1. Introduction
Plants respond to abiotic and biotic stresses and developmental stimuli by transcriptional reprogramming through constitutive and alternative RNA splicing events. RNA splicing is a highly ordered and dynamic posttranscriptional modification [
1] catalyzed by a highly conserved ribonucleoprotein complex known as the spliceosome comprising five small nuclear ribonucleoproteins (snRNPs) (U1, U2, U4/U6, and U5) and numerous non-snRNP proteins, including serine/arginine-rich (SR) proteins [
2,
3]. The SR proteins have a modular structure (20 to 75 kDa) consisting of one or two N-terminal RNA recognition motifs (RRMs) and serine and arginine-rich C-terminal domains (RS domains with multiple RS dipeptide repeats) [
4] that can bind and interact with target pre-mRNAs and proteins to modulate splicing [
5]. The SR protein and pre-mRNA interactions either activate or repress constitutive and alternative splicing for the maintenance of cellular and tissue homeostasis [
6,
7]. The phosphorylation and/or dephosphorylation of the RS domain alters the binding affinity of SR proteins for pre-mRNAs and other proteins, thus affecting splicing differently [
8]. Protein kinases phosphorylate SR proteins, which initiate spliceosome complex formation [
3]. The spliceosome performs the two transesterification reactions that are necessary to excise introns and join together the selected exons [
1]. In response to stress, plant SR genes initially undergo alternative splicing prior to the alternative splicing of the target genes, including stress response genes [
6]. The stress-dependent alternative splicing of SR proteins, which are key splice factors for protein-encoding genes, have distinct biological functions [
9,
10], suggesting the existence of a posttranscriptional level regulatory mechanism for stress response genes [
11].
The nomenclature of SR proteins has been designed solely according to their sequence properties; they comprise one or two N-terminal RRMs (RBDs; PF00076) followed by a downstream RS domain of at least 50 amino acids with 40% RS content characterized by consecutive RS or SR repeats [
12]. Nonetheless, two RS domains at the N- and the C-termini are observed in
atSR45a [
13]. Moreover, some AS events in
A. thaliana genes and in three Brassica species (
B. rapa, B. oleracea, and
B. napus) have been evolutionarily conserved [
14]. The model plant
Arabidopsis thaliana encodes 18 SR proteins, while
Brassica rapa has 25 SR genes (BrSR) that are classified into six subfamilies, named SR, RSZ, SC, SCL, RS2Z, and RS according to their distinct domain organization; most of them are responsive to oxidative, cold, and heat stress [
15]. The SR genes are known to be actively alternatively spliced under stress conditions [
16]. Heat and drought stress induce the alternative splicing of
atSR45a (AT1G07350.1 and AT1G07350.2) and enhance its expression [
17]. The levels of the transcripts
atSR45a, atSR30, and
SF2/ASF-like SR were increased by high-light irradiation and salinity stress [
13]. Nonetheless, the determination of the
SR45a target genes and their splicing patterns under particular stress conditions will provide vital clues as to their functional role in stress responses, which have yet to be identified. SR proteins participate in multiple signaling pathways, including the glucose signaling pathway [
18], the nonsense-mediated mRNA decay pathway (NMD), which is involved in the RNA surveillance system [
19], the ABA signaling pathway [
20], the autonomous flowering pathway [
21], and stress response gene expression pathways. There is mounting evidence implicating
AtSR genes in the signal transduction of the abscisic acid (ABA) phytohormone, a key modulator of various abiotic stress responses (drought, high salinity, light, cold, and heat) in plants [
5,
22]. In addition, SR-mediated pre-mRNA splicing processes were also influenced in a concentration-dependent manner [
16]. As central coordinators of plant abiotic stress responses, SR genes have drawn the attention of plant researchers focusing on the development of stress-resilient crops. Recently, a high-throughput approach was implemented to profile the binding targets of SR proteins, which are crucial stress response splicing factors in plants [
19]. For example,
AtSR45 either directly or indirectly interacts with 4000 RNAs/genes, which mostly include abscisic acid (ABA) signaling genes [
23]. Herein, we attempted to characterize the functions of the
BrSR45a gene (also known as
BrSR-like 3) in drought and heat stress responses by gathering phenotypic and molecular evidence supporting our hypothesis from
BrSR45a overexpressors and mutant and control Arabidopsis lines. This study will increase our understanding of the molecular underpinnings of the relationship between
BrSR45a and drought stress tolerance.
