Genetic Risk of Autism Spectrum Disorder in a Pakistani Population


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Procedure and Participants
2.2. DNA Extraction and Quantification
2.3. Genotyping
2.4. Genotyping Data Analysis
2.5. Protein–Protein Interaction Network
3. Results
3.1. Patient Characteristics and Minor Allele Fequencies
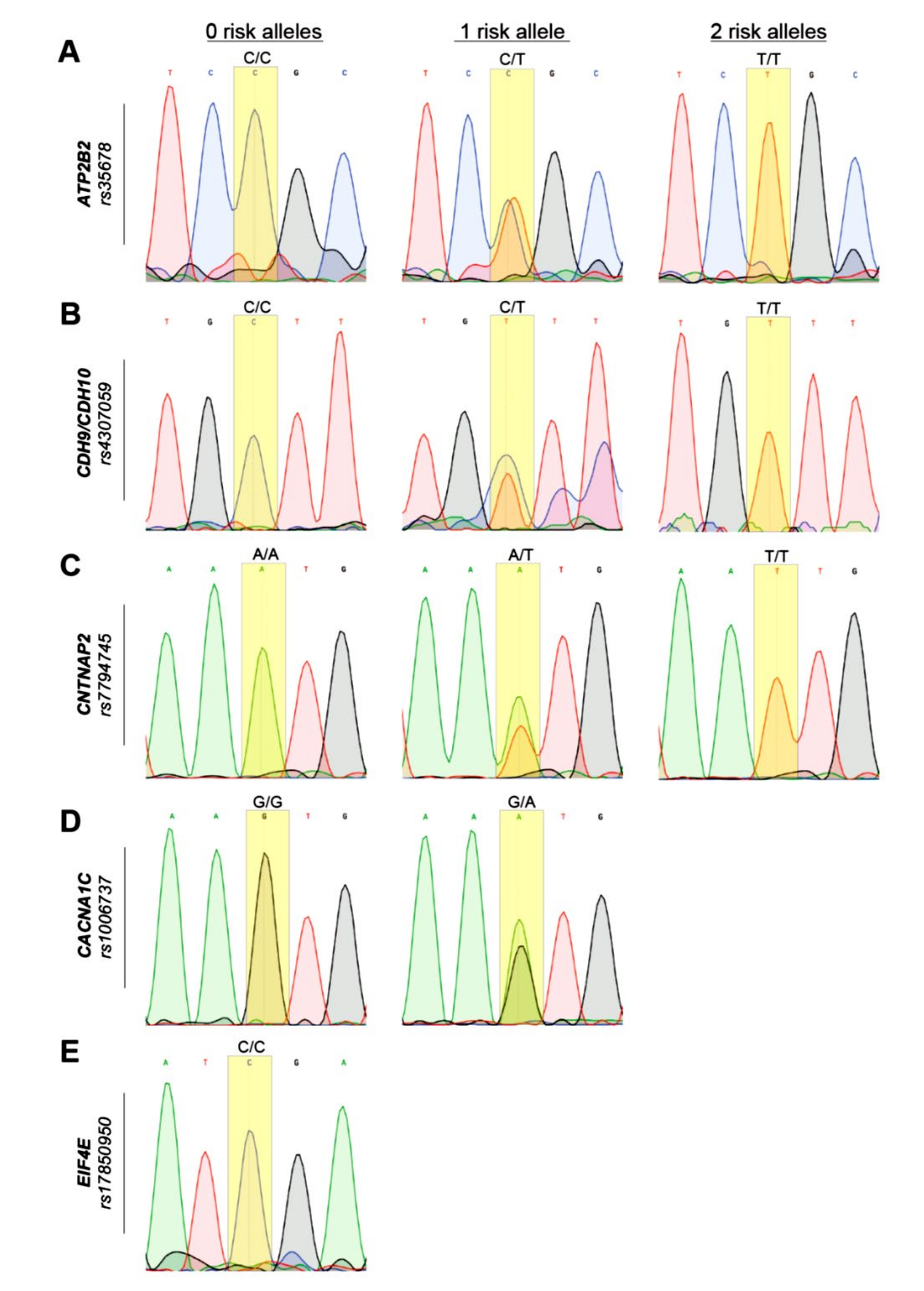
3.2. Genotyping and Sanger Sequencing
3.3. Comparison of Allele Frequencies of Studied Population with Global Allele Frequencies
3.4. Association Analysis
3.5. Risk Score Analysis
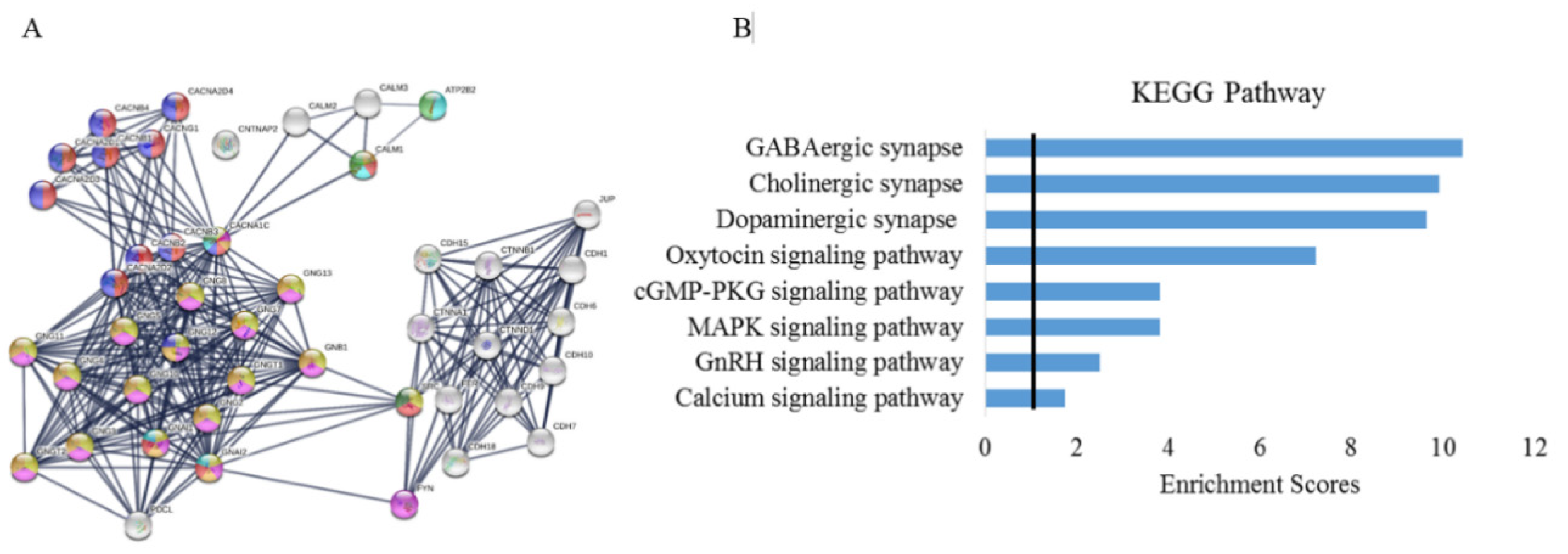
3.6. Protein–Protein Interactions and Pathway Analysis
3.7. Association between SNPs and Autistic-Like Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ASD | Autism spectrum disorder |
| SNP | Single nucleotide polymorphisms |
| GWAS | Genome wide association studies |
| OR | Odds’ ratio |
| CNTNAP2 | Contactin Association Protein-like 2 |
| CACNA1C | L-type voltage-dependent calcium channel |
| PCR | Polymerase chain reaction |
| DSM | Diagnostic and Statistical Manual |
| ADOS | Autism Diagnosis Observation Schedule |
| RBC | Red blood cells |
| WBC | White blood cells |
| KEGG | Kyoto encyclopedia of genes and genomes |
References
- Strock, M. Autism spectrum disorders (pervasive developmental disorders). Natl. Inst. Ment. Health 2007, 41. [Google Scholar] [CrossRef] [Green Version]
- Robert, C.; Pasquier, L.; Cohen, D.; Fradin, M.; Canitano, R.; Damaj, L.; Odent, S.; Tordjman, S. Role of genetics in the etiology of autistic spectrum disorder: Towards a hierarchical diagnostic strategy. Int. J. Mol. Sci. 2017, 18, 618. [Google Scholar] [CrossRef] [Green Version]
- AlSalehi, S.M.; Alhifthy, E.H. Autism Spectrum Disorder. In Clinical Child Neurology; Springer International Publishing: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Kawamura, Y.; Takahashi, O.; Ishii, T. Reevaluating the incidence of pervasive developmental disorders: Impact of elevated rates of detection through implementation of an integrated system of screening in Toyota, Japan. Psychiatry Clin. Neurosci. 2008, 62, 152–159. [Google Scholar] [CrossRef]
- Baird, G.; Simonoff, E.; Pickles, A.; Chandler, S.; Loucas, T.; Meldrum, D.; Charman, T. Prevalence of disorders of the autism spectrum in a population cohort of children in South Thames: The Special Needs and Autism Project (SNAP). Lancet 2006, 368, 210–215. [Google Scholar] [CrossRef]
- Sun, X.; Allison, C.; Auyeung, B.; Zhang, Z.; Matthews, F.E.; Baron-Cohen, S.; Brayne, C. Validation of existing diagnosis of autism in mainland China using standardised diagnostic instruments. Autism 2015, 19, 1010–1017. [Google Scholar] [CrossRef] [Green Version]
- Baio, J.; Wiggins, L.; Christensen, D.L.; Maenner, M.J.; Daniels, J.; Warren, Z.; Kurzius-Spencer, M.; Zahorodny, W.; Rosenberg, C.R.; White, T.; et al. Prevalence of Autism spectrum disorder among children aged 8 Years—Autism and developmental disabilities monitoring network, 11 Sites, United States 2014. MMWR Surveill. Summ. 2018. [Google Scholar] [CrossRef]
- Kim, Y.S.; Leventhal, B.L.; Koh, Y.-J.; Fombonne, E.; Laska, E.; Lim, E.-C.; Cheon, K.-A.; Kim, S.-J.; Kim, Y.-K.; Lee, H.; et al. Prevalence of Autism spectrum disorders in a total population sample. Am. J. Psychiatry 2011, 168, 904–912. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Allison, C.; Matthews, F.E.; Sharp, S.J.; Auyeung, B.; Baron-Cohen, S.; Brayne, C. Prevalence of Autism in mainland China, Hong Kong and Taiwan: A systematic review and meta-analysis. Mol. Autism 2013, 4, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werling, D.M.; Geschwind, D.H. Understanding sex bias in Autism spectrum disorder. Curr. Opin. Neurol. 2013, 26, 146–153. [Google Scholar] [CrossRef] [Green Version]
