Transcriptomic Profiling Identifies Differentially Expressed Genes in Palbociclib-Resistant ER+ MCF7 Breast Cancer Cells

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Generation of Palbociclib-Resistant Cells and Palbociclib Treatment
2.2. RNA Extraction and Next-Generation Sequencing
2.3. DEG Analysis
2.4. In Silico Ingenuity Network Analysis
2.5. GFP-LC3 Visualization
3. Results
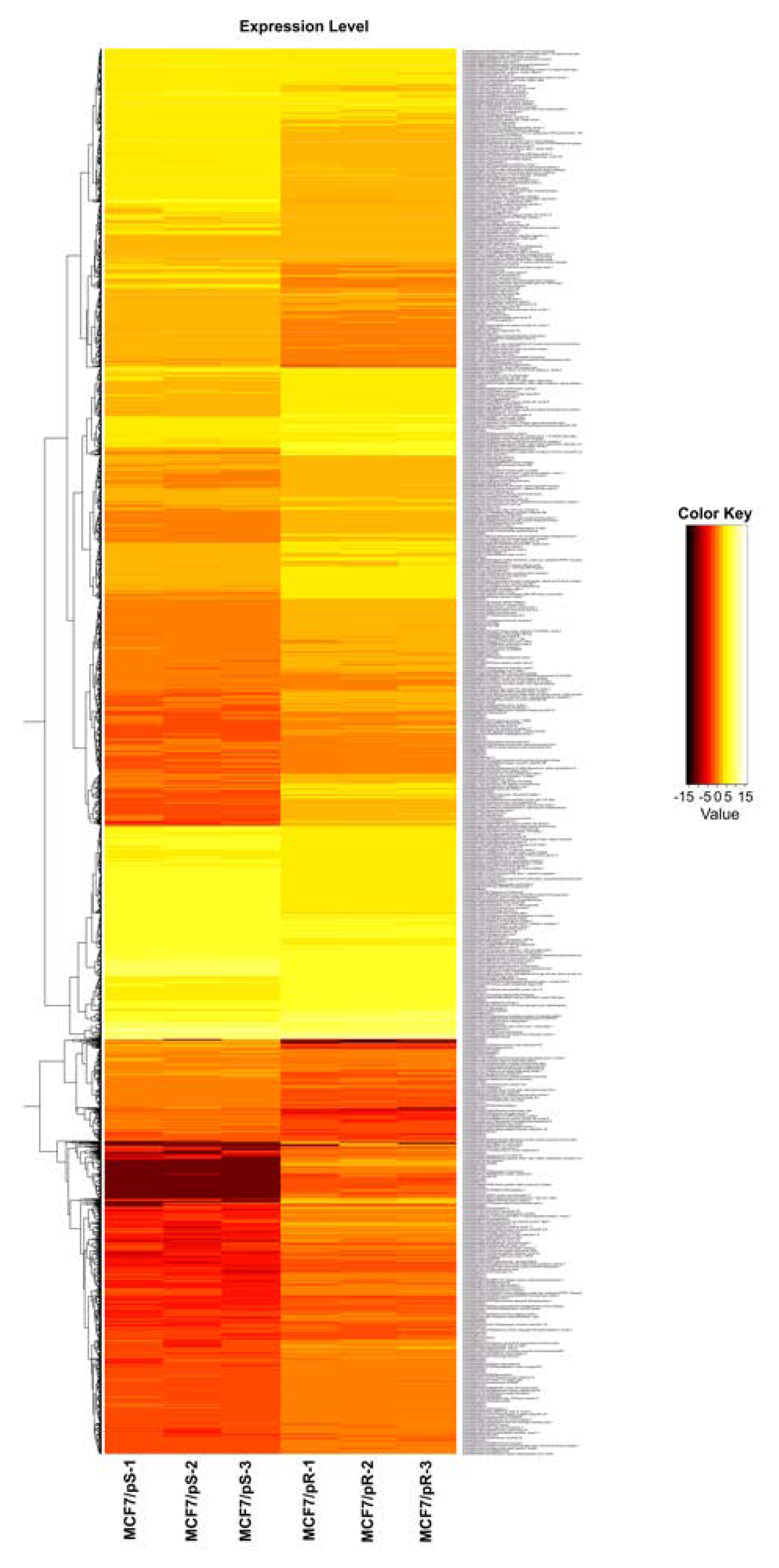
3.1. RNA-Seq Profiling Reveals a Distinct Transcriptomic Profiling in Palbociclib-Resistant Cells
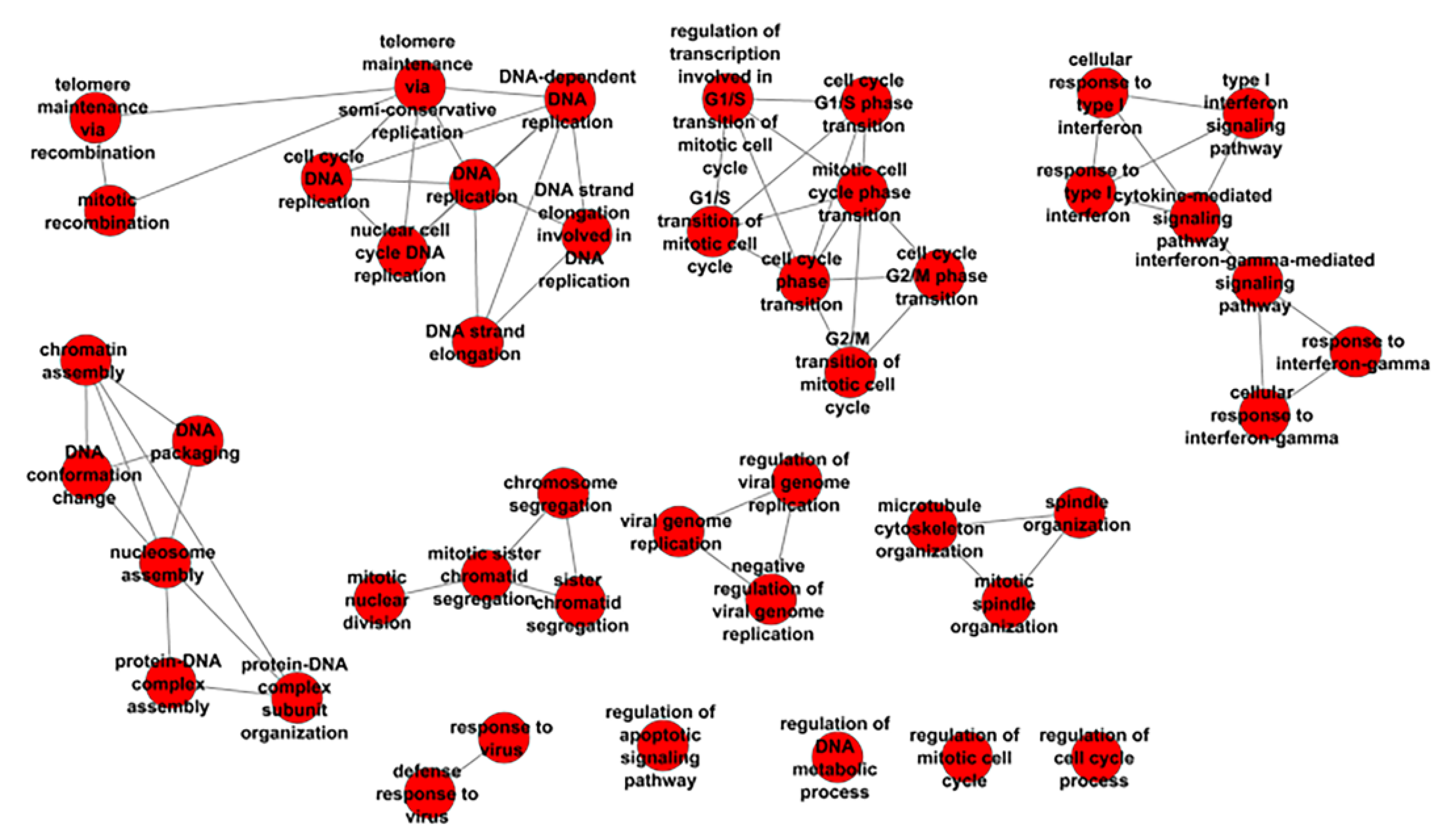
3.2. KEGG Annotation of DEG and Enriched Biological Processes Analysis
3.3. Resistance to Palbociclib Is Associated with Increased Autophagosome Formation
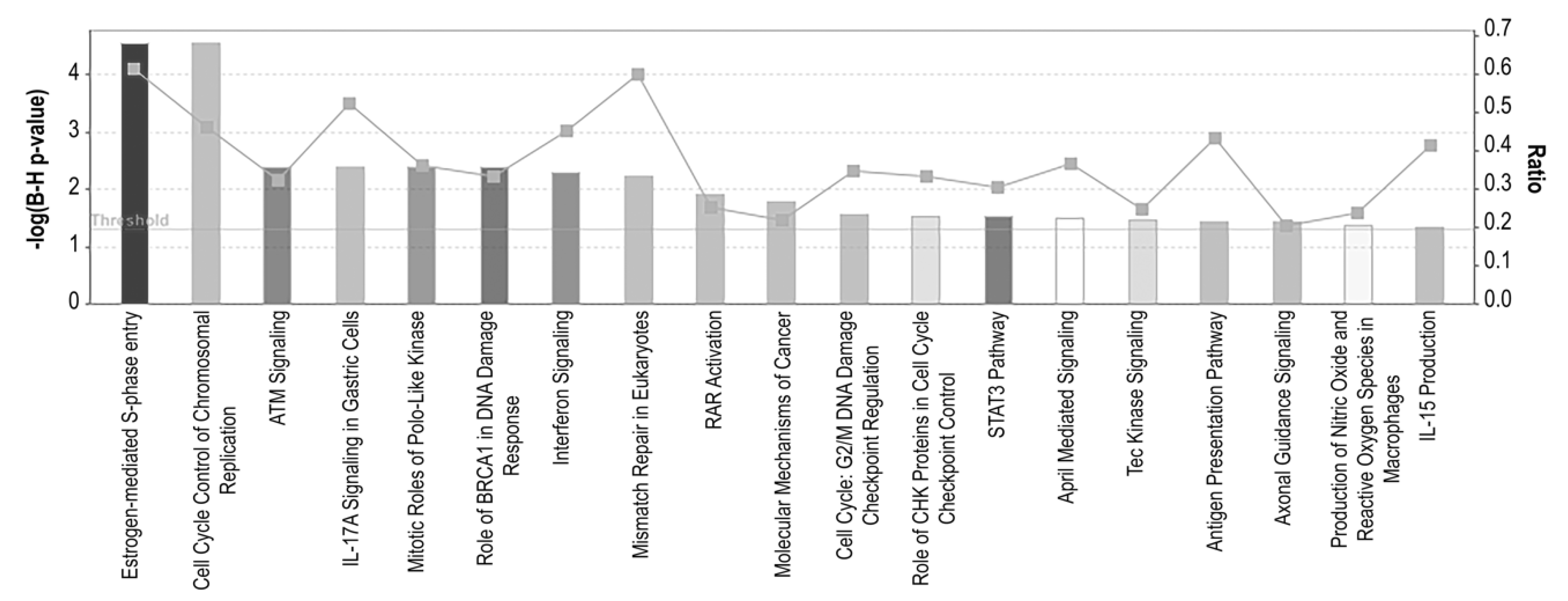
3.4. Pathway Enrichment Analysis of DEG
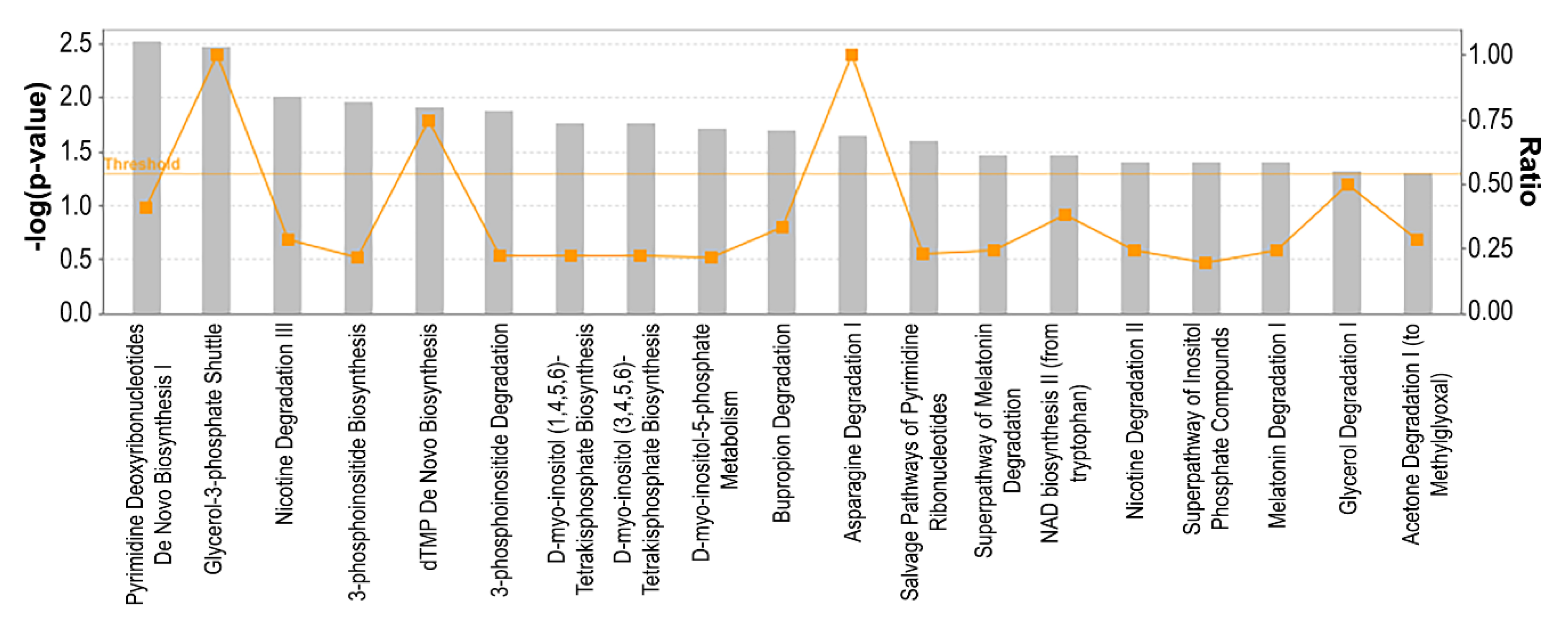
3.5. Metabolic Pathways Associated with Resistance to Palbociclib
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rossi, L.; McCartney, A.; Risi, E.; De Santo, I.; Migliaccio, I.; Malorni, L.; Biganzoli, L.; Di Leo, A. Cyclin-Dependent Kinase 4/6 Inhibitors in Neoadjuvant Endocrine Therapy of Hormone Receptor-Positive Breast Cancer. Clin. Breast Cancer 2019, 19, 392–398. [Google Scholar] [CrossRef]
- Sobhani, N.; D’Angelo, A.; Pittacolo, M.; Roviello, G.; Miccoli, A.; Corona, S.P.; Bernocchi, O.; Generali, D.; Otto, T. Updates on the CDK4/6 Inhibitory Strategy and Combinations in Breast Cancer. Cells 2019, 8, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wang, Q.; Wang, Q.; Cao, J.; Sun, J.; Zhu, Z. Mechanisms of resistance to estrogen receptor modulators in ER+/HER2- advanced breast cancer. Cell. Mol. Life Sci. 2020, 77, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Abreu, M.T.; Palafox, M.; Asghar, U.; Rivas, M.A.; Cutts, R.J.; Garcia-Murillas, I.; Pearson, A.; Guzman, M.; Rodriguez, O.; Grueso, J.; et al. Early Adaptation and Acquired Resistance to CDK4/6 Inhibition in Estrogen Receptor-Positive