3. Results
Herein, we attempted to characterize BrSR45a through its heterologous expression in Arabidopsis. We investigated the impact of its overexpression and loss of function on the leaf and root phenotypes, chlorophyll a fluorescence, stress responses to drought and heat stress and, finally, the alternative splicing of genes that have been implicated in drought tolerance and BrSR45a interacting partners. In addition, we also analyzed the spatial expression of BrSR45a promoters in transgenic reporter lines.
3.1. Abiotic Stress Response Promoter Motifs and GUS Activity in Transgenic Reporter Lines (BrSR45a::GUS)
The promoter analysis showed that the SR45a promoter sequences had three important cis-acting elements that were responsive to abiotic stress. The promoter motifs were ABRE (GTGCAT), MYB (TAACTG), and HSE (AAAAATTTTC), which are known to participate in plant responses to ABA, drought and heat stress, respectively. Our preliminary analysis revealed that the exogenous application of stress hormones, ABA, heat and drought stress for different time intervals regulated
BrSR45a expression (
Figure 1).
The presence of promoter motifs and
SR45a regulation by ABA and drought stress indicated that the stress responses of
SR45a might be mediated by its promoter motifs. To understand the spatiotemporal expression of
SR45a, transgenic Arabidopsis reporter lines expressing GUS via the promoter of
BrSR45a were developed (
Figure 2a). Histochemical localization in transgenic reporter lines revealed that
BrSR45a expression was prominent in shoots, vascular root tissues, and the primary and secondary veins of developing leaves in young plants (
Figure 2b). The presence of GUS activity proved the role of
SR45a in plant development.
3.2. Phylogeny, Phenotypes, and Drought Stress Responses of BrSR45a
The phylogenetic analysis of the BrSR45a (BrSR-like 3) protein sequence (361 AA) in Brassica and Brassicaceae members (
Figure 3a) showed a fair amount of conservation in the amino acid residues. Further analysis revealed that
BrSR45a is more closely related to that in
B. campestris in Brassica and
Camelina sativa and
Raphanus sativus in Brassicaceae. Brassica members, such as
B. oleracea and
B. cretica, were placed in different clades, indicating that evolutionary divergence occurred during natural selection among Brassica members. To functionally characterize the
BrSR45agene, it was PCR-amplified from
B. rapa cv. Dongbu and overexpressed in
A. thaliana (Col-0) using Agrobacterium-mediated genetic transformation (
Figure 3b, c), and the resulting plants were compared in terms of their phenotype with SR45a knock-out mutants and the control Col-0.
