Induction of Chicken Host Defense Peptides within Disease-Resistant and -Susceptible Lines


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Cell Culture
2.2.1. Chicken Embryonic Fibroblast (CEF) Culture
2.2.2. Bone Marrow-Derived Cell (BMC) Culture
2.3. Immune Stimulation
2.3.1. Lipopolysaccharide (LPS) Treatment
2.3.2. Polyinosinic: Polycytidylic Acid (Poly(I:C)) Treatment
2.4. Analysis of Gene Expression
2.4.1. Primer Design
2.4.2. Expression Analysis of Host Defense Peptide Genes
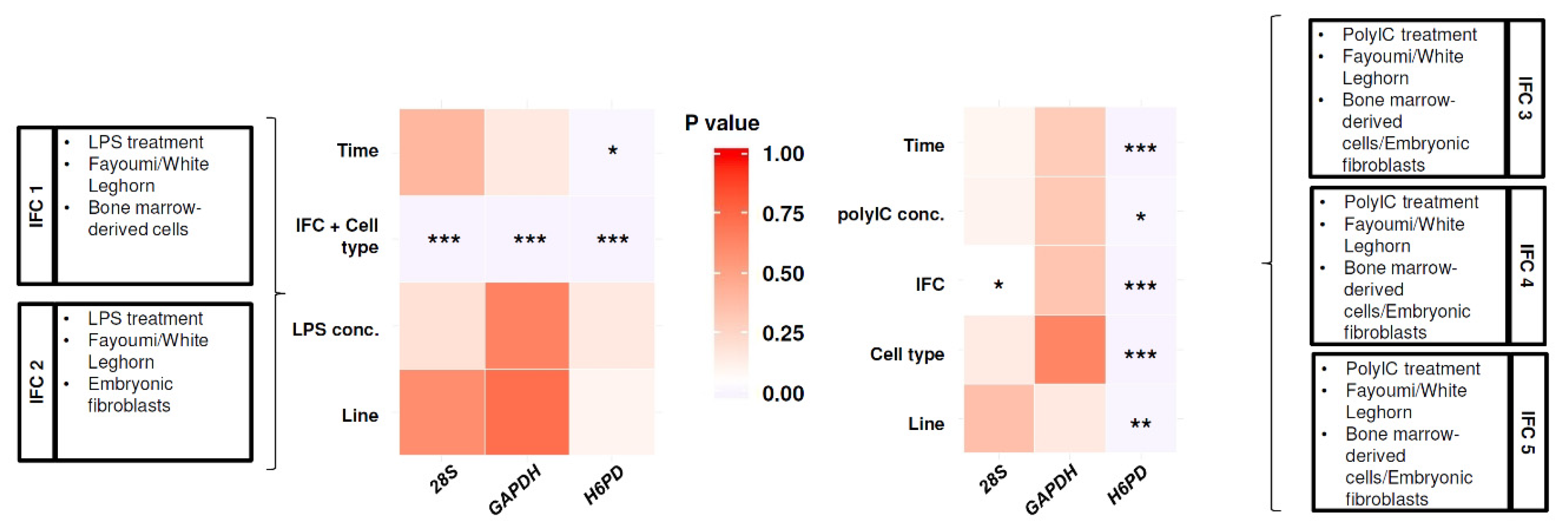
2.4.3. Data Imputation and Reference Gene Selection
2.5. Statistical Analysis
3. Results
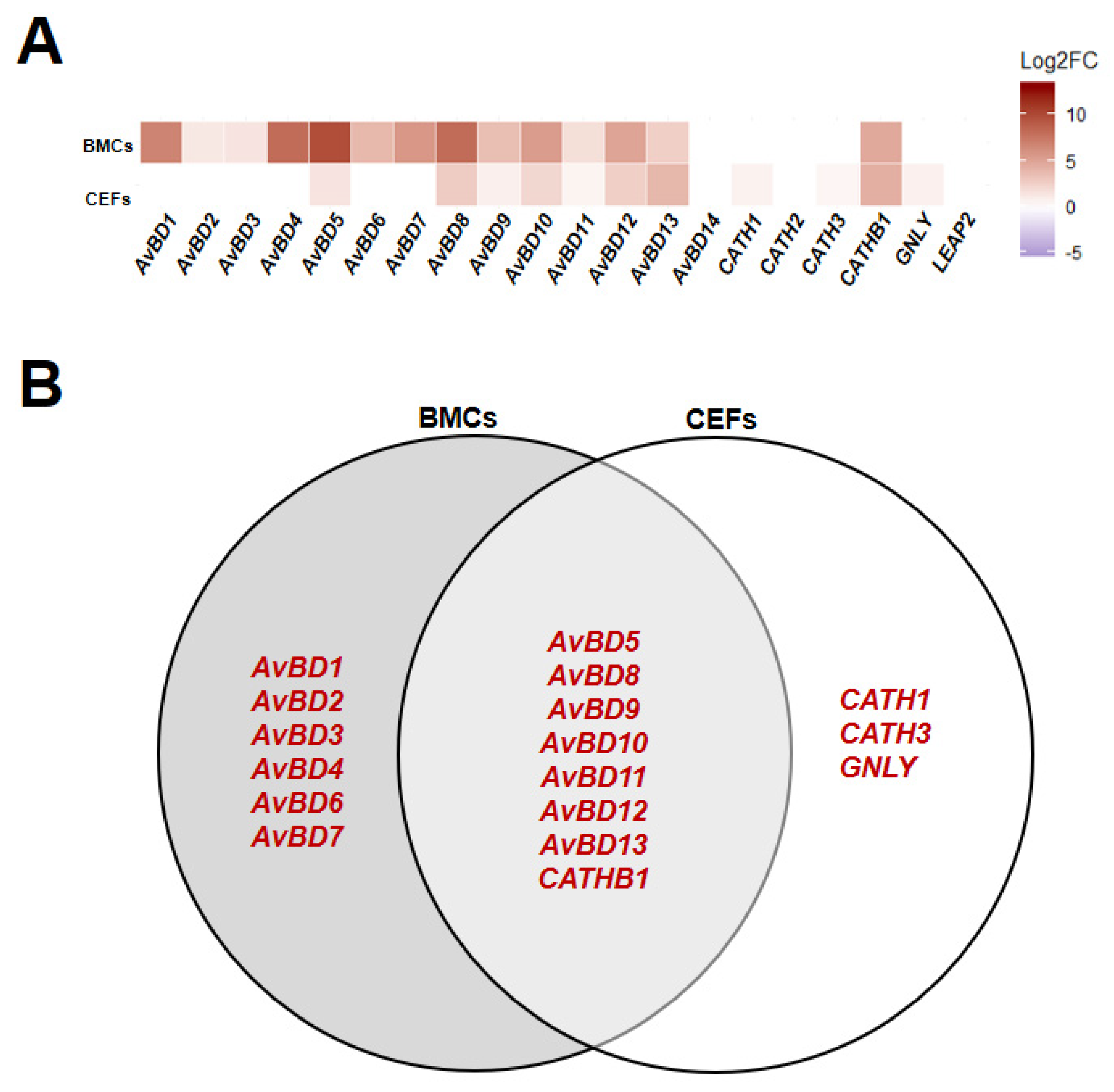
3.1. Line Differences in Expression of HDPs in Homeostatic State
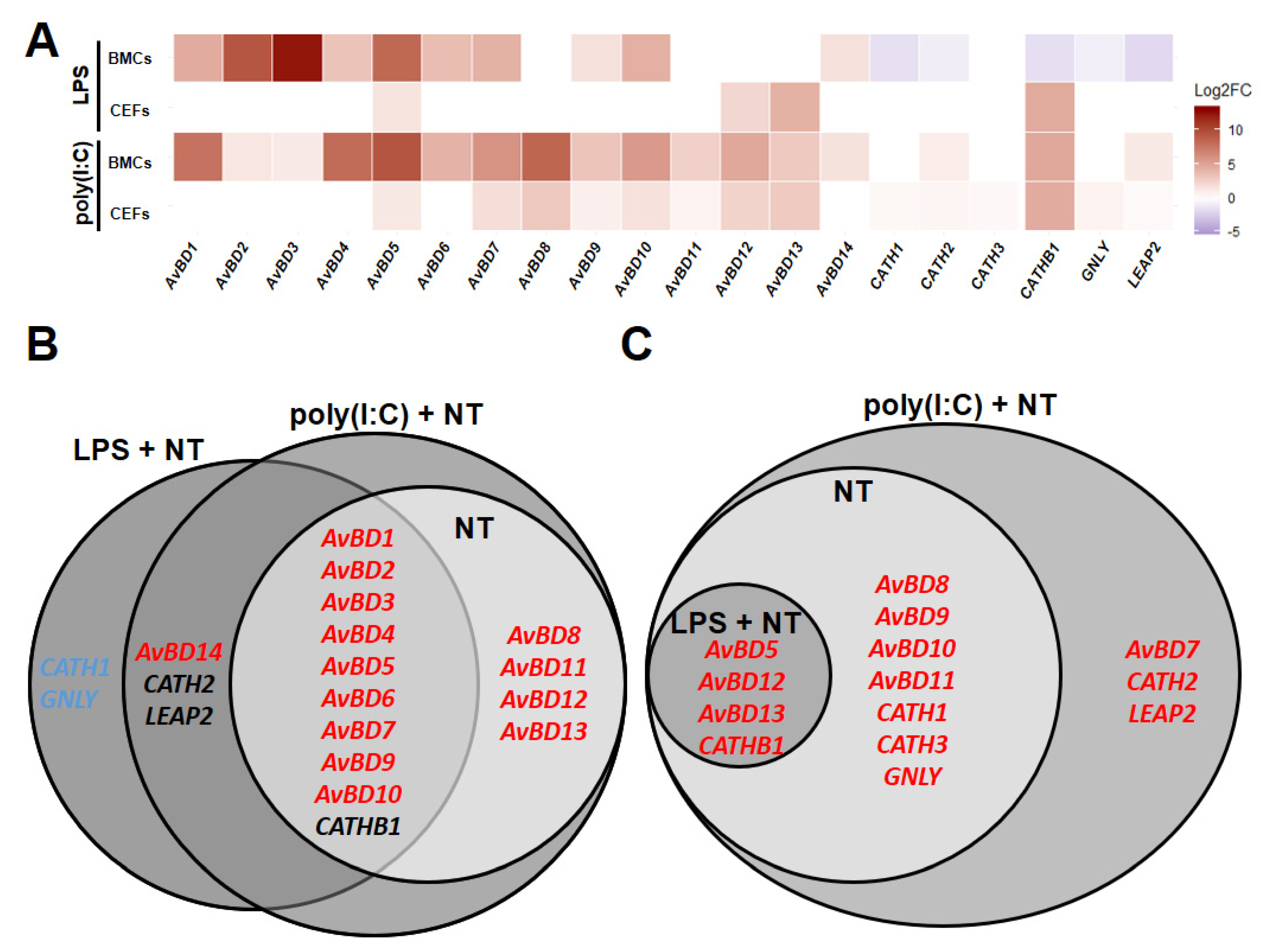
3.2. HDPs Responded to LPS and Poly(I:C)
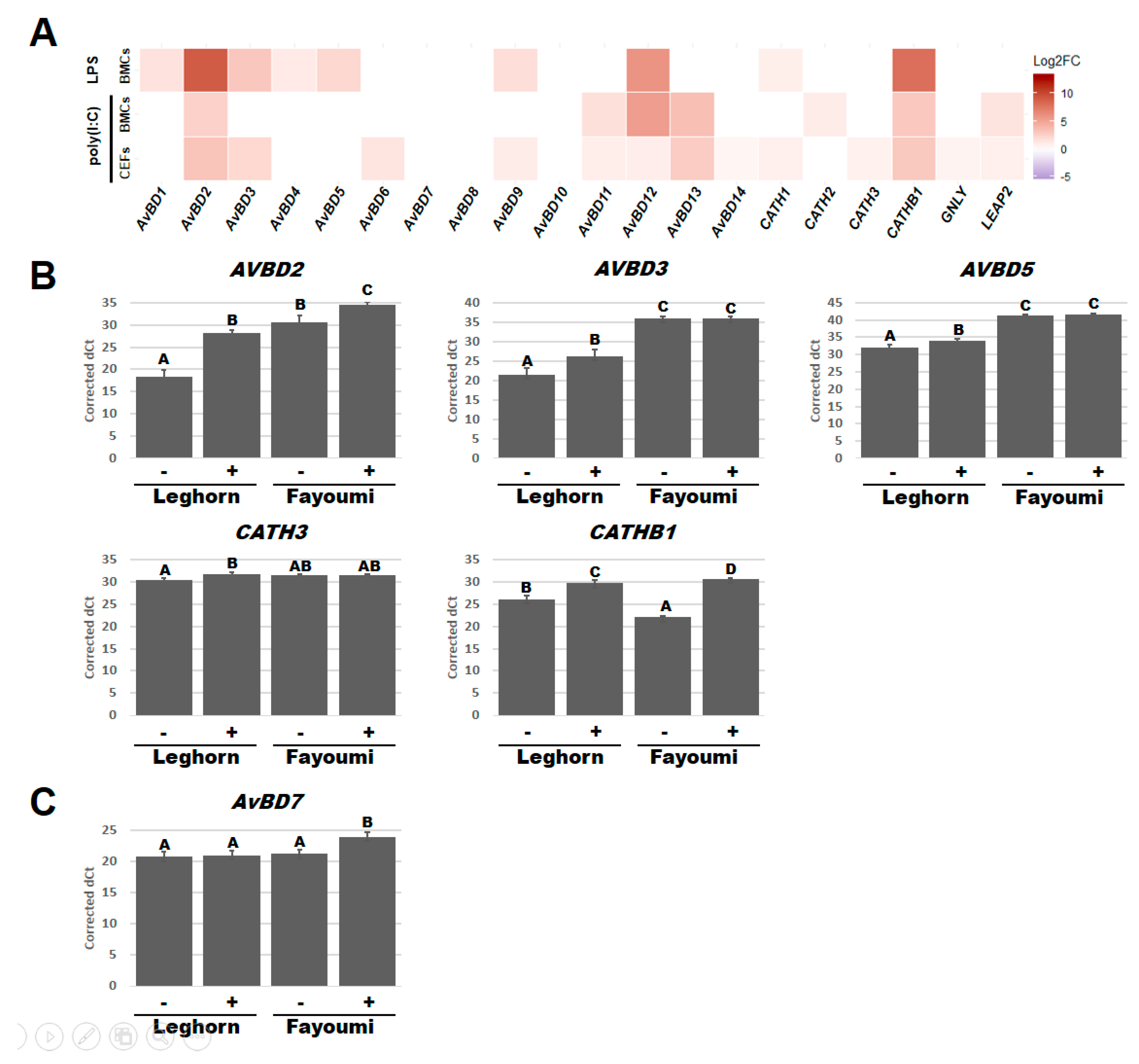
3.3. Line-Dependent Expression of HDPs Impacted by LPS and Poly(I:C) Treatments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jenssen, H.; Hamill, P.; Hancock, R.E. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosciuczuk, E.M.; Lisowski, P.; Jarczak, J.; Strzalkowska, N.; Jozwik, A.; Horbanczuk, J.; Krzyzewski, J.; Zwierzchowski, L.; Bagnicka, E. Cathelicidins: Family of antimicrobial peptides. A review. Mol. Biol. Rep. 2012, 39, 10957–10970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starr, C.G.; Maderdrut, J.L.; He, J.; Coy, D.H.; Wimley, W.C. Pituitary adenylate cyclase-activating polypeptide is a potent broad-spectrum antimicrobial peptide: Structure-activity relationships. Peptides 2018, 104, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Zanetti, M. Cathelicidins, multifunctional peptides of the innate immunity. J. Leukoc. Biol. 2004, 75, 39–48. [Google Scholar] [CrossRef]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Moravej, H.; Moravej, Z.; Yazdanparast, M.; Heiat, M.; Mirhosseini, A.; Moosazadeh Moghaddam, M.; Mirnejad, R. Antimicrobial Peptides: Features, Action, and Their Resistance Mechanisms in Bacteria. Microb. Drug Resist. 2018, 24, 747–767. [Google Scholar] [CrossRef]
- Taniguchi, M.; Saito, K.; Aida, R.; Ochiai, A.; Saitoh, E.; Tanaka, T. Wound healing activity and mechanism of action of antimicrobial and lipopolysaccharide-neutralizing peptides from enzymatic hydrolysates of rice bran proteins. J. Biosci. Bioeng. 2019, 128, 142–148. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial Peptides of Multicellular Organisms: My Perspective. Adv. Exp. Med. Biol. 2019, 1117, 3–6. [Google Scholar] [CrossRef]