Breast Cancer. Cancer Res. 2016, 76, 2301–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Li, Z.; Bhatt, T.; Dickler, M.; Giri, D.; Scaltriti, M.; Baselga, J.; Rosen, N.; Chandarlapaty, S. Acquired CDK6 amplification promotes breast cancer resistance to CDK4/6 inhibitors and loss of ER signaling and dependence. Oncogene 2017, 36, 2255–2264. [Google Scholar] [CrossRef] [Green Version]
- Lypova, N.; Lanceta, L.; Gibson, A.; Vega, S.; Garza-Morales, R.; McMasters, K.M.; Chesney, J.; Gomez-Gutierrez, J.G.; Imbert-Fernandez, Y. Targeting Palbociclib-Resistant Estrogen Receptor-Positive Breast Cancer Cells via Oncolytic Virotherapy. Cancers (Basel) 2019, 11, 684. [Google Scholar] [CrossRef] [Green Version]
- Condorelli, R.; Spring, L.; O’Shaughnessy, J.; Lacroix, L.; Bailleux, C.; Scott, V.; Dubois, J.; Nagy, R.J.; Lanman, R.B.; Iafrate, A.J.; et al. Polyclonal RB1 mutations and acquired resistance to CDK 4/6 inhibitors in patients with metastatic breast cancer. Ann. Oncol. 2018, 29, 640–645. [Google Scholar] [CrossRef]
- O’Leary, B.; Hrebien, S.; Morden, J.P.; Beaney, M.; Fribbens, C.; Huang, X.; Liu, Y.; Bartlett, C.H.; Koehler, M.; Cristofanilli, M.; et al. Early circulating tumor DNA dynamics and clonal selection with palbociclib and fulvestrant for breast cancer. Nat. Commun. 2018, 9, 896. [Google Scholar] [CrossRef]
- O’Leary, B.; Cutts, R.J.; Liu, Y.; Hrebien, S.; Huang, X.; Fenwick, K.; Andre, F.; Loibl, S.; Loi, S.; Garcia-Murillas, I.; et al. The Genetic Landscape and Clonal Evolution of Breast Cancer Resistance to Palbociclib plus Fulvestrant in the PALOMA-3 Trial. Cancer Discov. 2018, 8, 1390–1403. [Google Scholar] [CrossRef] [Green Version]
- Formisano, L.; Lu, Y.; Servetto, A.; Hanker, A.B.; Jansen, V.M.; Bauer, J.A.; Sudhan, D.R.; Guerrero-Zotano, A.L.; Croessmann, S.; Guo, Y.; et al. Aberrant FGFR signaling mediates resistance to CDK4/6 inhibitors in ER+ breast cancer. Nat. Commun. 2019, 10, 1373. [Google Scholar] [CrossRef] [Green Version]