- Mandell, D.S.; Novak, M.M.; Zubritsky, C.D. Spectrum disorders factors associated with age of diagnosis among children with Autism factors associated with age of diagnosis among children with Autism spectrum disorders. Pediatrics 2005, 116, 1480–1486. [Google Scholar] [CrossRef] [Green Version]
- Norbury, C.F.; Sparks, A. Difference or disorder? Cultural issues in understanding neurodevelopmental disorders. Dev. Psychol. 2013, 49, 45–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandin, S.; Lichtenstein, P.; Kuja-Halkola, R.; Hultman, C.; Larsson, H.; Reichenberg, A. The heritability of Autism spectrum disorder. JAMA J. Am. Med. Assoc. 2017, 318, 1182–1184. [Google Scholar] [CrossRef] [PubMed]
- Veenstra-Vanderweele, J.; Christian, S.L.; Cook, E.H., Jr. Autism as a paradigmatic complex genetic disorder. Annu. Rev. Genomics Hum. Genet. 2004, 5, 379–405. [Google Scholar] [CrossRef] [Green Version]
- Zoghbi, H.Y.; Bear, M.F. Synaptic dysfunction in neurodevelopmental disorders associated with Autism and Intellectual Disabilities. Cold Spring Harb. Perspect. Biol. 2012, 4, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Geschwind, D.H.; Levitt, P. Autism spectrum disorders: Developmental disconnection syndromes. Curr. Opin. Neurobiol. 2007, 17, 103–111. [Google Scholar] [CrossRef]
- Arndt, T.L.; Stodgell, C.J.; Rodier, P.M. The teratology of autism. Int. J. Dev. Neurosci. 2005, 23, 189–199. [Google Scholar] [CrossRef]
- Libbey, J.E.; Sweeten, T.L.; McMahon, W.M.; Fujinami, R.S. Autistic disorder and viral infections. J. NeuroVirol. 2005, 11, 1–10. [Google Scholar] [CrossRef]
- Lloyd-Fox, S.; Blasi, A.; Elwell, C.E.; Charman, T.; Murphy, D.; Johnson, M.H. Reduced neural sensitivity to social stimuli in infants at risk for Autism. Proc. R. Soc. B Biol. Sci. 2013, 280, 20123026. [Google Scholar] [CrossRef] [Green Version]
- Gekas, C.; Dieterlen-Lièvre, F.; Orkin, S.H.; Mikkola, H.K.A. The placenta is a niche for hematopoietic stem cells. Dev. Cell 2005, 8, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Grove, J.; Ripke, S.; Als, T.D.; Mattheisen, M.; Walters, R.K.; Won, H.; Pallesen, J.; Agerbo, E.; Andreassen, O.A.; Anney, R.; et al. Identification of common genetic risk variants for Autism spectrum disorder. Nat. Genet. 2019, 51, 431–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Gonzalez, A.; Calaza, M.; Rodriguez-Fontenla, C.; Carracedo, A. Novel gene-based analysis of ASD GWAS: Insight into the biological role of associated genes. Front. Genet. 2019, 10, 733. [Google Scholar] [CrossRef] [Green Version]
- The Autism Spectrum Disorders Working Group of The Psychiatric Genomics Consortium. Meta-analysis of GWAS of over 16,000 individuals with Autism spectrum disorder highlights a novel locus at 10q24.32 and a significant overlap with Schizophrenia. Mol. Autism 2017, 8, 1–17. [Google Scholar] [CrossRef]
- Daghsni, M.; Rima, M.; Fajloun, Z.; Ronjat, M.; Brusés, J.L.; M’rad, R.; De Waard, M. Autism throughout genetics: Perusal of the implication of ion channels. Brain Behav. 2018, 8, e00978. [Google Scholar] [CrossRef]
- Yang, W.; Liu, J.; Zheng, F.; Jia, M.; Zhao, L.; Lu, T.; Ruan, Y.; Zhang, J.; Yue, W.; Zhang, D.; et al. The evidence for association of ATP2B2 polymorphisms with Autism in Chinese Han population. PLoS ONE 2013, 8, e61021. [Google Scholar] [CrossRef] [Green Version]
