Expression of Piwi Genes during the Regeneration of Lineus sanguineus (Nemertea, Pilidiophora, Heteronemertea)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Gene Identification Analysis
2.3. Riboprobe Synthesis
2.4. Regeneration and In Situ Hybridization
2.5. Histology
3. Results
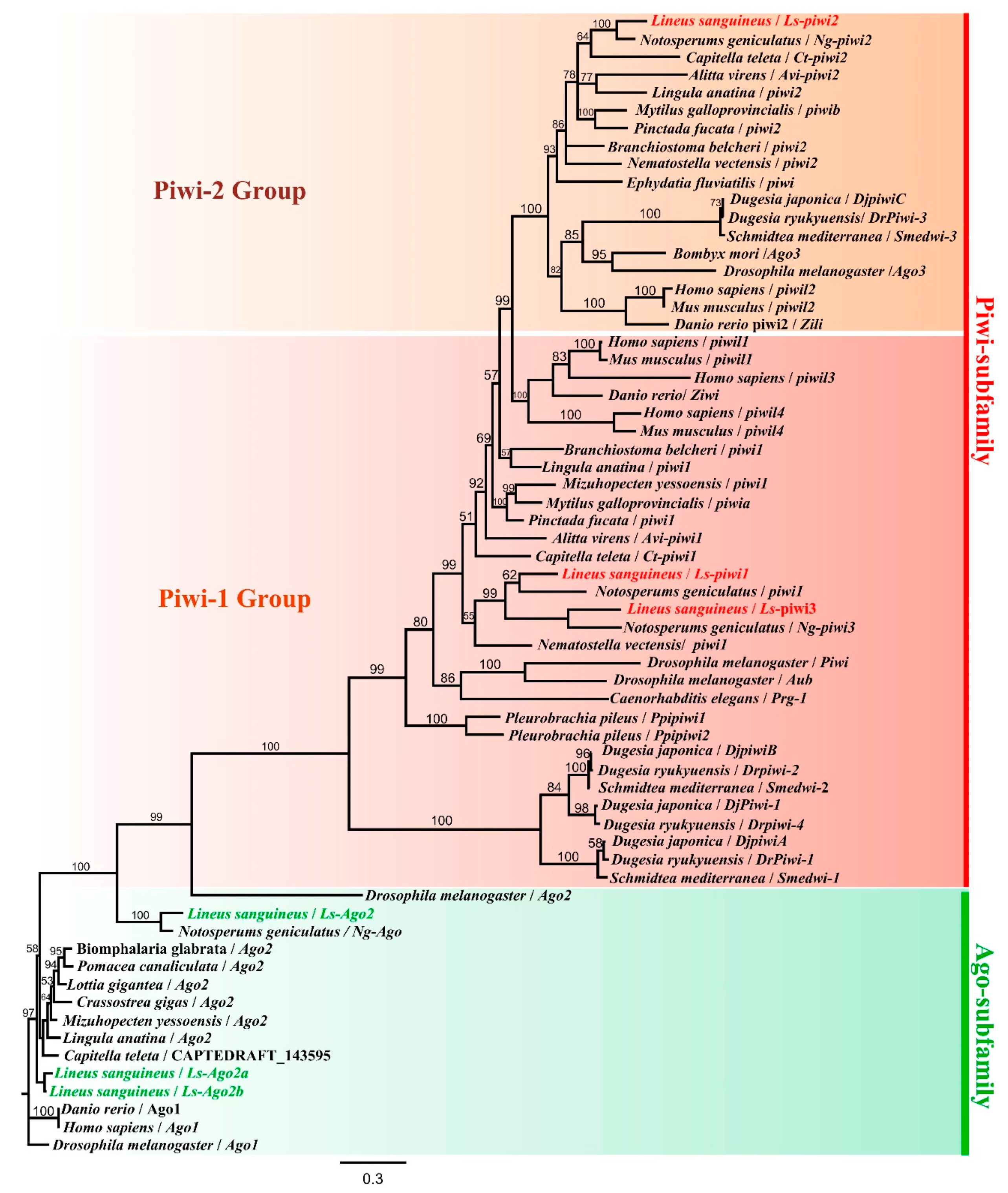
3.1. Characterization of Argonaute Family in Lineus sanguineus and Notospermus geniculatus
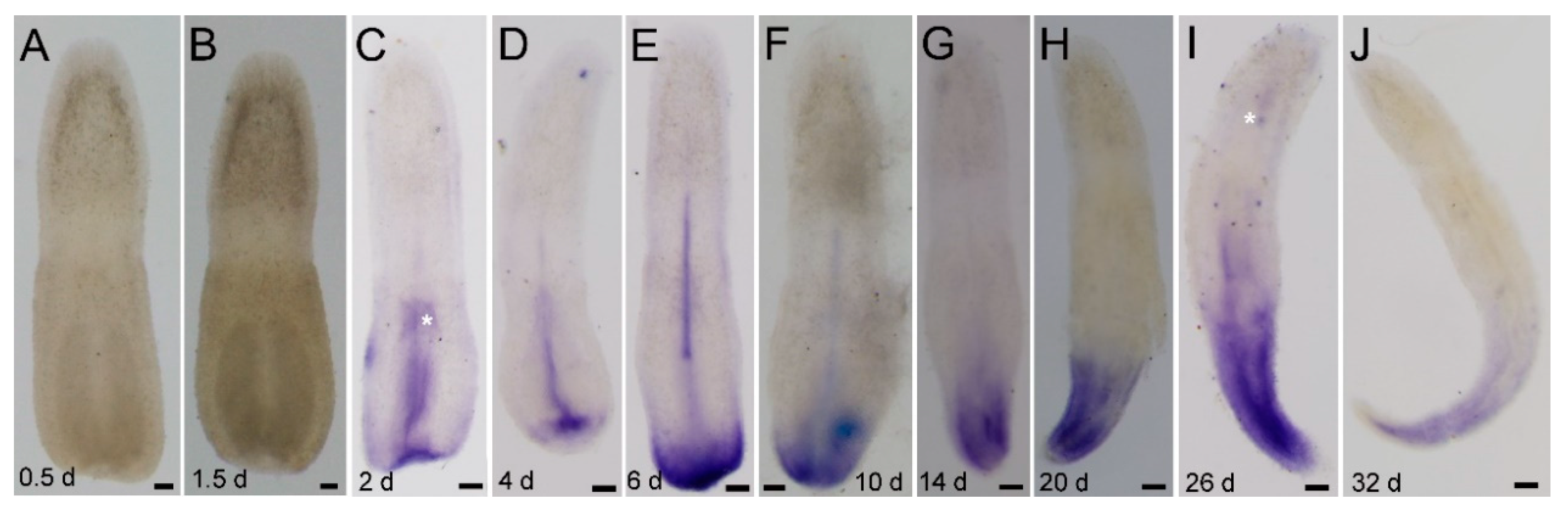
3.2. Regenerative Time Course of Lineus sanguineus
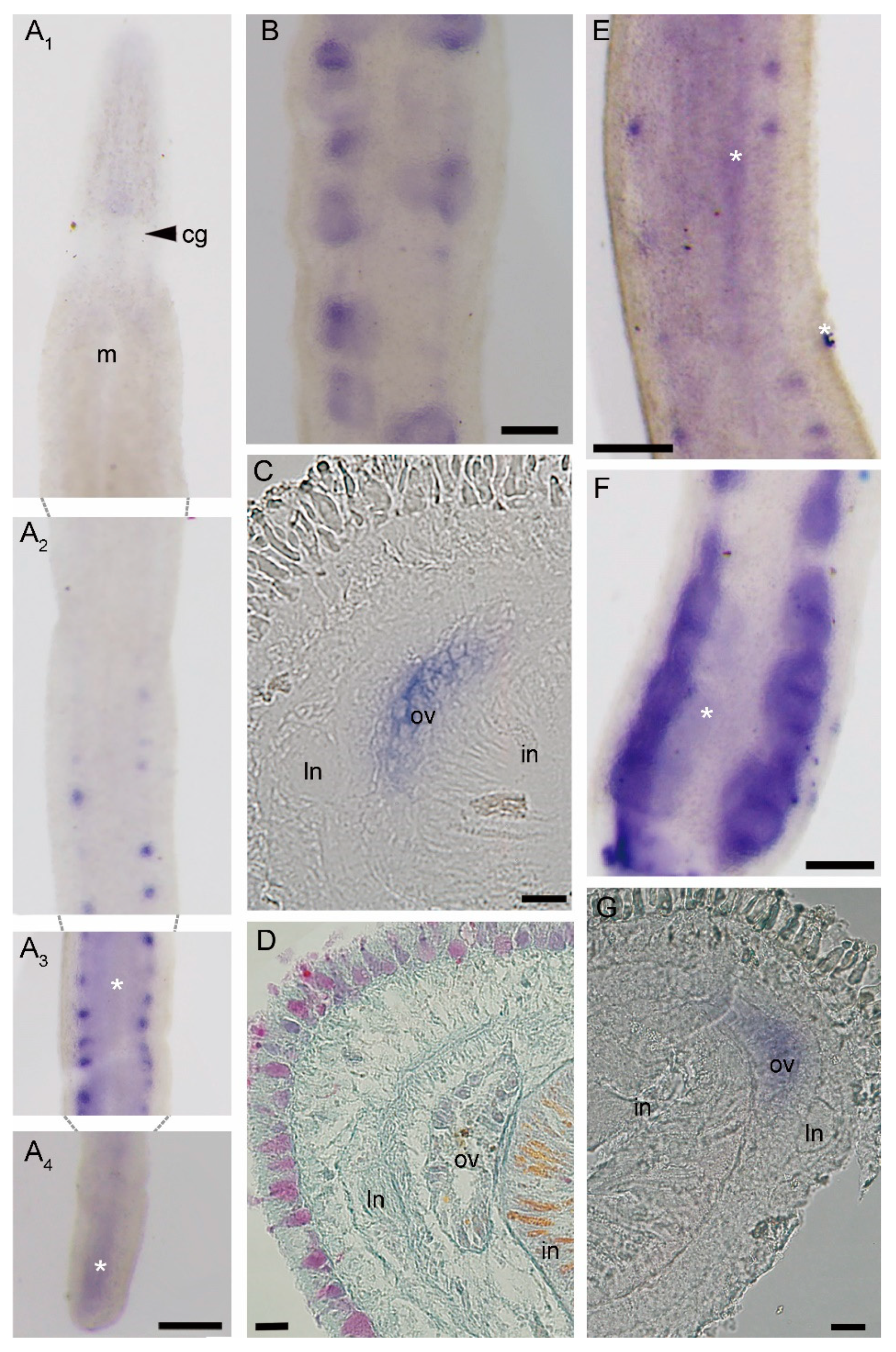
3.3. Expression Patterns of piwi and pcna Genes in Intact Worms
3.4. Expression of piwi and pcna Genes during Regeneration
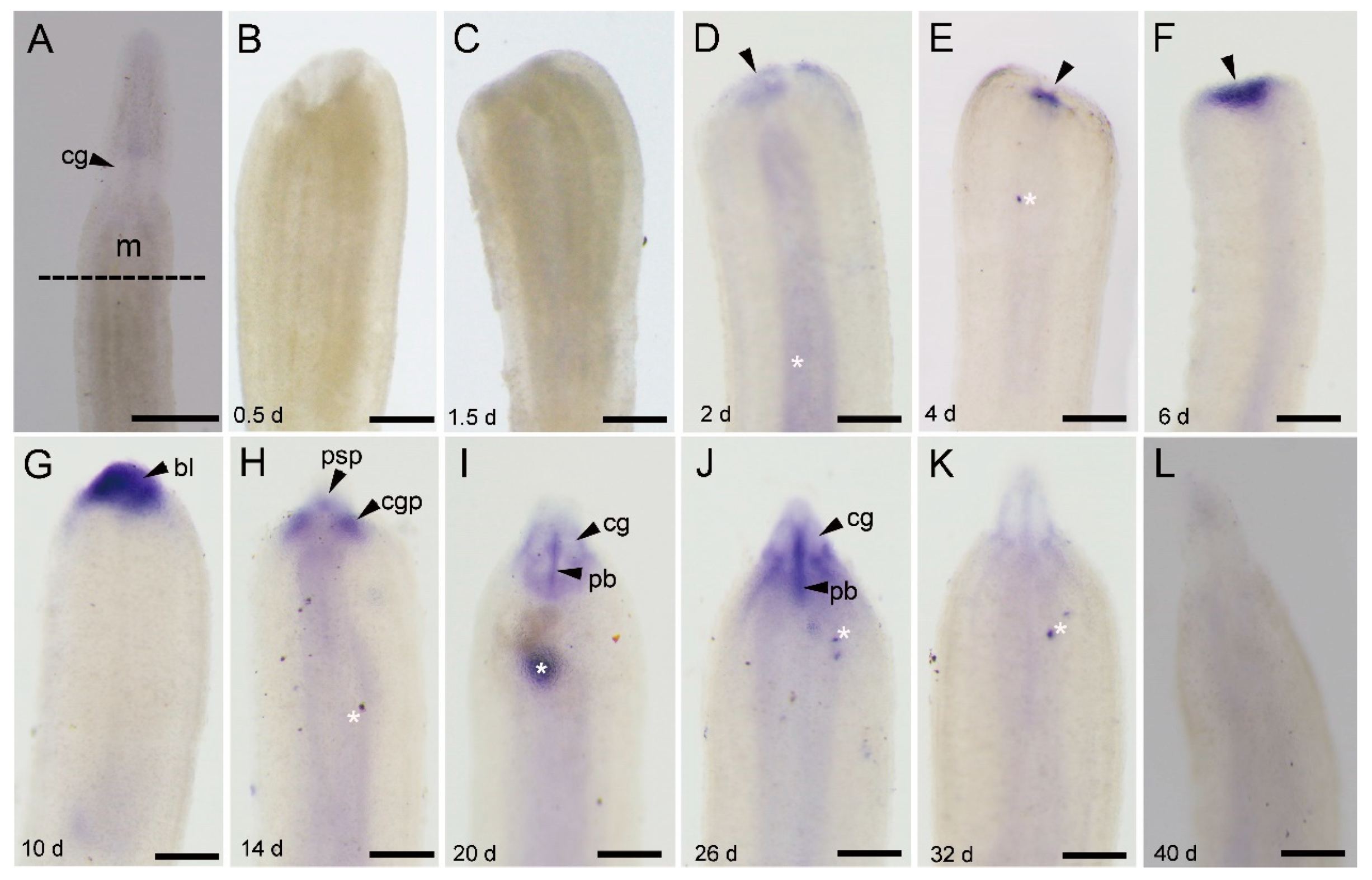
3.4.1. Ls-piwi1 and Ls-pcna Expression during Anterior Regeneration
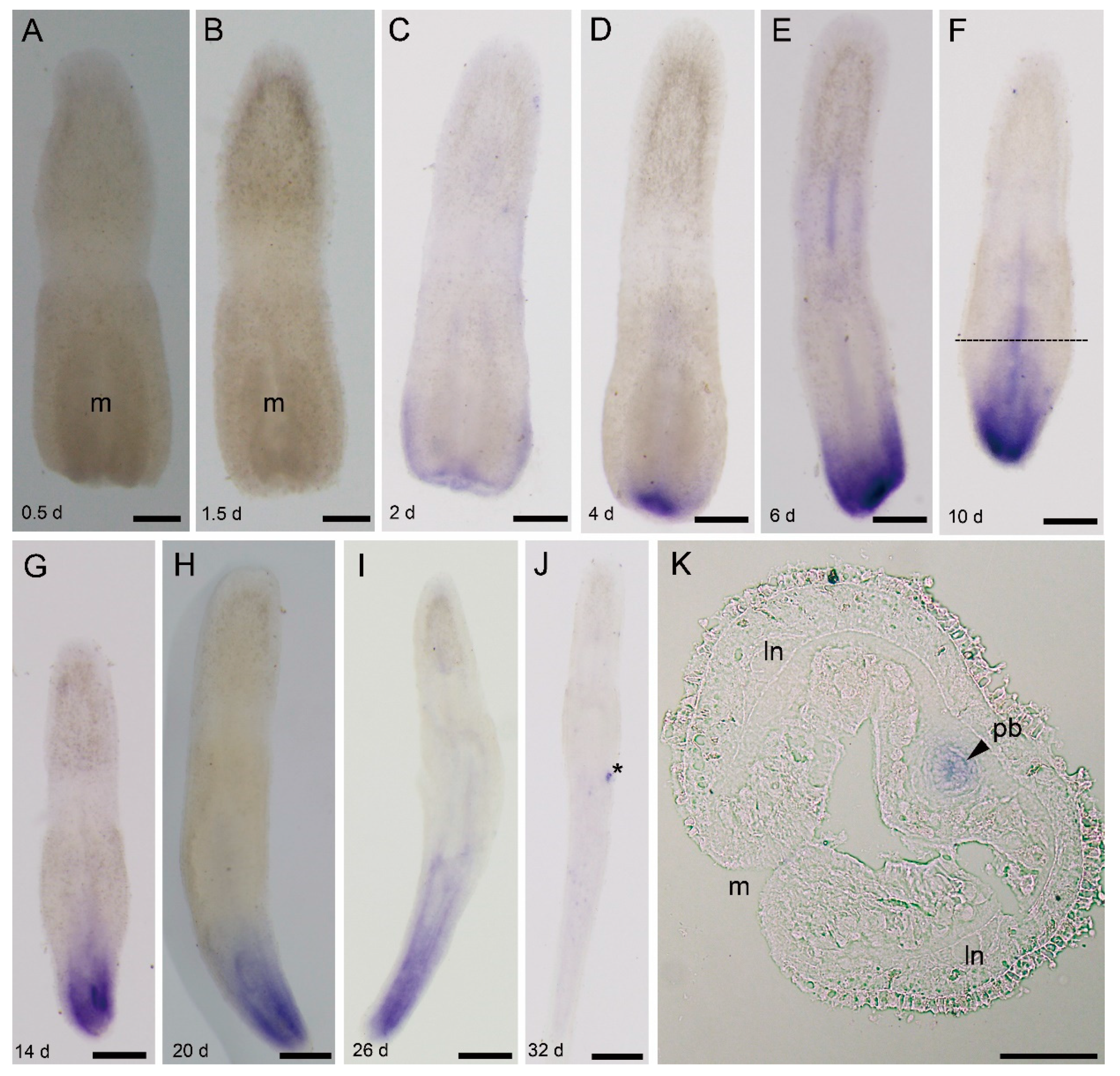
3.4.2. Ls-piwi1 and Ls-pcna Expression during Posterior Regeneration
3.5. Ls-piwi2 and Ls-piwi3 Expression during Regeneration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coe, W.R. Regeneration in nemerteans. II. Regeneration of small sections of the body split or partially split longitudinally. J. Exp. Zool. 1930, 57, 109–144. [Google Scholar] [CrossRef]
- Coe, W.R. Regeneration in nemerteans. J. Exp. Zool. 1929, 54, 411–459. [Google Scholar] [CrossRef]
- Zhuravleva, N.G.; Korotkevich, V.S.; Korotkova, G.P. Vosstanovitel’ nye morfogenezy u nemertin. Arkh. Anat. Gistol. Embriol. 1970, 59, 12–22. [Google Scholar] [PubMed]
- Coe, W.R. Regeneration in nemerteans. IV. Cellular changes involved in restitution and reorganization. J. Exp. Zool. 1934, 67, 283–314. [Google Scholar] [CrossRef]
- Coe, W.R. Analysis of the regenerative processes in nemerteans. Biol. Bull. 1934, 66, 304–315. [Google Scholar] [CrossRef]