- Jansen, V.M.; Bhola, N.E.; Bauer, J.A.; Formisano, L.; Lee, K.M.; Hutchinson, K.E.; Witkiewicz, A.K.; Moore, P.D.; Estrada, M.V.; Sanchez, V.; et al. Kinome-Wide RNA Interference Screen Reveals a Role for PDK1 in Acquired Resistance to CDK4/6 Inhibition in ER-Positive Breast Cancer. Cancer Res. 2017, 77, 2488–2499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, X.; Du, J.; Parsons, S.H.; Merzoug, F.F.; Webster, Y.; Iversen, P.W.; Chio, L.C.; Van Horn, R.D.; Lin, X.; Blosser, W.; et al. Aurora A Kinase Inhibition Is Synthetic Lethal with Loss of the RB1 Tumor Suppressor Gene. Cancer Discov. 2019, 9, 248–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kettner, N.M.; Vijayaraghavan, S.; Durak, M.G.; Bui, T.; Kohansal, M.; Ha, M.J.; Liu, B.; Rao, X.; Wang, J.; Yi, M.; et al. Combined Inhibition of STAT3 and DNA Repair in Palbociclib-Resistant ER-Positive Breast Cancer. Clin. Cancer Res. 2019, 25, 3996–4013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Leeuw, R.; McNair, C.; Schiewer, M.J.; Neupane, N.P.; Brand, L.J.; Augello, M.A.; Li, Z.; Cheng, L.C.; Yoshida, A.; Courtney, S.M.; et al. MAPK Reliance via Acquired CDK4/6 Inhibitor Resistance in Cancer. Clin. Cancer Res. 2018, 24, 4201–4214. [Google Scholar] [CrossRef] [Green Version]
- Illumina, I. BaseSpace User Guide; Part # 15044182 Rev. E; Ilumina: San Diego, CA, USA, 2014; pp. 1–98. [Google Scholar]
- Flight, R.M.; Harrison, B.J.; Mohammad, F.; Bunge, M.B.; Moon, L.D.; Petruska, J.C.; Rouchka, E.C. categoryCompare, an analytical tool based on feature annotations. Front. Genet. 2014, 5, 98. [Google Scholar] [CrossRef] [Green Version]
- Kabeya, Y.; Mizushima, N.; Ueno, T.; Yamamoto, A.; Kirisako, T.; Noda, T.; Kominami, E.; Ohsumi, Y.; Yoshimori, T. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J. 2000, 19, 5720–5728. [Google Scholar] [CrossRef]