- Cuperus, T.; Coorens, M.; van Dijk, A.; Haagsman, H.P. Avian host defense peptides. Dev. Comp. Immunol. 2013, 41, 352–369. [Google Scholar] [CrossRef]
- Lee, M.O.; Jang, H.J.; Rengaraj, D.; Yang, S.Y.; Han, J.Y.; Lamont, S.J.; Womack, J.E. Tissue expression and antibacterial activity of host defense peptides in chicken. BMC Vet. Res. 2016, 12, 231. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.O.; Kim, E.H.; Jang, H.J.; Park, M.N.; Woo, H.J.; Han, J.Y.; Womack, J.E. Effects of a single nucleotide polymorphism in the chicken NK-lysin gene on antimicrobial activity and cytotoxicity of cancer cells. Proc. Natl. Acad. Sci. USA 2012, 109, 12087–12092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynn, D.J.; Lloyd, A.T.; O’Farrelly, C. In silico identification of components of the Toll-like receptor (TLR) signaling pathway in clustered chicken expressed sequence tags (ESTs). Vet. Immunol. Immunopathol. 2003, 93, 177–184. [Google Scholar] [CrossRef]
- van Dijk, A.; Molhoek, E.M.; Bikker, F.J.; Yu, P.L.; Veldhuizen, E.J.; Haagsman, H.P. Avian cathelicidins: Paradigms for the development of anti-infectives. Vet. Microbiol. 2011, 153, 27–36. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, A.; Veldhuizen, E.J.; Kalkhove, S.I.; Tjeerdsma-van Bokhoven, J.L.; Romijn, R.A.; Haagsman, H.P. The β-defensin gallinacin-6 is expressed in the chicken digestive tract and has antimicrobial activity against food-borne pathogens. Antimicrob. Agents Chemother. 2007, 51, 912–922. [Google Scholar] [CrossRef] [Green Version]
- Selsted, M.E.; Ouellette, A.J. Mammalian defensins in the antimicrobial immune response. Nat. Immunol. 2005, 6, 551–557. [Google Scholar] [CrossRef]
- Xiao, Y.; Hughes, A.L.; Ando, J.; Matsuda, Y.; Cheng, J.F.; Skinner-Noble, D.; Zhang, G. A genome-wide screen identifies a single β-defensin gene cluster in the chicken: Implications for the origin and evolution of mammalian defensins. BMC Genom. 2004, 5, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Biragyn, A.; Hoover, D.M.; Lubkowski, J.; Oppenheim, J.J. Multiple roles of antimicrobial defensins, cathelicidins, and eosinophil-derived neurotoxin in host defense. Annu. Rev. Immunol. 2004, 22, 181–215. [Google Scholar] [CrossRef]