- Baguna, J.; Saló, E.; Auladell, C. Regeneration and pattern formation in planarians. III. Evidence that neoblasts are totipotent stem cells and the cells. Development 1989, 107, 77–86. [Google Scholar]
- Turbeville, J.M. Nemertinea. In Microscopic Anatomy of Invertebrates; Harrison, F.W., Bogitsh, B.J., Eds.; Wiley-Liss Press: New York, NY, USA, 1991; pp. 285–328. [Google Scholar]
- Cox, D.N.; Chao, A.; Baker, J.; Chang, L.; Qiao, D.; Lin, H. A novel class of evolutionarily conserved genes defined by piwi are essential for stem cell self-renewal. Genes Dev. 1998, 12, 3715–3727. [Google Scholar] [CrossRef] [Green Version]
- Hutvagner, G.; Simard, M.J. Argonaute proteins: Key players in RNA silencing. Nat. Rev. Mol. Cell Biol. 2008, 9, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Höck, J.; Meister, G. The Argonaute protein family. Genome Biol. 2008, 9, 210. [Google Scholar] [CrossRef]
- Juliano, C.; Wang, J.; Lin, H. Uniting germline and stem cells: The function of Piwi proteins and the piRNA pathway in diverse organisms. Annu. Rev. Genet. 2011, 45, 447–469. [Google Scholar] [CrossRef] [Green Version]
- Thomson, T.; Lin, H. The Biogenesis and Function PIWI Proteins and piRNAs: Progress and Prospect. Annu. Rev. Cell Dev. Biol. 2009, 25, 355–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Spradling, A.C. A novel group of pumilio mutations affects the asymmetric division of germline stem cells in the Drosophila ovary. Development 1997, 124, 2463–2476. [Google Scholar] [PubMed]
- Houwing, S.; Kamminga, L.M.; Berezikov, E.; Cronembold, D.; Girard, A.; van den Elst, H.; Filippov, D.V.; Blaser, H.; Raz, E.; Moens, C.B.; et al. A role for Piwi and piRNAs in germ cell maintenance and transposon silencing in Zebrafish. Cell 2007, 129, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Juliano, C.E.; Voronina, E.; Stack, C.; Aldrich, M.; Cameron, A.R.; Wessel, G.M. Germline determinants are not localized