- Shui, W.; Sheu, L.; Liu, J.; Smart, B.; Petzold, C.J.; Hsieh, T.Y.; Pitcher, A.; Keasling, J.D.; Bertozzi, C.R. Membrane proteomics of phagosomes suggests a connection to autophagy. Proc. Natl. Acad. Sci. USA 2008, 105, 16952–16957. [Google Scholar] [CrossRef] [Green Version]
- Vijayaraghavan, S.; Karakas, C.; Doostan, I.; Chen, X.; Bui, T.; Yi, M.; Raghavendra, A.S.; Zhao, Y.; Bashour, S.I.; Ibrahim, N.K.; et al. CDK4/6 and autophagy inhibitors synergistically induce senescence in Rb positive cytoplasmic cyclin E negative cancers. Nat. Commun. 2017, 8, 15916. [Google Scholar] [CrossRef]
- Chittaranjan, S.; Bortnik, S.; Dragowska, W.H.; Xu, J.; Abeysundara, N.; Leung, A.; Go, N.E.; DeVorkin, L.; Weppler, S.A.; Gelmon, K.; et al. Autophagy inhibition augments the anticancer effects of epirubicin treatment in anthracycline-sensitive and -resistant triple-negative breast cancer. Clin. Cancer Res. 2014, 20, 3159–3173. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Jiang, Y.Z.; Huang, L.; Zhou, R.J.; Yu, K.D.; Liu, Y.; Shao, Z.M. The residual tumor autophagy marker LC3B serves as a prognostic marker in local advanced breast cancer after neoadjuvant chemotherapy. Clin. Cancer Res. 2013, 19, 6853–6862. [Google Scholar] [CrossRef] [Green Version]
- Lefort, S.; Joffre, C.; Kieffer, Y.; Givel, A.M.; Bourachot, B.; Zago, G.; Bieche, I.; Dubois, T.; Meseure, D.; Vincent-Salomon, A.; et al. Inhibition of autophagy as a new means of improving chemotherapy efficiency in high-LC3B triple-negative breast cancers. Autophagy 2014, 10, 2122–2142. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Nicolay, B.N.; Chick, J.M.; Gao, X.; Geng, Y.; Ren, H.; Gao, H.; Yang, G.; Williams, J.A.; Suski, J.M.; et al. The metabolic function of cyclin D3-CDK6 kinase in cancer cell survival. Nature 2017, 546, 426–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, J.; Balaji, U.; Freinkman, E.; Witkiewicz, A.K.; Knudsen, E.S. Metabolic Reprogramming of Pancreatic Cancer Mediated by CDK4/6 Inhibition Elicits Unique Vulnerabilities. Cell Rep. 2016, 14, 979–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozeboom, B.; Dey, N.; De, P. ER+ metastatic breast cancer: Past, present, and a prescription for an apoptosis-targeted future. Am. J. Cancer Res. 2019, 9, 2821–2831. [Google Scholar]