- Hellgren, O.; Ekblom, R. Evolution of a cluster of innate immune genes (β-defensins) along the ancestral lines of chicken and zebra finch. Immunome Res. 2010, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Gong, D.; Wilson, P.W.; Bain, M.M.; McDade, K.; Kalina, J.; Herve-Grepinet, V.; Nys, Y.; Dunn, I.C. Gallin; an antimicrobial peptide member of a new avian defensin family, the ovodefensins, has been subject to recent gene duplication. BMC Immunol. 2010, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Brockus, C.W.; Jackwood, M.W.; Harmon, B.G. Characterization of β-defensin prepropeptide mRNA from chicken and turkey bone marrow. Anim. Genet. 1998, 29, 283–289. [Google Scholar] [CrossRef]
- Harwig, S.S.; Swiderek, K.M.; Kokryakov, V.N.; Tan, L.; Lee, T.D.; Panyutich, E.A.; Aleshina, G.M.; Shamova, O.V.; Lehrer, R.I. Gallinacins: Cysteine-rich antimicrobial peptides of chicken leukocytes. FEBS Lett. 1994, 342, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, A.; Veldhuizen, E.J.; van Asten, A.J.; Haagsman, H.P. CMAP27, a novel chicken cathelicidin-like antimicrobial protein. Vet. Immunol. Immunopathol. 2005, 106, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Cai, Y.; Bommineni, Y.R.; Fernando, S.C.; Prakash, O.; Gilliland, S.E.; Zhang, G. Identification and functional characterization of three chicken cathelicidins with potent antimicrobial activity. J. Biol. Chem. 2006, 281, 2858–2867. [Google Scholar] [CrossRef] [Green Version]
- Achanta, M.; Sunkara, L.T.; Dai, G.; Bommineni, Y.R.; Jiang, W.; Zhang, G. Tissue expression and developmental regulation of chicken cathelicidin antimicrobial peptides. J. Anim. Sci. Biotechnol. 2012, 3, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goitsuka, R.; Chen, C.L.; Benyon, L.; Asano, Y.; Kitamura, D.; Cooper, M.D. Chicken cathelicidin-B1, an antimicrobial guardian at the mucosal M cell gateway. Proc. Natl. Acad. Sci. USA 2007, 104, 15063–15068. [Google Scholar] [CrossRef] [Green Version]
- Lynn, D.J.; Higgs, R.; Gaines, S.; Tierney, J.; James, T.; Lloyd, A.T.; Fares, M.A.; Mulcahy, G.; O’Farrelly, C. Bioinformatic discovery and initial characterisation of nine novel antimicrobial peptide genes in the chicken. Immunogenetics 2004, 56, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Michailidis, G.; Avdi, M.; Argiriou, A. Transcriptional profiling of antimicrobial peptides avian β-defensins in the chicken ovary during sexual maturation and in response to Salmonella enteritidis infection. Res. Vet. Sci. 2012, 92, 60–65. [Google Scholar] [CrossRef]