No aberrant phenotypes were observed in them, except that the leaves in the mutant were slightly narrower than those in the
BrSR45a-overexpressing and Col-0 Arabidopsis lines; however, no significant difference between OE and Col-0 was found for leaf length and leaf width (
Figure 3d,e). Similarly, no difference was observed in the root phenotypes among them, although SR45a:
GUS constructs showed activity in vascular root tissues. However, we developed our main interest in the drought stress responses of OEs and mutants based on preliminary studies of their responses to drought, the stress hormone ABA, and promoter motifs (
Figure 1). We exposed OE, mutant and Col-0 lines to progressive drought stress for 17 consecutive days by suspending irrigation. The drought-induced subsequent stress was confirmed through the measurement of molecular markers of drought and oxidative stress. The expression of stress markers (
DREB2A, APX6, MDAR1, DHAR1, and
GR3) during drought stress was progressively upregulated, as shown via quantitative PCR assays (
Supplementary Figure S5). To measure the drought stress impact on photosystem II in each phenotype, chlorophyll a fluorescence transients (OJIPs) in dark-adopted leaves during progressive drought stress were measured (
Supplementary Figure S3). The comparative analysis of the OJIPs showed that the O-J, J-I, and I-P phases in the overexpressors were positive and relatively higher than those in the controls, Col-0 and SR45a mutants during drought stress. Interestingly, all phases of OJIP in mutants were reduced under stress in comparison with that in control plants. Moreover, the comparative drought phenotyping indicated that OEs could tolerate the dehydration imposed by soil moisture deficit stress, whereas mutants and Col-0 succumbed to stress and collapsed completely under stress conditions (
Figure 4a–d). Upon rewatering, OE lines with the highest expression (
Supplementary Figure S6) showed a better recovery rate than OE lines with moderate expression. Based on these results, we presumed that SR45a might participate in drought stress tolerance and/or stress recovery mechanisms, but its efficiency may depend on the magnitude of the expression level. The preliminary study of
BrSR45a in
B. rapa also showed that drought stress induced its expression (
Figure 1). Exposure to exogenous ABA also induced the expression of
BrSR45a, suggesting that
BrSR45a might play a role in both drought and ABA responses.
3.3. Expression Pattern of Drought and Antioxidative Stress Response Genes
The literature shows that stress response gene expression determines drought stress responses, including tolerance and avoidance, in crop plants. Therefore, we attempted to measure the expression of some of the well-known genes in the drought stress response pathway, namely,
AtDREB2A, AtNCED3, AtKIN1, and
AtRD29A. The qRT-PCR-based relative quantification results (
Figure 5a–d) showed that all 4 genes were upregulated in transgenic lines compared with control plants, indicating that the overexpression of
BrSR45a might induce stress response gene expression in transgenic plants. Interestingly, their expression in transgenic lines was significantly declined under drought conditions when compared to control lines.
The drought tolerance of plants is partially attributed to their inbuilt antioxidative system, comprising several antioxidants and free radical scavenging enzymes. Hence, we also investigated the expression of antioxidative genes such as
AtAPX1, AtGR1, AtSOD1, AtMDAR1, and
AtDHAR1 (
Supplementary Figure S7). In contrast to drought stress response gene expression, oxidative stress response gene expression showed no significant variation between transgenic and control lines under normal conditions. Consistent with the
AtSOD1 expression pattern, SOD activity in transgenic and control lines showed no statistically significant variation under normal conditions. However, during drought stress (7 d), the SOD activity in transgenic lines was slightly reduced, while 5 days of drought exposure caused no change in SOD activity (
Supplementary Figure S8).