early in sea urchin development, but do accumulate in the small micromere lineage. Dev. Biol. 2006, 300, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Alié, A.; Leclère, L.; Jager, H.; Dayraud, C.; Chang, P.; Guyader, L.H.; Quéinnec, E.; Manuel, M. Somatic stem cells express Piwi and Vasa genes in an adult ctenophore: Ancient association of “germline genes” with stemness. Dev. Biol. 2011, 350, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Seipel, K.; Yanze, N.; Schmid, V. The germ line and somatic stem cell gene Cniwi in the jellyfish Podocoryne carnea. Int. J. Dev. Biol. 2004, 48, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Reddien, P.W.; Oviedo, N.J.; Jennings, J.R.; Jenkin, J.C.; Alvarado, A.S. SMEDWI-2 is a PIWI-like protein that regulates planarian stem cells. Science 2005, 310, 1327–1330. [Google Scholar] [CrossRef] [Green Version]
- Palakodeti, D.; Smielewska, M.; Lu, Y.C.; Yeo, G.W.; Graveley, B.R. The PIWI proteins SMEDWI-2 and SMEDWI-3 are required for stem cell function and piRNA expression in planarians. RNA 2008, 14, 1174–1186. [Google Scholar] [CrossRef] [Green Version]
- Bravo, R.; Frank, R.; Blundell, P.A.; MacDonald-Bravo, H. Cyclin/PCNA is the auxiliary protein of DNA polymerase -δ. Nature 1987, 326, 515–517. [Google Scholar] [CrossRef]
- Kang, X.-X.; Fernández-Álvarez, F.Á.; Alfaya, J.E.F.; Machordom, A.; Strand, M.; Sundberg, P.; Sun, S.-C. Species diversity of Ramphogordius sanguineus/Lineus ruber like nemerteans (Nemertea: Heteronemertea) and geographic distribution of R. sanguineus. Zool. Sci. 2015, 32, 579–589. [Google Scholar] [CrossRef]