- Minich, R.R.; Li, J.; Tempel, B.L. Early growth response protein 1 regulates promoter activity of α-plasma membrane calcium ATPase 2, a major calcium pump in the brain and auditory system. BMC Mol. Biol. 2017, 18, 14. [Google Scholar] [CrossRef] [Green Version]
- Takata, A.; Miyake, N.; Tsurusaki, Y.; Fukai, R.; Miyatake, S.; Koshimizu, E.; Kushima, I.; Okada, T.; Morikawa, M.; Uno, Y.; et al. Integrative analyses of de novo mutations provide deeper biological insights into Autism spectrum disorder. Cell Rep. 2018, 22, 734–747. [Google Scholar] [CrossRef] [Green Version]
- Carayol, J.; Sacco, R.; Tores, F.; Rousseau, F.; Lewin, P.; Hager, J.; Persico, A.M. Converging evidence for an association of ATP2B2 allelic variants with Autism in male subjects. Biol. Psychiatry 2011, 70, 880–887. [Google Scholar] [CrossRef]
- Prandini, P.; Pasquali, A.; Malerba, G.; Marostica, A.; Zusi, C.; Xumerle, L.; Muglia, P.; Da Ros, L.; Ratti, E.; Trabetti, E.; et al. The association of Rs4307059 and Rs35678 markers with Autism spectrum disorders is replicated in Italian families. Psychiatr. Genet. 2012, 22, 177–181. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, H.; Ma, D.; Bucan, M.; Glessner, J.T.; Abrahams, B.S.; Salyakina, D.; Imielinski, M.; Bradfield, J.P.; Sleiman, P.M.A.; et al. Common genetic variants on 5p14.1 associate with Autism spectrum disorders. Nature 2009, 459, 528–533. [Google Scholar] [CrossRef]
- Glessner, J.T.; Wang, K.; Cai, G.; Korvatska, O.; Kim, C.E.; Wood, S.; Zhang, H.; Estes, A.; Brune, C.W.; Bradfield, J.P.; et al. Autism genome-wide copy number variation reveals ubiquitin and neuronal genes. Nature 2009, 459, 569–573. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.; Lu, T.; Zhang, T.; Jia, M.; Ruan, Y.; Zhang, D.; Li, J.; Wang, L. Replication of previous GWAS hits suggests the association between Rs4307059 near MSNP1AS and Autism in a Chinese Han population. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2019, 92, 194–198. [Google Scholar] [CrossRef]
- De Witt, J.J.; Grepo, N.; Wilkinson, B.; Evgrafov, O.V.; Knowles, J.A.; Campbell, D.B. Impact of the Autism-associated long noncoding RNA MSNP1AS on neuronal architecture and gene expression in human neural progenitor cells. Genes 2016, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Canali, G.; Goutebroze, L. CNTNAP2 heterozygous missense variants: Risk factors for Autism spectrum disorder and/or other pathologies? J. Exp. Neurosci. 2018, 12, 1179069518809666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiebe, S.; Nagpal, A.; Truong, V.T.; Park, J.; Skalecka, A.; He, A.J.; Gamache, K.; Khoutorsky, A.; Gantois, I.; Sonenberg, N. Inhibitory interneurons mediate autism-associated behaviors via 4E-BP2. Proc. Natl. Acad. Sci. USA 2019, 116, 18060–18067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.C.; Cole, T.B.; Costa, L.G. Behavioral phenotyping for Autism spectrum disorders in mice. Curr. Protoc. Toxicol. 2017, 72, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.X.; Kim, G.H.; Tan, J.W.; Riso, A.E.; Sun, Y.; Xu, E.Y.; Liao, G.Y.; Xu, H.; Lee, S.H.; Do, N.Y.; et al. Elevated protein synthesis in microglia causes Autism-like synaptic and behavioral aberrations. Nat. Commun. 2020, 11, 1–17. [Google Scholar] [CrossRef]
- Zare, S.; Mashayekhi, F.; Bidabadi, E. The association of CNTNAP2 Rs7794745 gene polymorphism and Autism in Iranian population. J. Clin. Neurosci. 2017, 39, 189–192. [Google Scholar] [CrossRef]
- Beiranvandi, F.; Akouchekian, M.; Javadi, G.R.; Darvish, H. The association of CNTNAP2 Rs2710102 and ENGRAILED-2 Rs1861972 genes polymorphism and autism in Iranian population. Meta Gene 2020, 24, 100664. [Google Scholar] [CrossRef]