- Deist, M.S.; Gallardo, R.A.; Bunn, D.A.; Kelly, T.R.; Dekkers, J.C.M.; Zhou, H.; Lamont, S.J. Novel Mechanisms Revealed in the Trachea Transcriptome of Resistant and Susceptible Chicken Lines following Infection with Newcastle Disease Virus. Clin. Vaccine Immunol. 2017, 24. [Google Scholar] [CrossRef]
- Kim, D.K.; Kim, C.H.; Lamont, S.J.; Keeler, C.L., Jr.; Lillehoj, H.S. Gene expression profiles of two B-complex disparate, genetically inbred Fayoumi chicken lines that differ in susceptibility to Eimeria maxima. Poult. Sci. 2009, 88, 1565–1579. [Google Scholar] [CrossRef]
- Schilling, M.A.; Memari, S.; Cavanaugh, M.; Katani, R.; Deist, M.S.; Radzio-Basu, J.; Lamont, S.J.; Buza, J.J.; Kapur, V. Conserved, breed-dependent, and subline-dependent innate immune responses of Fayoumi and Leghorn chicken embryos to Newcastle disease virus infection. Sci. Rep. 2019, 9, 7209. [Google Scholar] [CrossRef]
- Zhou, H.; Lamont, S.J. Chicken MHC class I and II gene effects on antibody response kinetics in adult chickens. Immunogenetics 2003, 55, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Saelao, P.; Wang, Y.; Gallardo, R.A.; Lamont, S.J.; Dekkers, J.M.; Kelly, T.; Zhou, H. Novel insights into the host immune response of chicken Harderian gland tissue during Newcastle disease virus infection and heat treatment. BMC Vet. Res. 2018, 14, 280. [Google Scholar] [CrossRef] [PubMed]
- Van Goor, A.; Slawinska, A.; Schmidt, C.J.; Lamont, S.J. Distinct functional responses to stressors of bone marrow derived dendritic cells from diverse inbred chicken lines. Dev. Comp. Immunol. 2016, 63, 96–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.; Miska, K.B.; Fetterer, R.H.; Jenkins, M.C.; Lamont, S.J.; Wong, E.A. Differential expression of intestinal nutrient transporters and host defense peptides in Eimeria maxima-infected Fayoumi and Ross chickens. Poult. Sci. 2018, 97, 4392–4400. [Google Scholar] [CrossRef]
- Hasenstein, J.R.; Lamont, S.J. Chicken gallinacin gene cluster associated with Salmonella response in advanced intercross line. Avian Dis. 2007, 51, 561–567. [Google Scholar] [CrossRef]
- Jankovic, B.D.; Isakovic, K.; Petrovic, S. Direct stimulation of lymphoid tissue of the chicken. 3. Haemagglutinin production, haemolysin-forming cells and changes in lymphoid tissues following injection of guinea-pig erythrocytes into the bone marrow. Immunology 1973, 25, 663–674. [Google Scholar]