- Ament-Velásquez, S.L.; Figuet, E.; Ballenghien, M.; Zattara, E.E.; Norenburg, J.L.; Fernández-Álvarez, F.A.; Bierne, J.; Bierne, N.; Galtier, N. Population genomics of sexual and asexual lineages in fissiparous ribbon worms (Lineus, Nemertea): Hybridization, polyploidy and the Meselson effect. Mol. Ecol. 2016, 25, 3356–3369. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. Bayesian analysis of molecular evolution using MrBayes. In Statistical Methods in Molecular Evolution; Nielsen, R., Ed.; Springer Press: New York, NY, USA, 2005; pp. 183–226. [Google Scholar]
- FigTree v1.4.4. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 25 November 2018).
- Charpignon, V. Homeobox-Containing Genes in the Nemertean Lineus: Key Players in the Antero-Posterior Body Patterning and in the Specification of the Visual Structures. Ph.D. Thesis, University of Basel, Reims, France, 2007. [Google Scholar]
- Pearson, B.J.; Eisenhoffer, G.T.; Gurley, K.A.; Rink, J.C.; Miller, D.E.; Alvarado, A.S. Formaldehyde-based whole-mount in situ hybridization method for planarians. Dev. Dyn. 2009, 238, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Özpolat, B.D.; Sloane, E.S.; Zattara, E.E.; Bely, A.E. Plasticity and regeneration of gonads in the annelid Pristina leidyi. EvoDevo 2016, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Swarts, D.C.; Makarova, K.; Wang, Y.; Nakanishi, K.; Ketting, R.F.; Koonin, E.V.; Patel, D.J.; Van Der Oost, J. The evolutionary journey of Argonaute proteins. Nat. Struct 2014, 21, 741–753. [Google Scholar] [CrossRef]
- Carmell, M.A.; Xuan, Z.; Zhang, M.Q.; Hannon, G.J. The Argonaute family: Tentacles that reach into RNAi, developmental control, stem cell maintenance, and tumorigenesis. Genes Dev. 2002, 16, 2733–2742. [Google Scholar] [CrossRef] [Green Version]
- Gibson, R. Nemerteans; Hutchinson Press: London, UK, 1972; pp. 28–144. [Google Scholar]