- Canali, G.; Garcia, M.; Hivert, B.; Pinatel, D.; Goullancourt, A.; Oguievetskaia, K.; Saint-Martin, M.; Girault, J.A.; Faivre-Sarrailh, C.; Goutebroze, L. Genetic variants in autism-related CNTNAP2 impair axonal growth of cortical neurons. Hum. Mol. Genet. 2018, 11, 1941–1954. [Google Scholar] [CrossRef] [Green Version]
- Liska, A.; Bertero, A.; Gomolka, R.; Sabbioni, M.; Galbusera, A.; Barsotti, N.; Panzeri, S.; Scattoni, M.L.; Pasqualetti, M.; Gozzi, A. Homozygous loss of Autism-risk gene CNTNAP2 results in reduced local and long-range prefrontal functional connectivity. Cereb. Cortex 2018, 28, 1141–1153. [Google Scholar] [CrossRef]
- Wei, Q.; Li, M.; Kang, Z.; Li, L.; Diao, F.; Zhang, R.; Wang, J.; Zheng, L.; Wen, X.; Zhang, J.; et al. ZNF804A Rs1344706 is associated with cortical thickness, surface area, and cortical volume of the unmedicated first episode Schizophrenia and healthy controls. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2015, 168, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Buddell, T.; Friedman, V.; Drozd, C.J.; Quinn, C.C. An Autism-causing calcium channel variant functions with selective autophagy to alter axon targeting and behavior. PLoS Genet. 2019, 12, e1008488. [Google Scholar] [CrossRef] [Green Version]
- Liao, X.; Liao, X.; Li, Y. Genetic associations between voltage-gated calcium channels and Autism spectrum disorder: A systematic review. Mol. Brain 2020, 13, 1–10. [Google Scholar] [CrossRef]
- Chien, Y.L.; Wu, Y.Y.; Chiu, Y.N.; Liu, S.K.; Tsai, W.C.; Lin, P.I.; Chen, C.H.; Gau, S.S.F.; Chien, W.H. Association study of the CNS patterning genes and Autism in Han Chinese in Taiwan. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2011, 35, 1512–1517. [Google Scholar] [CrossRef]
- Yoo, H.J.; Cho, I.H.; Park, M.; Yang, S.Y.; Kim, S.A. Family based association of GRIN2A and GRIN2B with Korean Autism spectrum disorders. Neurosci. Lett. 2012, 512, 89–93. [Google Scholar] [CrossRef]
- Lim, E.T.; Uddin, M.; De Rubeis, S.; Chan, Y.; Kamumbu, A.S.; Zhang, X.; D’Gama, A.M.; Kim, S.N.; Hill, R.S.; Goldberg, A.P.; et al. Rates, Distribution and implications of postzygotic mosaic mutations in Autism spectrum disorder. Nat. Neurosci. 2017, 20, 1217–1224. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Das, S.; Guhathakurta, S.; Sen, B.; Sinha, S.; Chatterjee, A.; Ghosh, S.; Ahmed, S.; Ghosh, S.; Usha, R. Glutamate receptor 6 gene (GluR6 or GRIK2) polymorphisms in the Indian population: A genetic association study on Autism spectrum disorder. Cell. Mol. Neurobiol. 2007, 27, 1035–1047. [Google Scholar] [CrossRef]
- Akhter, M.; Ashraf, M.; Ali, A.; Rizwan, I.; Rehman, R. Integration of therapies in Autistic children; A survey based in Karachi, Pakistan. J. Pak. Med. Assoc. 2018, 68, 1508. [Google Scholar]
- Furrukh, J.; Anjum, G. Coping with Autism spectrum disorder (ASD) in Pakistan: A phenomenology of mothers who have children with ASD. Cogent Psychol. 2020, 7, 1728108. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001; p. 999. [Google Scholar] [CrossRef]
- Clarke, G.M.; Anderson, C.A.; Pettersson, F.H.; Cardon, L.R.; Morris, A.P.; Zondervan, K.T. Basic statistical analysis in genetic case-control studies. Nat. Protoc. 2011, 6, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.A.; Pettersson, F.H.; Clarke, G.M.; Cardon, L.R.; Morris, A.P.; Zondervan, K.T. Data quality control in genetic case-control association studies. Nat. Protoc. 2010, 5, 1564–1573. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING V10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Amorim, I.S.; Lach, G.; Gkogkas, C.G. The role of the eukaryotic translation initiation factor 4E (EIF4E) in neuropsychiatric disorders. Front. Genet. 2018, 9, 561. [Google Scholar] [CrossRef] [Green Version]