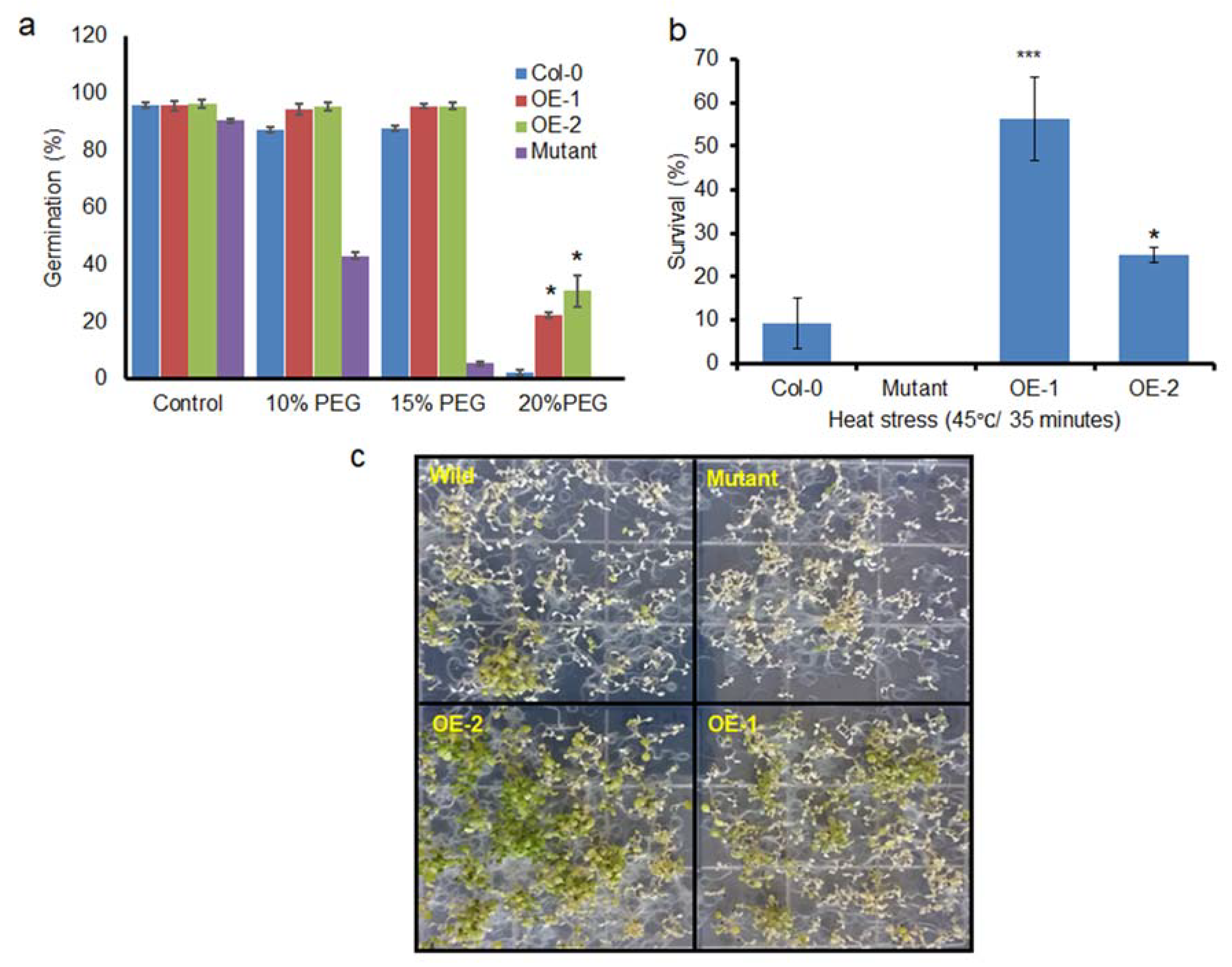
3.4. BrSR45a Overexpression Improved Basal Drought Tolerance in Seed Germination
Seed germination is one of the most critical stages for plant survival; hence, the basal stress tolerance efficiency of transgenic lines during seed germination was evaluated under PEG-induced drought stress conditions (
Figure 6a). Sterilized seeds from OEs and Col-0 were sown on 3MM filter papers that were moistened with different percentages of PEG solution. The germination assays revealed that there was no increase in the variation of the germination rate in the presence of 10–15% PEG compared with that in the control; however, the germination rate of mutants at 15% and 20% and the germination rate of Col-0 at 20% PEG were reduced to a greater extent than in the transgenic lines. The OE lines showed a higher drought tolerance index (DTI; calculated as mentioned in
Section 2.4) that ranged from 25–38%, while Col-0 and mutants showed a zero DTI during 20% PEG-induced drought stress. It is evident from the results that seed germination was constrained by PEG-induced drought stress. A high concentration (20%) of PEG almost completely inhibited the germination of the control, while 22.2% of the OE-1 line seeds and 34.38% of the OE-2 line seeds germinated, indicating that the overexpression of
BrSR45a improved the basal drought tolerance level in transgenic plants in the presence of drought stress.