- Cerutti, H.; Casas-Mollano, J.A. On the origin and functions of RNA-mediated silencing: From protists to man. Curr. Genet. 2006, 50, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Cerutti, L.; Mian, N.; Bateman, A. Domains in gene silencing and cell differentiation proteins: The novel PAZ domain and redefinition of the Piwi domain. Trends Biochem. Sci. 2000, 25, 481–482. [Google Scholar] [CrossRef]
- Sigova, A.; Rhind, N.; Zamore, P.D. A single Argonaute protein mediates both transcriptional and posttranscriptional silencing in Schizosaccharomyces pombe. Genes Dev. 2004, 18, 2359–2367. [Google Scholar] [CrossRef] [PubMed]
- Yigit, E.; Batista, P.J.; Bei, Y.; Pang, K.M.; Chen, C.G. Analysis of the C. elegans Argonaute family reveals that distinct Argonautes act sequentially during RNAi. Cell 2006, 127, 747–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, C.; Sun, H.; Poethig, R.S. The Arabidopsis heterochronic gene ZIPPY is an ARGONAUTE family member. Curr. Biol. 2003, 13, 1734–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, W.; Lin, H. Miwi, a murine homolog of piwi, encodes a cytoplasmic protein essential for spermatogenesis. Dev. Cell 2002, 2, 819–830. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Reinke, V.A. A C. elegans Piwi, PRG-1, regulates 21U-RNAs during spermatogenesis. Curr. Biol. 2008, 18, 861–867. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, H.; Ishizu, H.; Hasegawa, R.; Kobayashi, K.; Matsumoto, M. Drpiwi-1 is essential for germline cell formation during sexualization of the planarian Dugesia ryukyuensis. Dev. Biol. 2012, 361, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Cox, D.N.; Chao, A.; Lin, H. Piwi encodes a nucleoplasmic factor whose activity modulates the number and division rate of germline stem cells. Development 2000, 127, 503–514. [Google Scholar]
- Harris, A.N.; Macdonald, P.M. Aubergine encodes a Drosophila polar granule component required for pole cell formation and related to eIF2C. Development 2001, 128, 2823–2832. [Google Scholar]