- McCartney, A.; Migliaccio, I.; Bonechi, M.; Biagioni, C.; Romagnoli, D.; De Luca, F.; Galardi, F.; Risi, E.; De Santo, I.; Benelli, M.; et al. Mechanisms of Resistance to CDK4/6 Inhibitors: Potential Implications and Biomarkers for Clinical Practice. Front. Oncol. 2019, 9, 666. [Google Scholar] [CrossRef]
- Fry, D.W.; Harvey, P.J.; Keller, P.R.; Elliott, W.L.; Meade, M.; Trachet, E.; Albassam, M.; Zheng, X.; Leopold, W.R.; Pryer, N.K.; et al. Specific inhibition of cyclin-dependent kinase 4/6 by PD 0332991 and associated antitumor activity in human tumor xenografts. Mol. Cancer Ther. 2004, 3, 1427–1438. [Google Scholar]
- Finn, R.S.; Dering, J.; Conklin, D.; Kalous, O.; Cohen, D.J.; Desai, A.J.; Ginther, C.; Atefi, M.; Chen, I.; Fowst, C.; et al. PD 0332991, a selective cyclin D kinase 4/6 inhibitor, preferentially inhibits proliferation of luminal estrogen receptor-positive human breast cancer cell lines in vitro. Breast Cancer Res. 2009, 11, R77. [Google Scholar] [CrossRef] [Green Version]
- Verde, G.; De Llobet, L.I.; Wright, R.H.G.; Quilez, J.; Peiro, S.; Le Dily, F.; Beato, M. Unliganded Progesterone Receptor Governs Estrogen Receptor Gene Expression by Regulating DNA Methylation in Breast Cancer Cells. Cancers (Basel) 2018, 10, 371. [Google Scholar] [CrossRef] [Green Version]
- Barr, F.A.; Sillje, H.H.; Nigg, E.A. Polo-like kinases and the orchestration of cell division. Nat. Rev. Mol. Cell Biol. 2004, 5, 429–440. [Google Scholar] [CrossRef]
- Song, B.; Liu, X.S.; Liu, X. Polo-like kinase 1 (Plk1): An Unexpected Player in DNA Replication. Cell Div. 2012, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.B.; Im, J.H.; Yoon, J.H.; Bui, Q.T.; Lim, S.C.; Song, J.M.; Shim, Y.; Yun, J.; Hong, J.; Kang, K.W. Essential Role of Polo-like Kinase 1 (Plk1) Oncogene