- Vainio, O.; Imhof, B.A. The immunology and developmental biology of the chicken. Immunol. Today 1995, 16, 365–370. [Google Scholar] [CrossRef]
- Sun, H.; Bi, R.; Liu, P.; Nolan, L.K.; Lamont, S.J. Combined analysis of primary lymphoid tissues’ transcriptomic response to extra-intestinal Escherichia coli (ExPEC) infection. Dev. Comp. Immunol. 2016, 57, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Wong, T.; McGrath, J.A.; Navsaria, H. The role of fibroblasts in tissue engineering and regeneration. Br. J. Dermatol. 2007, 156, 1149–1155. [Google Scholar] [CrossRef]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell; Garland Science: New York, NY, USA, 2015; pp. 1227–1242. [Google Scholar]
- Steinhauser, M.L.; Kunkel, S.L.; Hogaboam, C.M.; Evanoff, H.; Strieter, R.M.; Lukacs, N.W. Macrophage/fibroblast coculture induces macrophage inflammatory protein-1alpha production mediated by intercellular adhesion molecule-1 and oxygen radicals. J. Leukoc. Biol. 1998, 64, 636–641. [Google Scholar] [CrossRef] [Green Version]
- Blenis, J.; Hawkes, S.P. Transformation-sensitive protein associated with the cell substratum of chicken embryo fibroblasts. Proc. Natl. Acad. Sci. USA 1983, 80, 770–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arslan, M.; Yang, X.; Santhakumar, D.; Liu, X.; Hu, X.; Munir, M.; Li, Y.; Zhang, Z. Dynamic Expression of Interferon Lambda Regulated Genes in Primary Fibroblasts and Immune Organs of the Chicken. Genes 2019, 10, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szmolka, A.; Wiener, Z.; Matulova, M.E.; Varmuzova, K.; Rychlik, I. Gene Expression Profiles of Chicken Embryo Fibroblasts in Response to Salmonella Enteritidis Infection. PLoS ONE 2015, 10, e0127708. [Google Scholar] [CrossRef] [PubMed]
- Rengaraj, D.; Truong, A.D.; Lillehoj, H.S.; Han, J.Y.; Hong, Y.H. Expression and regulation of avian β-defensin 8 protein in immune tissues and cell lines of chickens. Asian Australas. J. Anim. Sci. 2018, 31, 1516–1524. [Google Scholar] [CrossRef]
- Calabrese, V.; Cighetti, R.; Peri, F. Molecular simplification of lipid A structure: TLR4-modulating cationic and anionic amphiphiles. Mol. Immunol. 2015, 63, 153–161. [Google Scholar] [CrossRef]
- Fortier, M.E.; Kent, S.; Ashdown, H.; Poole, S.; Boksa, P.; Luheshi, G.N. The viral mimic, polyinosinic:polycytidylic acid, induces fever in rats via an interleukin-1-dependent mechanism. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R759–R766. [Google Scholar] [CrossRef] [Green Version]