- Li, C.; Vagin, V.V.; Lee, S.; Xu, J.; Ma, S.; Xi, H.; Seitz, H.; Horwich, M.D.; Syrzycka, M.; Barry, M.; et al. Collapse of germline piRNAs in the absence of Argonaute3 reveals somatic piRNAs in flies. Cell 2009, 137, 509–521. [Google Scholar] [CrossRef] [Green Version]
- Houwing, S.; Berezikov, E.; Ketting, R.F. Zili is required for germ cell differentiation and meiosis in zebrafish. EMBO J. 2008, 27, 2702–2711. [Google Scholar] [CrossRef] [Green Version]
- Kuramochi-Miyagawa, S.; Kimura, T.; Yomogida, K.; Kuroiwa, A.; Tadokoro, Y.; Fujitaa, Y.; Satoa, M.; Matsudac, Y.; Nakanoa, T. Two mouse piwi-related genes: Miwi and mili. Mech. Dev. 2001, 108, 121–133. [Google Scholar] [CrossRef]
- Carmell, M.A.; Girard, A.; van de Kant, H.J.G.; Bourc’his, D.; Bestor, T.H.; Dirk, G.; de Rooij, D.G.; Hannon, G.J. MIWI2 is essential for spermatogenesis and repression of transposons in the mouse male germline. Dev. Cell 2007, 12, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Giani, V.C.; Yamaguchi, E.; Boyle, M.J.; Seaver, E.C. Somatic and germline expression of piwi during development and regeneration in the marine polychaete annelid Capitella teleta. Evodevo 2011, 2, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, R.S.; Anand, A.; Nishimiya-Fujisawa, C.; Kobayashi, S.; Kai, T. Analysis of Hydra PIWI proteins and piRNAs uncover early evolutionary origins of the piRNA pathway. Dev. Biol. 2013, 386, 237–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponz-Segrelles, G.; Bleidorn, C.; Aguado, M.T. Expression of vasa, piwi, and nanos during gametogenesis in Typosyllis antoni (Annelida, Syllidae). Evol. Dev. 2018, 20, 132–145. [Google Scholar] [CrossRef]
- Bely, A.E.; Nyberg, K.G. Evolution of animal regeneration: Re-emergence of a field. Trends Eco. Evol. 2010, 25, 161–170. [Google Scholar] [CrossRef]