- Yonan, A.L.; Alarcón, M.; Cheng, R.; Magnusson, P.K.E.; Spence, S.J.; Palmer, A.A.; Grunn, A.; Hank Juo, S.-H.; Terwilliger, J.D.; Liu, J.; et al. A Genome-wide screen of 345 families for Autism-susceptibility loci. Am. J. Hum. Genet. 2003, 73, 886–897. [Google Scholar] [CrossRef] [Green Version]
- Schellenberg, G.D.; Dawson, G.; Sung, Y.J.; Estes, A.; Munson, J.; Rosenthal, E.; Rothstein, J.; Flodman, P.; Smith, M.; Coon, H.; et al. Evidence for multiple loci from a genome scan of Autism kindreds. Mol. Psychiatry 2006, 11, 1049–1060. [Google Scholar] [CrossRef] [Green Version]
- So, H.C.; Chau, C.K.L.; Chiu, W.T.; Ho, K.S.; Lo, C.P.; Yim, S.H.Y.; Sham, P.C. Analysis of genome-wide association data highlights candidates for drug repositioning in psychiatry. Nat. Neurosci. 2017, 20, 1342. [Google Scholar] [CrossRef]
- Inoue, Y.U.; Inoue, T. Brain enhancer activities at the gene-poor 5p14.1 Autism-associated locus. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Salyakina, D.; Jaworski, J.M.; Konidari, I.; Whitehead, P.L.; Andersen, A.N.; Hoffman, J.D.; Slifer, S.H.; Hedges, D.J.; Cukier, H.N.; et al. A genome-wide association study of Autism reveals a common novel risk locus at 5p14.1. Ann. Hum. Genet. 2009, 73, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Bigos, K.L.; Mattay, V.S.; Callicott, J.H.; Straub, R.E.; Vakkalanka, R.; Kolachana, B.; Hyde, T.M.; Lipska, B.K.; Kleinman, J.E.; Weinberger, D.R. Genetic variation in CACNA1C affects brain circuitries related to mental illness. Arch. Gen. Psychiatry 2010, 67, 939–945. [Google Scholar] [CrossRef] [Green Version]
- Ivorra, J.L.; Rivero, O.; Costas, J.; Iniesta, R.; Arrojo, M.; Ramos-Ríos, R.; Carracedo, Á.; Palomo, T.; Rodriguez-Jimenez, R.; Cervilla, J.; et al. Replication of previous genome-wide association studies of psychiatric diseases in a large Schizophrenia case-control sample from Spain. Schizophr. Res. 2014, 159, 107–113. [Google Scholar] [CrossRef]
- Nyegaard, M.; Demontis, D.; Foldager, L.; Hedemand, A.; Flint, T.J.; Sørensen, K.M.; Andersen, P.S.; Nordentoft, M.; Werge, T.; Pedersen, C.B.; et al. CACNA1C (Rs1006737) is associated with Schizophrenia. Mol. Psychiatry 2010, 15, 119–121. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhao, L.; You, Y.; Lu, T.; Jia, M.; Yu, H.; Ruan, Y.; Yue, W.; Liu, J.; Lu, L.; et al. Schizophrenia related variants in CACNA1C also confer risk of Autism. PLoS ONE 2015, 10, e0133247. [Google Scholar] [CrossRef]
- Scott-Van Zeeland, A.A.; Abrahams, B.S.; Alvarez-Retuerto, A.I.; Sonnenblick, L.I.; Rudie, J.D.; Ghahremani, D.; Mumford, J.A.; Poldrack, R.A.; Dapretto, M.; Geschwind, D.H.; et al. Altered functional connectivity in frontal lobe circuits is associated with variation in the Autism risk gene CNTNAP. Sci. Transl. Med. 2010, 56, 56ra80. [Google Scholar] [CrossRef] [Green Version]
- McNealy, K.; Mazziotta, J.C.; Dapretto, M. Cracking the language code: Neural mechanisms underlying speech parsing. J. Neurosci. 2006, 26, 7629–7639. [Google Scholar] [CrossRef]
- Knowlton, B.J.; Mangels, J.A.; Squire, L.R. A Neostriatal habit learning system in humans. Science 1996, 273, 1399–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, A.L.; Haan, N.; Wilkinson, L.S.; Thomas, K.L.; Hall, J. CACNA1C: Association with psychiatric disorders, behavior, and neurogenesis. Schizophr. Bull. 2018, 44, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Yang, J.; Qiu, X.; Yang, X.; Qiao, Z.; Song, X.; Wang, L.; Zhao, E.; Yang, Y.; Cao, D. CACNA1C Rs1006737, threatening life events, and gene–environment interaction predict major depressive disorder. Front. Psychiatry 2020, 10. [Google Scholar] [CrossRef]