- Imanishi, T.; Ishihara, C.; Badr, M.E.G.S.; Hashimoto-Tane, A.; Kimura, Y.; Kawai, T.; Takeuchi, O.; Ishii, K.J.; Taniguchi, S.; Noda, T.; et al. Nucleic acid sensing by T cells initiates Th2 cell differentiation. Nat. Commun. 2014, 5, 3566. [Google Scholar] [CrossRef] [Green Version]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. TLR signaling. Cell Death Differ. 2006, 13, 816–825. [Google Scholar] [CrossRef] [Green Version]
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef]
- Opitz, B.; van Laak, V.; Eitel, J.; Suttorp, N. Innate immune recognition in infectious and noninfectious diseases of the lung. Am. J. Respir. Crit. Care Med. 2010, 181, 1294–1309. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.L.; Brooks, C.; Douwes, J. Innate immunity in asthma. Paediatr. Respir. Rev. 2008, 9, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Song, K.D. Expression patterns of innate immunity-related genes in response to polyinosinic:polycytidylic acid (poly[I:C]) stimulation in DF-1 chicken fibroblast cells. J. Anim. Sci. Technol. 2020, 62, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Kint, J.; Fernandez-Gutierrez, M.; Maier, H.J.; Britton, P.; Langereis, M.A.; Koumans, J.; Wiegertjes, G.F.; Forlenza, M. Activation of the chicken type I interferon response by infectious bronchitis coronavirus. J. Virol. 2015, 89, 1156–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michailidis, G.; Anastasiadou, M.; Guibert, E.; Froment, P. Activation of innate immune system in response to lipopolysaccharide in chicken Sertoli cells. Reproduction 2014, 148, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Slawinska, A.; Hsieh, J.C.; Schmidt, C.J.; Lamont, S.J. Heat Stress and Lipopolysaccharide Stimulation of Chicken Macrophage-Like Cell Line Activates Expression of Distinct Sets of Genes. PLoS ONE 2016, 11, e0164575. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Deist, M.S.; Gallardo, R.A.; Bunn, D.A.; Kelly, T.R.; Dekkers, J.C.M.; Zhou, H.; Lamont, S.J. Novel analysis of the Harderian gland transcriptome response to Newcastle disease virus in two inbred chicken lines. Sci. Rep. 2018, 8, 6558. [Google Scholar] [CrossRef] [Green Version]