- Wolfswinkel, V.; Josien, C. Piwi and potency: PIWI proteins in animal stem cells and regeneration. Integr. Comp. Biol. 2014, 54, 700–713. [Google Scholar] [CrossRef] [Green Version]
- Funayama, N.; Nakatsukasa, M.; Mohri, K.; Masuda, Y.; Agata, K. Piwi expression in archeocytes and choanocytes in demosponges: Insights into the stem cell system in demosponges. Evol. Dev. 2010, 12, 275–287. [Google Scholar] [CrossRef]
- David, C.N. Interstitial stem cells in Hydra: Multipotency and decision-making. Int. J. Dev. Biol. 2012, 56, 489–497. [Google Scholar] [CrossRef]
- Morita, M.; Best, J.B. Electron microscopic studies of planarian regeneration. IV. Cell division of neoblasts in Dugesia dorotocephala. J. Exp. Zool. 1984, 229, 425–436. [Google Scholar] [CrossRef]
- Planques, A.; Malem, J.; Parapar, J.; Vervoort, M.; Gazave, E. Morphological, cellular and molecular characterization of posterior regeneration in the marine annelid Platynereis dumerilii. Dev. Biol. 2018, 445, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Pao, G.M.; Satoh, A.; Cummings, G.; Monaghan, G.R.; Harkins, T.T.; Bryant, S.V.; Voss, S.R.; Gardiner, D.M.; Hunter, T. Activation of germline-specific genes is required for limb regeneration in the Mexican axolotl. Dev. Biol. 2013, 370, 42–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, N.; Kashima, M.; Ishiko, T.; Nishimura, O.; Rouhana, L.; Misaki, K.; Yonemura, S.; Saito, K.; Siomi, H.; Siomi, M.C.; et al. Inheritance of a nuclear PIWI from pluripotent stem cells by somatic descendants ensures differentiation by silencing transposons in planarian. Dev. Cell 2016, 37, 226–237. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, M.; Mazza-Curll, K.L.; van Wolfswinkel, J.C.; Reddien, P.W. Whole-body acoel regeneration is controlled by Wnt and Bmp-Admp signaling. Curr. Biol. 2014, 24, 1107–1113. [Google Scholar] [CrossRef] [Green Version]