- Roopasree, B.; Joseph, J.; Mukkadan, J.K. Oxytocin-functions: An overview. MOJ Anat. Physiol. 2019, 6, 128–133. [Google Scholar] [CrossRef]
- Nguyen, R.L.; Medvedeva, Y.V.; Ayyagari, T.E.; Schmunk, G.; Gargus, J.J. Intracellular calcium dysregulation in Autism spectrum disorder: An analysis of converging organelle signaling pathways. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 1718–1732. [Google Scholar] [CrossRef]
- Wen, Y.; Alshikho, M.J.; Herbert, M.R. Pathway network analyses for Autism reveal multisystem involvement, major overlaps with other diseases and convergence upon MAPK and calcium signaling. PLoS ONE 2016, 11, e0153329. [Google Scholar] [CrossRef]
- Wen, Y. Involvement of Calcium, Ras, MAPK, PI3K-Akt and MTOR signaling pathways in Autism spectrum disorders. Neurol. Neurother. Open Access J. 2017, 2, 000110. [Google Scholar] [CrossRef]
- Kim, H.; Lim, C.S.; Kaang, B.K. Neuronal mechanisms and circuits underlying repetitive behaviors in mouse models of Autism spectrum disorder. Behav. Brain Funct. 2016, 12, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, J.; Li, J.; O’Hara, B.; Alberts, I.; Xiong, L.; Li, J.; Li, X. The role of GABAergic neural circuits in the pathogenesis of Autism spectrum disorder. Int. J. Dev. Neurosci. 2020, 80, 73–85. [Google Scholar] [CrossRef]
- Cioana, M.; Michalski, B.; Fahnestock, M. Insulin-like growth factor and insulin-like growth factor receptor expression in human idiopathic Autism fusiform gyrus tissue. Autism Res. 2020, 6, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Lo, F.S.; Erzurumlu, R.S. Insulin receptor sensitization restores neocortical excitation/inhibition balance in a mouse model of Autism. Mol. Autism 2018, 13, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmood, H.M.; Aldhalaan, H.M.; Alshammari, T.K.; Alqasem, M.A.; Alshammari, M.A.; Albekairi, N.A.; AlSharari, S.D. the role of nicotinic receptors in the attenuation of autism-related behaviors in a murine BTBR T + Tf/J Autistic model. Autism Res. 2020, 13, 1311–1334. [Google Scholar] [CrossRef]
- Marotta, R.; Risoleo, M.C.; Messina, G.; Parisi, L.; Carotenuto, M.; Vetri, L.; Roccella, M. The neurochemistry of autism. Brain Sci. 2020, 10, 163. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Controls | ASD | p-Value | |
|---|---|---|---|
| Total Samples | 93 | 93 | data |
| Mean Age ± SD | 39.98 ± 1.87 | 8.409 ± 0.5784 | <0.001 a |
| Median | 40 | 7 | |
| Interquartile Range | 26.5 | 5.5 | |
| Total Number (Percentage) Males | 41 (44.09%) | 68 (73.12%) | <0.001 b |
| Total Number (Percentage) Females | 52 (55.91%) | 25 (26.88%) |
| Genotype | Controls (n = 93) | Cases (n = 93) | OR (95% CI) p-Value | Adjusted OR (95% CI) p-Value |
|---|---|---|---|---|
| ATP2B2/rs35678 (C/T) | ||||
| CC | 14 (15.1%) | 3 (3.2%) | Reference | |
| CT | 47 (50.5%) | 48 (51.6%) | *4.76 (1.28–17.66) 0.01 | 0.42 (0.02–6.23) 0.52 |
| TT | 32 (34.4%) | 42 (45.2%) | *6.12 (1.62–23.13) 0.007 | 1.37 (0.09–19.88) 0.81 |
| CNTNAP2/rs7794745 (A/T) | ||||
| AA | 38 (40.9%) | 28 (30.1%) | Reference | |
| AT | 47 (50.5%) | 45 (48.4%) | 1.29 (0.68–2.45) 0.42 | 0.98 (0.26–3.68) 0.977 |
| TT | 8 (8.6%) | 20 (21.5%) | *3.39 (1.30–8.81) 0.01 | 2.98 (0.28–31.16) 0.361 |
| CACNA1C/rs1006737 (G/A) | ||||
| GG | 58 (62.4%) | 52 (55.9%) | Reference | |