- Monson, M.S.; Van Goor, A.G.; Ashwell, C.M.; Persia, M.E.; Rothschild, M.F.; Schmidt, C.J.; Lamont, S.J. Immunomodulatory effects of heat stress and lipopolysaccharide on the bursal transcriptome in two distinct chicken lines. BMC Genom. 2018, 19, 643. [Google Scholar] [CrossRef]
- Monson, M.S.; Van Goor, A.G.; Persia, M.E.; Rothschild, M.F.; Schmidt, C.J.; Lamont, S.J. Genetic lines respond uniquely within the chicken thymic transcriptome to acute heat stress and low dose lipopolysaccharide. Sci. Rep. 2019, 9, 13649. [Google Scholar] [CrossRef] [Green Version]
- Haney, E.F.; Mansour, S.C.; Hancock, R.E. Antimicrobial Peptides: An Introduction. Methods Mol. Biol. 2017, 1548, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Santiago, B.; Serrano, C.J.; Enciso-Moreno, J.A. Susceptibility to infectious diseases based on antimicrobial peptide production. Infect. Immun. 2009, 77, 4690–4695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, P.; Bajaj-Elliott, M.; Katubulushi, M.; Zulu, I.; Poulsom, R.; Feldman, R.A.; Bevins, C.L.; Dhaliwal, W. Reduced gene expression of intestinal α-defensins predicts diarrhea in a cohort of African adults. J. Infect. Dis. 2006, 193, 1464–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zasloff, M. Innate immunity, antimicrobial peptides, and protection of the oral cavity. Lancet 2002, 360, 1116–1117. [Google Scholar] [CrossRef]
- Derache, C.; Labas, V.; Aucagne, V.; Meudal, H.; Landon, C.; Delmas, A.F.; Magallon, T.; Lalmanach, A.C. Primary structure and antibacterial activity of chicken bone marrow-derived β-defensins. Antimicrob. Agents Chemother. 2009, 53, 4647–4655. [Google Scholar] [CrossRef] [Green Version]
- Lynn, D.J.; Higgs, R.; Lloyd, A.T.; O’Farrelly, C.; Herve-Grepinet, V.; Nys, Y.; Brinkman, F.S.; Yu, P.L.; Soulier, A.; Kaiser, P.; et al. Avian β-defensin nomenclature: A community proposed update. Immunol. Lett. 2007, 110, 86–89. [Google Scholar] [CrossRef]
- van Dijk, A.; Veldhuizen, E.J.; Haagsman, H.P. Avian defensins. Vet. Immunol. Immunopathol. 2008, 124, 1–18. [Google Scholar] [CrossRef]
- Zhang, G.; Sunkara, L.T. Avian antimicrobial host defense peptides: From biology to therapeutic applications. Pharmaceuticals 2014, 7, 220–247. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Entrez ID | Forward Primer (5’–3’) | Reverse Primer (5’–3’) | Amplicon Length (bp) |
|---|---|---|---|---|
| AvBD1 | 395841 | ACAGACGTAAACCATGCGGA | GACTTCCTTCCTAGAGCCTGG | 99 |
| AvBD2 | 395840 | GGGTGTCCCAGCCATCTAAT | TCCAAGGCCATTTGCAGCAG | 70 |