3.5. Basal therMotolerance of Transgenic Seedlings
The ability of seedlings to survive and remain photosynthetically active from heat shock stress can be considered as basal heat stress tolerance. As described previously [
26], 9-d-old seedlings of OEs, mutants, and controls grown on plates were immersed in a water bath for 35 min at 45 °C. Heat stress tolerance was estimated as the percentage of surviving seedlings after a recovery period of 8 d at 23 °C (
Figure 6b,c). The results showed that seedling viability was completely lost in
SR45a loss of function mutants, while more than 50% of the OE-2 seedlings remain viable when subjected to heat stress. Similarly, around 20% of the OE-1 seedlings withstood heat stress and stayed with green leaves, the rate of survivability was higher in comparison with control seedlings. The heat phenotypes of OE-2 and OE-1 were consistent with
BrSR45a expression pattern indicating
BrSR45a possibly associated with heat stress responses.
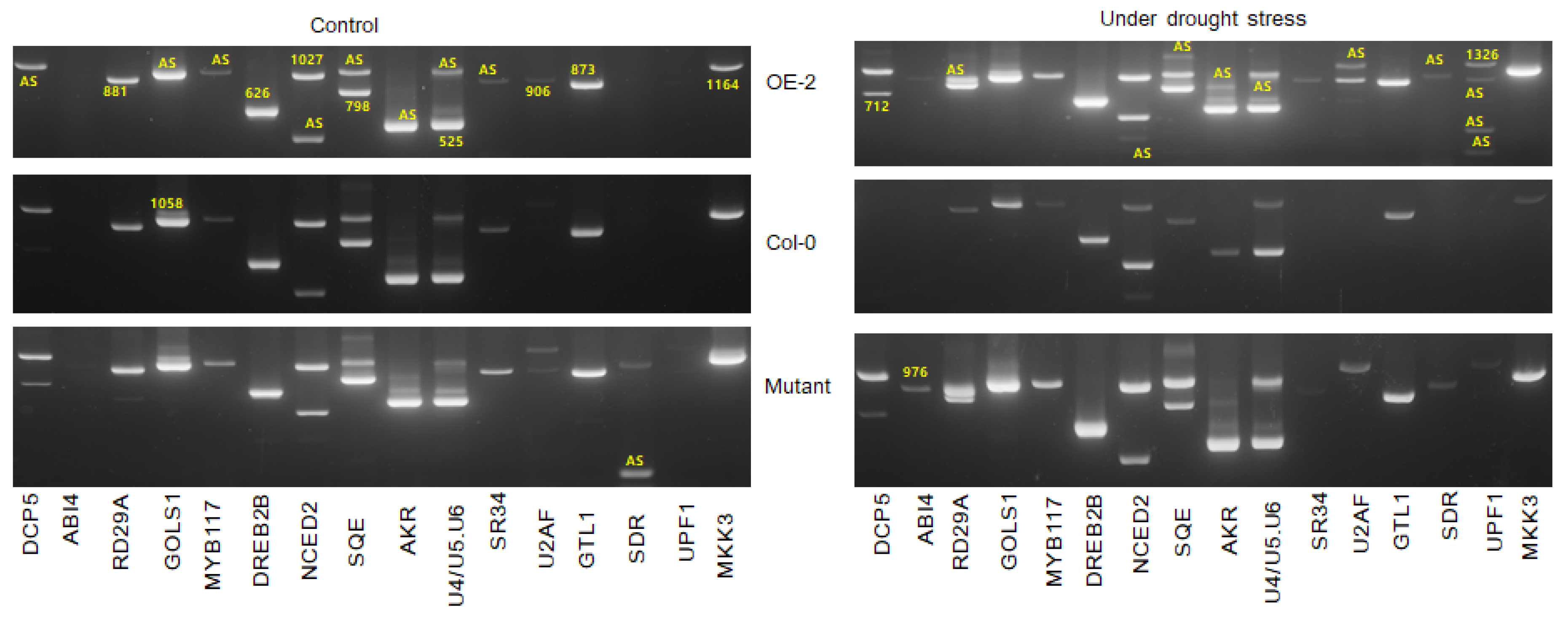
3.6. Disruption or Overexpression of SR45a Alters the Splicing Pattern of Genes Interacting with SR45a and Drought Stress Response Target Genes
The impact of the overexpression and loss of function of SR45a on the splicing patterns of drought tolerance genes were investigated (
Figure 7). The splicing pattern of all the genes in OEs and Col-0 under normal conditions appeared almost similar, while the differential splicing of
DCP5, RD29A, GOLS1, AKR, and
SDR were observed between OEs and mutants.