| GA | 35 (37.6%) | 41 (44.1%) | 1.30 (0.72–2.34) 0.37 | 1.41 (0.40–4.92) 0.584 |
| (CDH9/CDH10)/rs4307059 (C/T) | ||||
| CC | 12 (12.9%) | 10 (10.8%) | Reference | |
| CT | 47 (50.5) | 45 (48.4) | 1.14 (0.45–2.92) 0.77 | 0.26 (0.01–3.62) 0.319 |
| TT | 34 (36.6) | 38 (40.9) | 1.34 (0.51–3.49) 0.54 | 0.05 (0.002–1.02) 0.05 |
| EIF4E/rs17850950 | ||||
| CC | 93 (100%) | 93 (100%) | -- | -- |
| CT | 0 | 0 | -- | -- |
| TT | 0 | 0 | -- | -- |
| Genes/SNPs IDs | Alleles | Controls (%) | Cases (%) | OR (95%CI) p-Value | Adjusted OR (95%CI) p-Value |
|---|---|---|---|---|---|
| ATP2B2/rs35678 | T | 60 | 71 | 1.04 (0.59–1.86) 0.88 | 0.29 (0.06–1.38) 0.11 |
| C | 40 | 29 | |||
| CNTNAP2/rs7794745 | T | 34 | 46 | *1.68 (1.09–*2.61) 0.017 | 0.86 (0.28–2.65) 0.79 |
| A | 66 | 54 | |||
| CACNA1C/rs1006737 | A | 19 | 22 | 1.21 (0.73–2.02) 0.44 | 1.21 (0.73–2.02) 0.44 |
| G | 81 | 78 | |||
| (CDH9/CDH10)/rs4307059 | T | 62 | 65 | 1.16 (0.75–1.80) 0.5 | 0.72 (0.20–2.58) 0.61 |
| C | 38 | 35 |
| Number of Risk Alleles | Controls (n = 93) | Cases (n = 93) | OR (95% CI) p-Value | Adjusted OR (95% CI) p-Value |
|---|---|---|---|---|
| 1 | 6 (6.5%) | 1 (1.1%) | Reference | |
| 2 | 15 (16.1%) | 8 (8.6%) | 3.19 (0.326–31.391) 0.31 | 2.175 (0.010–463.87) 0.776 |
| 3 | 27 (29.03%) | 25 (26.9%) | 5.55 (0.624–49.38) 0.124 | 3.53 (0.019–650.02) 0.634 |
| 4 | 24 (25.8%) | 22 (23.7%) | 5.49 (0.613–49.32) 0.127 | 3.16 (0.016–611.30) 0.667 |
| 5+ | 21 (22.6%) | 37 (39.7%) | 10.56 (1.19–93.77) 0.034 | 2.114 (0.011–40.7.04) 0.780 |
| Clinical Phenotype | Classification | ATP2B2 rs35678 | CNTNAP2 rs7794745 | CACNA1C rs1006737 | rs4307059 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CC | CT | TT | p-Value | GG | GA | AA | p-Value | GG | GA | p-Value | CC | CT | TT | p-Value | ||
| Licking | Yes | 2 | 29 | 23 | 0.824 | 17 | 26 | 11 | 0.923 | 26 | 28 | 0.076 | 4 | 27 | 23 | 0.471 |
| No | 1 | 19 | 19 | 11 | 19 | 9 | 26 | 23 | 6 | 18 | 15 | |||||
| Hand functioning | Ok | 2 | 45 | 40 | 0.150 | 25 | 43 | 19 | 0.545 | 47 | 40 | 0.162 | 10 | 42 | 35 | 0.662 |
| Poor | 1 | 3 | 2 | 3 | 2 | 1 | 5 | 1 | 0 | 3 | 3 | |||||
| Self-care skills | Fully dependent | 2 | 29 | 25 | 0.983 | 19 | 30 | 7 | 0.031 | 32 | 24 | 0.721 | 8 | 28 | 20 | 0.527 |
| Needs help | 1 | 17 | 16 | 7 | 14 | 13 | 19 | 15 | 2 | 15 | 17 | |||||
| Good | 0 | 2 | 1 | 2 | 1 | 0 | 1 | 2 | 0 | 2 | 1 | |||||
| Vision problems | Yes | 0 | 4 | 1 | 0.420 | 0 | 3 | 2 | 0.275 | 1 | 4 | 0.096 | 1 | 1 | 3 | 0.412 |
| No | 3 | 44 | 41 | 28 | 42 | 18 | 51 | 37 | 9 | 44 | 35 | |||||
| Smells everything | Yes | 0 | 15 | 22 | 0.045 | 12 | 21 | 4 | 0.118 | 18 | 19 | 0.251 | 3 | 20 | 14 | 0.623 |
| No | 3 | 33 | 20 | 16 | 24 | 16 | 34 | 22 | 7 | 25 | 24 | |||||
| Looks closely from eye corner | Yes | 1 | 34 | 29 | 0.396 | 18 | 34 | 12 | 0.378 | 33 | 31 | 0.209 | 6 | 33 | 25 | 0.621 |
| No | 2 | 14 | 13 | 10 | 11 | 8 | 19 | 10 | 4 | 12 | 13 | |||||
| Cover ear in noise | Yes | 0 | 28 | 21 | 0.130 | 12 | 25 | 12 | 0.435 | 26 | 23 | 0.559 | 5 | 28 | 16 | 0.185 |
| No | 3 | 20 | 21 | 16 | 20 | 8 | 26 | 18 | 5 | 17 | 22 | |||||
| Rocking and swaying | Yes | 3 | 40 | 33 | 0.596 | 25 | 36 | 15 | 0.413 | 46 | 30 | 0.058 | 7 | 41 | 28 | 0.074 |
| No | 0 | 8 | 9 | 3 | 9 | 5 | 6 | 11 | 3 | 4 | 10 | |||||
| Repetitive behaviors | Yes | 3 | 44 | 39 | 0.862 | 26 | 41 | 19 | 0.857 | 46 | 40 | 0.099 | 10 | 43 | 33 | 0.206 |
| No | 0 | 4 | 3 | 2 | 4 | 1 | 6 | 1 | 0 | 2 | 5 | |||||
| Likes circular moving objects | Yes | 1 | 21 | 24 | 0.381 | 14 | 24 | 8 | 0.610 | 22 | 24 | 0.120 | 6 | 23 | 17 | 0.659 |
| No | 2 | 27 | 18 | 14 | 21 | 12 | 30 | 17 | 4 | 22 | 21 | |||||