in Tumor Growth and Metastasis of Tamoxifen-Resistant Breast Cancer. Mol. Cancer Ther. 2018, 17, 825–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafer, D.; Grant, S. Update on rational targeted therapy in AML. Blood Rev. 2016, 30, 275–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudolph, D.; Steegmaier, M.; Hoffmann, M.; Grauert, M.; Baum, A.; Quant, J.; Haslinger, C.; Garin-Chesa, P.; Adolf, G.R. BI 6727, a Polo-like kinase inhibitor with improved pharmacokinetic profile and broad antitumor activity. Clin. Cancer Res. 2009, 15, 3094–3102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afratis, N.; Gialeli, C.; Nikitovic, D.; Tsegenidis, T.; Karousou, E.; Theocharis, A.D.; Pavao, M.S.; Tzanakakis, G.N.; Karamanos, N.K. Glycosaminoglycans: Key players in cancer cell biology and treatment. FEBS J. 2012, 279, 1177–1197. [Google Scholar] [CrossRef]
- Nikitovic, D.; Kouvidi, K.; Voudouri, K.; Berdiaki, A.; Karousou, E.; Passi, A.; Tzanakakis, G.N. The motile breast cancer phenotype roles of proteoglycans/glycosaminoglycans. Biomed. Res. Int. 2014, 2014, 124321. [Google Scholar] [CrossRef] [Green Version]
- Kasamatsu, A.; Uzawa, K.; Minakawa, Y.; Ishige, S.; Kasama, H.; Endo-Sakamoto, Y.; Ogawara, K.; Shiiba, M.; Takiguchi, Y.; Tanzawa, H. Decorin in human oral cancer: A promising predictive biomarker of S-1 neoadjuvant chemosensitivity. Biochem. Biophys. Res. Commun. 2015, 457, 71–76. [Google Scholar] [CrossRef]
- Pope, W.B.; Mirsadraei, L.; Lai, A.; Eskin, A.; Qiao, J.; Kim, H.J.; Ellingson, B.; Nghiemphu, P.L.; Kharbanda, S.; Soriano, R.H.; et al. Differential gene expression in glioblastoma defined by ADC histogram analysis: Relationship to extracellular matrix molecules and survival. AJNR Am. J. Neuroradiol. 2012, 33, 1059–1064. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.X.; Lu, C.Y.; Yang, Y.L.; Dou, K.F.; Tao, K.S. Lumican expression in pancreatic ductal adenocarcinoma. Hepatogastroenterology 2013, 60, 349–353. [Google Scholar] [CrossRef]
