng, S.; Székely, T.; Zhang, G.; et al. Phylogeny, transposable element and sex chromosome evolution of the basal lineage of birds. bioRxiv 2019, 750109. [Google Scholar] [CrossRef]
- Marçais, G.; Delcher, A.L.; Phillippy, A.M.; Coston, R.; Salzberg, S.L.; Zimin, A.V. MUMmer4: A fast and versatile genome alignment system. PLoS Comput. Boil. 2018, 14, e1005944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonge, M.; Soyk, S.; Ramakrishnan, S.; Wang, X.; Goodwin, S.; Sedlazeck, F.J.; Lippman, Z.B.; Schatz, M.C. RaGOO: Fast and accurate reference-guided scaffolding of draft genomes. Genome Boil. 2019, 20, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudchenko, O.; Batra, S.S.; Omer, A.D.; Nyquist, S.K.; Hoeger, M.; Durand, N.C.; Shamim, M.S.; Machol, I.; Lander, E.S.; Aiden, A.P.; et al. De novo assembly of the Aedes aegypti genome using Hi-C yields chromosome-length scaffolds. Science 2017, 356, 92–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudchenko, O.; Shamim, M.S.; Batra, S.; Durand, N.C.; Musial, N.T.; Mostofa, R.; Pham, M.; Hilaire, B.G.S.; Yao, W.; Stamenova, E.; et al. The Juicebox Assembly Tools module facilitates de novo assembly of mammalian genomes with chromosome-length scaffolds for under $1000. bioRxiv 2018, 254797. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutierrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Minh, B.Q.; A Schmidt, H.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Boil. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Uebbing, S.; Konzer, A.; Xu, L.; Backström, N.; Brunström, B.; Bergquist, J.; Ellegren, H. Quantitative Mass Spectrometry Reveals Partial Translational Regulation for Dosage Compensation in Chicken. Mol. Boil. Evol. 2015, 32, 2716–2725. [Google Scholar] [CrossRef] [Green Version]
- Uebbing, S.; Künstner, A.; Mäkinen, H.; Ellegren, H. Transcriptome Sequencing Reveals the Character of Incomplete Dosage Compensation across Multiple Tissues in Flycatchers. Genome Boil. Evol. 2013, 5, 1555–1566. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Boil. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, N.; Lee, I.; Marcotte, E.M.; Hurles, M.E. Characterising and Predicting Haploinsufficiency in the Human Genome. PLoS Genet. 2010, 6, e1001154. [Google Scholar] [CrossRef] [Green Version]
- De Clare, M.; Pir, P.; Oliver, S.G. Haploinsufficiency and the sex chromosomes from yeasts to humans. BMC Boil. 2011, 9, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marçais, G.; Kingsford, C. A fast, lock-free approach for efficient parallel counting of occurrences of k-mers. Bioinformatics 2011, 27, 764–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smeds, L.; Warmuth, V.; Bolivar, P.; Uebbing, S.; Burri, R.; Suh, A.; Nater, A.; Bures, S.; Garamszegi, L.Z.; Hogner, S.; et al. Evolutionary analysis of the female-specific avian W chromosome. Nat. Commun. 2015, 6, 7330. [Google Scholar] [CrossRef] [Green Version]
- Ceplitis, H.; Ellegren, H. Adaptive Molecular Evolution of HINTW, a Female-Specific Gene in Birds. Mol. Boil. Evol. 2003, 21, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Graves, J. Evolution of vertebrate sex chromosomes and dosage compensation. Nat. Rev. Genet. 2015, 17, 33–46. [Google Scholar] [CrossRef]
- Ellegren, H.; Hultin-Rosenberg, L.; Brunström, B.; Dencker, L.; Kultima, K.; Scholz, B. Faced with inequality: Chicken do not have a general dosage compensation of sex-linked genes. BMC Boil. 2007, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Itoh, Y.; Melamed, E.; Yang, X.; Kampf, K.; Wang, S.S.; Yehya, N.; Van Nas, A.; Replogle, K.; Band, M.; Clayton, D.F.; et al. Dosage compensation is less effective in birds than in mammals. J. Boil. 2007, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- E Mank, J.; Ellegren, H. All dosage compensation is local: Gene-by-gene regulation of sex-biased expression on the chicken Z chromosome. Heredity 2008, 102, 312–320. [Google Scholar] [CrossRef] [Green Version]
- Melamed, E.; Arnold, A.P. The role of LINEs and CpG islands in dosage compensation on the chicken Z chromosome. Chromosom. Res. 2009, 17, 727–736. [Google Scholar] [CrossRef] [Green Version]
- Melamed, E.; Elashoff, D.; Arnold, A.P. Evaluating dosage compensation on the chicken Z chromosome: Should effective dosage compensation eliminate sexual bias? Heredity 2009, 103, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, F.; Harrison, P.W.; Dessimoz, C.; Mank, J.E. Compensation of Dosage-Sensitive Genes on the Chicken Z Chromosome. Genome Boil. Evol. 2016, 8, 1233–1242. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Maney, D.L.; Layman, T.S.; Chatterjee, P.; Yi, S.V. Regional epigenetic differentiation of the Z Chromosome between sexes in a female heterogametic system. Genome Res. 2019, 29, 1673–1684. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Zhou, Q. Female-specific and dosage selections restore genes through transpositions onto the degenerated songbird W chromosomes. bioRxiv 2019, 692194. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Li, C.; Li, Q.; Li, B.; Larkin, D.M.; Lee, C.; Storz, J.F.; Antunes, A.; Greenwold, M.J.; Meredith, R.W.; et al. Comparative genomics reveals insights into avian genome evolution and adaptation. Science 2014, 346, 1311–1320. [Google Scholar] [CrossRef] [Green Version]
- International Chicken Genome. Sequencing Consortium Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef]
- Suh, A. The Specific Requirements for CR1 Retrotransposition Explain the Scarcity of Retrogenes in Birds. J. Mol. Evol. 2015, 81, 18–20. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Zhou, Q. The Female-Specific W Chromosomes of Birds Have Conserved Gene Contents but Are Not Feminized. Genes 2020, 11, 1126. https://doi.org/10.3390/genes11101126
Xu L, Zhou Q. The Female-Specific W Chromosomes of Birds Have Conserved Gene Contents but Are Not Feminized. Genes. 2020; 11(10):1126. https://doi.org/10.3390/genes11101126
Chicago/Turabian StyleXu, Luohao, and Qi Zhou. 2020. "The Female-Specific W Chromosomes of Birds Have Conserved Gene Contents but Are Not Feminized" Genes 11, no. 10: 1126. https://doi.org/10.3390/genes11101126
APA StyleXu, L., & Zhou, Q. (2020). The Female-Specific W Chromosomes of Birds Have Conserved Gene Contents but Are Not Feminized. Genes, 11(10), 1126. https://doi.org/10.3390/genes11101126