| Reciprocates smiles | Yes | 2 | 14 | 12 | 0.554 | 4 | 15 | 9 | 0.209 | 15 | 13 | 0.760 | 3 | 15 | 10 | 0.748 |
| Rarely | 0 | 18 | 13 | 11 | 14 | 6 | 19 | 12 | 4 | 16 | 11 | |||||
| No | 1 | 16 | 17 | 13 | 16 | 5 | 18 | 16 | 3 | 14 | 17 | |||||
| Eye contact | Good | 2 | 15 | 16 | 0.744 | 13 | 15 | 5 | 0.485 | 20 | 13 | 0.616 | 5 | 13 | 15 | 0.666 |
| Poor | 1 | 31 | 25 | 15 | 28 | 14 | 31 | 26 | 5 | 30 | 22 | |||||
| No | 0 | 2 | 1 | 0 | 2 | 1 | 1 | 3 | 0 | 2 | 1 | |||||
| Unusual noise | Yes | 1 | 27 | 20 | 0.582 | 14 | 24 | 10 | 0.950 | 27 | 21 | 0.946 | 2 | 22 | 24 | 0.046 |
| No | 2 | 21 | 22 | 14 | 21 | 10 | 25 | 20 | 8 | 23 | 14 | |||||
| Echolalia | Yes | 0 | 18 | 15 | 0.420 | 7 | 18 | 8 | 0.382 | 18 | 15 | 0.844 | 1 | 17 | 15 | 0.201 |
| No | 3 | 30 | 27 | 21 | 27 | 12 | 34 | 26 | 9 | 28 | 23 | |||||
| Sense of being praised | Yes | 1 | 13 | 7 | 0.450 | 7 | 10 | 4 | 0.917 | 9 | 12 | 0.171 | 0 | 10 | 11 | 0.149 |
| No | 2 | 35 | 35 | 21 | 35 | 16 | 43 | 29 | 10 | 35 | 27 | |||||
| Aggressive | Yes | 0 | 18 | 21 | 0.159 | 14 | 12 | 13 | 0.009 | 16 | 23 | 0.014 | 4 | 17 | 18 | 0.672 |
| No | 3 | 30 | 21 | 14 | 33 | 7 | 36 | 18 | 6 | 28 | 20 | |||||
| Speech | No Speech | 2 | 21 | 12 | 0.249 | 13 | 16 | 6 | 0.592 | 21 | 14 | 0.920 | 4 | 18 | 13 | 0.969 |
| Able to make short sentences | 0 | 13 | 15 | 7 | 14 | 7 | 15 | 13 | 4 | 12 | 12 | |||||
| Can string few words | 0 | 8 | 8 | 4 | 8 | 4 | 9 | 7 | 1 | 9 | 6 | |||||
| Only a few single words | 1 | 3 | 7 | 4 | 6 | 1 | 6 | 5 | 1 | 5 | 5 | |||||
| Normal | 0 | 3 | 0 | 0 | 1 | 2 | 1 | 2 | 0 | 1 | 2 | |||||
| Idiosyncratic language | Yes | 0 | 6 | 8 | 0.522 | 2 | 8 | 4 | 0.365 | 7 | 7 | 0.629 | 2 | 7 | 5 | 0.858 |
| No | 3 | 42 | 34 | 26 | 37 | 16 | 45 | 34 | 8 | 38 | 33 | |||||
| Understanding (Cognitive) | Good | 1 | 4 | 0 | 0.020 | 1 | 3 | 1 | 0.664 | 3 | 2 | 0.349 | 0 | 3 | 2 | 0.034 |
| Limited | 0 | 11 | 18 | 6 | 16 | 7 | 13 | 16 | 5 | 7 | 17 | |||||
| Poor | 2 | 33 | 24 | 21 | 26 | 12 | 36 | 23 | 5 | 35 | 19 | |||||
| Shared enjoyments with parents | Yes | 0 | 5 | 3 | 0.742 | 2 | 2 | 4 | 0.112 | 3 | 5 | 0.273 | 0 | 3 | 5 | 0.340 |
| No | 3 | 43 | 39 | 26 | 43 | 16 | 49 | 36 | 10 | 42 | 33 | |||||
| Follow instructions | Yes | 0 | 6 | 5 | 0.976 | 2 | 4 | 5 | 0.07 | 8 | 3 | 0.08 | 4 | 12 | 14 | 0.810 |
| Very simple | 1 | 15 | 14 | 7 | 14 | 9 | 12 | 18 | 1 | 5 | 5 | |||||
| No | 2 | 27 | 23 | 19 | 27 | 6 | 32 | 20 | 5 | 28 | 19 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalid, M.; Raza, H.; M. Driessen, T.; J. Lee, P.; Tejwani, L.; Sami, A.; Nawaz, M.; Mehmood Baig, S.; Lim, J.; Kaukab Raja, G. Genetic Risk of Autism Spectrum Disorder in a Pakistani Population. Genes 2020, 11, 1206. https://doi.org/10.3390/genes11101206
Khalid M, Raza H, M. Driessen T, J. Lee P, Tejwani L, Sami A, Nawaz M, Mehmood Baig S, Lim J, Kaukab Raja G. Genetic Risk of Autism Spectrum Disorder in a Pakistani Population. Genes. 2020; 11(10):1206. https://doi.org/10.3390/genes11101206
Chicago/Turabian StyleKhalid, Madiha, Hashim Raza, Terri M. Driessen, Paul J. Lee, Leon Tejwani, Abdul Sami, Muhammad Nawaz, Shahid Mehmood Baig, Janghoo Lim, and Ghazala Kaukab Raja. 2020. "Genetic Risk of Autism Spectrum Disorder in a Pakistani Population" Genes 11, no. 10: 1206. https://doi.org/10.3390/genes11101206
APA StyleKhalid, M., Raza, H., M. Driessen, T., J. Lee, P., Tejwani, L., Sami, A., Nawaz, M., Mehmood Baig, S., Lim, J., & Kaukab Raja, G. (2020). Genetic Risk of Autism Spectrum Disorder in a Pakistani Population. Genes, 11(10), 1206. https://doi.org/10.3390/genes11101206