- Nikitovic, D.; Chalkiadaki, G.; Berdiaki, A.; Aggelidakis, J.; Katonis, P.; Karamanos, N.K.; Tzanakakis, G.N. Lumican regulates osteosarcoma cell adhesion by modulating TGFbeta2 activity. Int. J. Biochem. Cell Biol. 2011, 43, 928–935. [Google Scholar] [CrossRef]
- Yamano, Y.; Uzawa, K.; Saito, K.; Nakashima, D.; Kasamatsu, A.; Koike, H.; Kouzu, Y.; Shinozuka, K.; Nakatani, K.; Negoro, K.; et al. Identification of cisplatin-resistance related genes in head and neck squamous cell carcinoma. Int. J. Cancer 2010, 126, 437–449. [Google Scholar] [CrossRef]
- Kwon, W.S.; Kim, T.S.; Nahm, C.H.; Moon, Y.; Kim, J.J. Aberrant cystatin-C expression in blood from patients with breast cancer is a suitable marker for monitoring tumor burden. Oncol. Lett. 2018, 16, 5583–5590. [Google Scholar] [CrossRef] [PubMed]
- Decock, J.; Obermajer, N.; Vozelj, S.; Hendrickx, W.; Paridaens, R.; Kos, J. Cathepsin B, cathepsin H, cathepsin X and cystatin C in sera of patients with early-stage and inflammatory breast cancer. Int. J. Biol. Markers 2008, 23, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Moyano, J.V.; Evans, J.R.; Chen, F.; Lu, M.; Werner, M.E.; Yehiely, F.; Diaz, L.K.; Turbin, D.; Karaca, G.; Wiley, E.; et al. AlphaB-crystallin is a novel oncoprotein that predicts poor clinical outcome in breast cancer. J. Clin. Investig. 2006, 116, 261–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelouche-Lev, D.; Kluger, H.M.; Berger, A.J.; Rimm, D.L.; Price, J.E. alphaB-crystallin as a marker of lymph node involvement in breast carcinoma. Cancer 2004, 100, 2543–2548. [Google Scholar] [CrossRef]
- Ivanov, O.; Chen, F.; Wiley, E.L.; Keswani, A.; Diaz, L.K.; Memmel, H.C.; Rademaker, A.; Gradishar, W.J.; Morrow, M.; Khan, S.A.; et al. alphaB-crystallin is a novel predictor of resistance to neoadjuvant chemotherapy in breast cancer. Breast Cancer Res. Treat. 2008, 111, 411–417. [Google Scholar] [CrossRef]