Moreover, the overexpression of SR45a did not alter the normal splicing of those genes unless the plants were exposed to drought stress conditions. Interestingly, the splicing patterns of most genes in the mutants under normal conditions were similar to those in the drought-stressed OE lines, suggesting that a loss of function would trigger stress response-induced alternative splicing. In general, the splicing patterns of DCP5, RD29A, GOLS1, NCED2, SQE, AKR, and UPF1 in OEs and NCED2, SQE, AKR, and UPF1 in mutants were different from those in Col-0, indicating that these genes could play a vital role in stress responses and alternative splicing when plants encountered stress signals from the environment. In comparison, the splicing patterns of NCED2, AKR, and UPF1 were different between OEs and mutants, while all other tested genes showed a similar splicing pattern during drought stress. Two additional splice variants with slightly higher molecular sizes than the original amplicons were observed for AKR in the OE lines; however, the challenge is to uncover the precise functions of each splice variant, warranting the characterization of the splice variants. Some classes of AKRs were previously implicated in osmolyte production, which confers drought tolerance. Although no clear information is available for the mechanism by which the higher molecular weight splice variants of AKR were produced, the molecular weight of new splice variants were higher than the original transcripts. We found four variants of the UPF1 gene during drought stress in OE, and the size of the variant was much smaller than the expected product size, presumably indicating the presence of truncated transcripts. However, further characterization is needed to understand their functional roles in drought stress responses. Moreover, we revealed the functional linkage between SR45a and key post-transcriptional modifications via the NMD pathway through the alternative splicing of UPF1. In comparison with those in the respective controls, the splicing patterns of DCP5, RD29A, NCED2, SQE, AKR, and UPF1 in OEs and RD29A, GOLS1, and SDR were different during drought conditions. Among the drought-tolerance genes, the splicing of DCP5, SQE, AKR, SDR, GOLS1, NCED2, UPF1, and RD29A were different under normal and drought stress conditions, suggesting that the alternative splicing of those genes is regulated by SR45a.
Apart from drought-responsive genes, SR45a also differentially alters the splicing of its interacting partners such as U2 SnRNP auxiliary splicing factor small subunit (Bra010926.1) and U4/U6-U5 tri-snRNP-associated protein 2-like (Bra013613.1) in OEs and mutants. In OEs, U2AF35A was alternatively spliced and produced 2 mRNA variants, possibly altering the splicing of other genes. The importance of splice variants during drought stress is unknown. Nonetheless, it is confirmed that the altered expression of BrSR45a controls the splicing pattern of U2AF in response to drought stress. Similarly, Bra013613.1 produced three and two splice variants in OEs and mutants, respectively, during drought stress. These results suggest that BrSR45a alters the splicing of its interacting partners in addition to that of other target genes.