| AvBD3 | 395363 | CTCTTGTTTCTCCAGGGTGCT | GCTCCCAACACGACAGAATC | 75 |
| AvBD4 | 414342 | CAGTCTGCCTTCTGCCATGA | GGTCCCGCGATATCCACATT | 124 |
| AvBD5 | 414340 | GTCATGTCCTCCAGGGATCG | CGTGAAGGGACATCAGAGGC | 101 |
| AvBD6 | 407776 | GGGTTGGATCATGTGGCAGT | AGTGCCAGAGAGGCCATTTG | 120 |
| AvBD7 | 407777 | GGGATCTGTCGAAGGCCATA | TTCCCAGAAGTCAGGGAGGT | 105 |
| AvBD8 | 414875 | CTTGGCCGTTCTCCTCACTG | ACTGTGCCTCGTTGTTAGGT | 71 |
| AvBD9 | 414343 | ACACCGTCAGGCATCTTCAC | GTCTTCTTGGCTGTAAGCTGG | 129 |
| AvBD10 | 414341 | TCAGGGGAATTTCTGCCGTG | CTTACTGCGCCGGAATCTTG | 107 |
| AvBD11 | 414876 | ATGCTCTTGGCGTCAGAAAAC | GGAGATACGCAATGGCCCC | 87 |
| AvBD12 | 414339 | CTGCTCGCTCACGGAAGCA | TATTCCCCAGGGTTGCAGTTC | 89 |
| AvBD13 | 414877 | AGCTGCTCTTTGCCATCGTT | CAGTGGCCATGGTTGTTCCT | 97 |
| AvBD14 | 100858701 | GGCGACACGACAATGTCAAC | TTGCCCTTCATCTTCCGACA | 119 |
| CATH1 | 414337 | GACTCCATGGCTGACCCTGT | ATCGCCCGGTAGAGGTTGTA | 89 |
| CATH2 | 420407 | CGACTGCGACTTCAAGGAGAA | GATCTCGGGAGTGTCCTGC | 79 |
| CATH3 | 100858343 | GATGTCACCTGCGTGGACTC | TTGTAGAGGTTGATGCCCGC | 107 |
| CATHB1 | 100858412 | GGTTGCTCAACCAGAGGATCT | TCCTCCACAAGGAAGCTCAC | 118 |
| GNLY | 693257 | TTCTGCGTCAGTCTGGTGAA | AGATACTCCTCTGGCGCCTC | 118 |
| LEAP2 | 414338 | GTTGGAGCCTCATGTAGGGA | GAGGCCGTTCTAAGGAAGCAG | 80 |
| 28S | 112533599 | GGCGAAGCCAGAGGAAACT | GACGACCGATTTGCACGTC | 62 |
| GAPDH | 374193 | GCAGCAGGAACACTATAAAGGC | TTTGCCAGAGAGGACGGC | 100 |
| H6PD | 428188 | ATGTACCGGGTGGACCACTA | AACTGACGGTTCTGATCTCGAAA | 80 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, H.-J.; Monson, M.; Kaiser, M.; Lamont, S.J. Induction of Chicken Host Defense Peptides within Disease-Resistant and -Susceptible Lines. Genes 2020, 11, 1195. https://doi.org/10.3390/genes11101195
Jang H-J, Monson M, Kaiser M, Lamont SJ. Induction of Chicken Host Defense Peptides within Disease-Resistant and -Susceptible Lines. Genes. 2020; 11(10):1195. https://doi.org/10.3390/genes11101195
Chicago/Turabian StyleJang, Hyun-Jun, Melissa Monson, Michael Kaiser, and Susan J Lamont. 2020. "Induction of Chicken Host Defense Peptides within Disease-Resistant and -Susceptible Lines" Genes 11, no. 10: 1195. https://doi.org/10.3390/genes11101195
APA StyleJang, H. -J., Monson, M., Kaiser, M., & Lamont, S. J. (2020). Induction of Chicken Host Defense Peptides within Disease-Resistant and -Susceptible Lines. Genes, 11(10), 1195. https://doi.org/10.3390/genes11101195