- Rinkevich, Y.; Rosner, A.; Rabinowitz, C.; Lapidot, Z.; Moiseeva, E.; Rinkevich, B. Piwi positive cells that line the vasculature epithelium underlie whole body regeneration in a basal chordate. Dev. Biol. 2010, 345, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Struck, T.H.; Wey-Fabrizius, A.R.; Golombek, A.; Hering, L.; Weigert, A.; Bleidorn, C.; Klebow, S.; Iakovenko, N.; Hausdorf, B.; Petersen, M.; et al. Platyzoan Paraphyly Based on Phylogenomic Data Supports a Noncoelomate Ancestry of Spiralia. Mol. Biol. Evol. 2014, 31, 1833–1849. [Google Scholar] [CrossRef] [Green Version]
- Kozin, V.V.; Kostyuchenko, R.P. Vasa, PL10, and Piwi gene expression during caudal regeneration of the polychaete annelid Alitta virens. Dev. Genes Evol. 2010, 225, 129–138. [Google Scholar] [CrossRef]
- Coe, W.R. Biology of the nemerteans of the Atlantic coast of North America. Trans. Conn. Acad. Arts Sci. 1943, 35, 129–328. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.-M.; Sun, S.-C. Expression of Piwi Genes during the Regeneration of Lineus sanguineus (Nemertea, Pilidiophora, Heteronemertea). Genes 2020, 11, 1484. https://doi.org/10.3390/genes11121484
Xu C-M, Sun S-C. Expression of Piwi Genes during the Regeneration of Lineus sanguineus (Nemertea, Pilidiophora, Heteronemertea). Genes. 2020; 11(12):1484. https://doi.org/10.3390/genes11121484
Chicago/Turabian StyleXu, Cong-Mei, and Shi-Chun Sun. 2020. "Expression of Piwi Genes during the Regeneration of Lineus sanguineus (Nemertea, Pilidiophora, Heteronemertea)" Genes 11, no. 12: 1484. https://doi.org/10.3390/genes11121484
APA StyleXu, C. -M., & Sun, S. -C. (2020). Expression of Piwi Genes during the Regeneration of Lineus sanguineus (Nemertea, Pilidiophora, Heteronemertea). Genes, 11(12), 1484. https://doi.org/10.3390/genes11121484