4. Discussion
Our previous study showed that abiotic stresses—including salt, drought, oxidative and heat shock—upregulate the expression of
BrSR45a in
B. rapa [
15]. Herein, we characterized the functional role of
BrSR45a in drought and heat stress responses. The promoter analysis showed that the SR45a promoter sequences had three important cis-acting elements that were responsive to abiotic stress. Further, the exogenous application of stress hormones, ABA, heat, and drought stress regulated
BrSR45a expression (
Figure 1). Previously, high-light irradiation and salinity stress were shown to increase the expression of
atSR45a [
13]. These results clearly indicate that
SR45a play a critical role in abiotic stress responses of plants. The phenotyping of SR45a knock-out mutants with either control or
BrSR45a overexpressors revealed no aberrant phenotypes for leaves and roots between them. This result corroborates the earlier findings [
27], where the phenotypes of
AtSR45a mutant lines were similar to that of wild types. However, difference in photosynthesis rate and degree of tolerance between mutants and overexpressors was observed during drought stress. The comparative analysis of the OJIPs showed that the O-J, J-I, and I-P phases in the overexpressors were positive and relatively higher than those in mutants indicating SR45a associated with photosynthesis efficiency under stress conditions. Maintaining a better photosynthesis rate under stress conditions expected to have positive impact on drought tolerance [
27]. In fact, the first report on SR45a had demonstrated positive correlation between SR45a overexpression and photosynthesis efficiency in Arabidopsis [
28].
Drought tolerance is an important agronomic trait contributed by a complex network of genetic factors. The role of ABA in drought tolerance is inevitable because of its critical role in modulating stress responsive gene expression to coordinate plant stress responses, growth, and development under adverse conditions. AtSR45a also participates in ABA signal transduction to modulate abiotic stress responses [
5,
22,
29]. In this study, we found that expression of stress-related genes such as
AtDREB2A, AtNCED3, AtKIN1, and
AtRD29A were upregulated in transgenic lines compared with control plants, indicating that the overexpression of
BrSR45a might induce stress response gene expression in transgenic plants. Previous studies on
DREB2A, NCED3, and
RD29A showed that their overexpression in
Robinia pseudoacacia, grapevines and
A. thaliana, respectively, were associated with improved tolerance to drought stress [
30,
31,
32]. KIN1 produces LEA class proteins that aid in damage repair during stress [
33]. Therefore, the induced expression of stress-related genes in SR45a overexpressors expected to alleviate the drought stress mediated negative impacts in plants [
34]. Contrastingly, oxidative stress response gene expression (e.g.,
APX1,
SOD1,
DHAR, and
MDAR) showed no significant variation between transgenic and control lines under normal conditions. Consistent with the
AtSOD1 expression pattern, SOD activity in transgenic and control lines showed no statistically significant variation under normal conditions suggesting that the expression of
SR45a and oxidative stress are not correlated. Interestingly,
SR45a homologs in Arabidopsis was unperturbed by reactive oxygen species [
13] indicating possible shared functions for SR45a genes between these species. Furthermore, it is confirmed that BrSR45a do not alternatively spliced under hydrogen peroxide mediated oxidative stress while splice variants in
B. rapa were reported [
15]
Seed germination is one of the most critical stages for plant survival [
25] and germination rate under drought stress indicates the degree of basal tolerance mechanism [
35]. The overexpression of
BrSR45a in transgenic lines had germination rate around 22–34% while the germination of mutants and control was completely inhibited in 15% or 20% PEG-induced drought stress. It is evident from the results that the overexpression of
BrSR45a improved the basal drought tolerance level in transgenic plants in the presence of drought stress. Therefore, we investigated the association of
BrSR45a with heat stress responses in this study by incubating 9-day-old seedlings of OEs, mutants, and controls grown on plates 45 °C (
Figure 6c,d) for 35 min [
26]. The viability of OE-2, OE-1, control and
SR45a mutants were positively correlated with SR45a expression under stress conditions. Transgenic lines with highest expression of
BrSR45a had better survivability while the loss of function causes severe susceptibility to heat stress. This result clearly indicates that
BrSR45a overexpression resulted in heat-tolerant phenotypes in Arabidopsis seedlings. However, it is not clear
BrSR45a or its splice isoforms induced during heat stress [
15] contributing tolerance phenotypes, and characterization of heat stress derived splice variants will shed light on the precise role in thermotolerance.
Notably, evidence from the literature has shown that stress response genes in plants undergo alternative splicing [
27]. SR45a proteins are important splicing factors in regulating gene expression through AS. The
SR45a-dependent alternative splicing of target protein-encoding genes might have distinct biological functions [
9,
10] or may simply operate as part of the mRNA decay pathway for RNA surveillance system during unfavorable conditions [
19]. To obtain knowledge on SR45a mediated alternative splicing of drought stress responsive genes both under normal and drought stress conditions, we investigated the splice variants of SR45a overexpressors, mutants, and controls. The comparative analysis of alternative splicing suggested that both enhanced activity and loss of activity change the splicing pattern in target genes but differentially. This suggests that SR45a is crucial for the splicing of target genes, including drought-responsive genes. Moreover, the overexpression of SR45a did not alter the normal splicing of those genes unless the plants were exposed to drought stress conditions. The splicing patterns of
NCED2, AKR, and
UPF1 were different between OEs and mutants during drought stress exposure. AKRs are primarily involved in the detoxification of toxic aldehydes and ketones produced during stress [
36]. NCED (9-cis-epoxycartoenoid dioxygenase), a key regulator of ABA biosynthesis and its overexpression, has been implicated in drought tolerance [
31]. UPF1, a key NMD factor, activates the NMD pathway upon its phosphorylation [
37]. However, the challenge is to uncover the precise functions of each splice variant, warranting the characterization of the splice variants. Nevertheless, some classes of AKRs were previously implicated in osmolyte production, which confers drought tolerance. The splice variants of
AKR were slightly higher in molecular weight than the original amplicons in the OE lines. The possibility of intron retention being the prime reason for higher molecular weight splice variants can be ruled out on the basis of earlier findings that the high-light induced expression of
atSR45a actively suppressed the efficiency of the intron-retention type mechanism [
27]. In some cases, these transcripts can be degraded through the NMD pathway [
38]. Despite that, SR45a-induced alternative splicing can be linked with stress tolerance in transgenic plants [
39].
Apart from drought-responsive genes,
SR45a also differentially alters the splicing of its interacting partners (
U2 SnRNP auxiliary splicing factor small subunit (Bra010926.1) and
U4/U6-U5 tri-snRNP-associated protein 2-like (Bra013613.1)) in OEs and mutants. A change in the expression of the U2 snRNP auxiliary splicing factor produced no variation in phenotypes [
40] indicates other regulatory roles although the mechanism is not clear. Additionally, studies have documented that
SR45a participates in the multiples roles, including glucose signaling pathway [
18], the nonsense-mediated mRNA decay pathway (NMD) in the RNA surveillance system [
19], ABA signaling [
20], the autonomous flowering pathway [
21], and stress response gene expression pathways. In this study, we showed that
BrSR45a is involved in drought stress responses through the alternative splicing of drought stress response genes. In addition, we found that the overexpression of
BrSR45a protects photosystem II during drought stress and improves basal thermotolerance in young seedlings. In conclusion,
SR45a overexpression is positively correlated with enhanced drought tolerance, thermotolerance, and drought recovery rates compared with that observed in the
SR45a mutant and Col-0 in an expression-dependent manner. Moreover, the OE and mutant lines differentially regulated several known drought-responsive genes, possibly contributing to enhanced drought tolerance. Although a precise understanding that supports SR45a-mediated drought tolerance is lacking, this study revealed that
BrSR45a mediated the alternative splicing of target genes and their interacting partner genes to possibly contribute to stress tolerance in a concentration-dependent manner. By discovering the role of SR45a in drought and heat stress responses and the alternative splicing of target genes during stress responses, this work will be helpful for the development of stress-resilient crops to improve crop productivity under adverse environmental conditions.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