- Zhang, P.; Tang, W.M.; Zhang, H.; Li, Y.Q.; Peng, Y.; Wang, J.; Liu, G.N.; Huang, X.T.; Zhao, J.J.; Li, G.; et al. MiR-646 inhibited cell proliferation and EMT-induced metastasis by targeting FOXK1 in gastric cancer. Br. J. Cancer 2017, 117, 525–534. [Google Scholar] [CrossRef]
- Xue, M.; Li, G.; Fang, X.; Wang, L.; Jin, Y.; Zhou, Q. hsa_circ_0081143 promotes cisplatin resistance in gastric cancer by targeting miR-646/CDK6 pathway. Cancer Cell Int. 2019, 19, 25. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, M.; Lee, J.Y.; Kim, M.H. CTCF negatively regulates HOXA10 expression in breast cancer cells. Biochem. Biophys. Res. Commun. 2015, 467, 828–834. [Google Scholar] [CrossRef]
- Ogrodzinski, M.P.; Bernard, J.J.; Lunt, S.Y. Deciphering metabolic rewiring in breast cancer subtypes. Transl. Res. 2017, 189, 105–122. [Google Scholar] [CrossRef]
- Lorito, N.; Bacci, M.; Smiriglia, A.; Mannelli, M.; Parri, M.; Comito, G.; Ippolito, L.; Giannoni, E.; Bonechi, M.; Benelli, M.; et al. Glucose Metabolic Reprogramming of ER Breast Cancer in Acquired Resistance to the CDK4/6 Inhibitor Palbociclib+. Cells 2020, 9, 668. [Google Scholar] [CrossRef] [Green Version]
- McCartney, A.; Bonechi, M.; De Luca, F.; Biagioni, C.; Curigliano, G.; Moretti, E.; Minisini, A.M.; Bergqvist, M.; Benelli, M.; Migliaccio, I.; et al. Plasma thymidine kinase activity as a biomarker in patients with luminal metastatic breast cancer treated with palbociclib within the TREnd trial. Clin. Cancer Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Romain, S.; Spyratos, F.; Descotes, F.; Daver, A.; Rostaing-Puissant, B.; Bougnoux, P.; Colonna, M.; Bolla, M.; Martin, P.M. Prognostic of DNA-synthesizing enzyme activities (thymidine kinase and thymidylate synthase) in 908 T1-T2, N0-N1, M0 breast cancers: A retrospective multicenter study. Int. J. Cancer 2000, 87, 860–868. [Google Scholar] [CrossRef]
- Broet, P.; Romain, S.; Daver, A.; Ricolleau, G.; Quillien, V.; Rallet, A.; Asselain, B.; Martin, P.M.; Spyratos, F. Thymidine kinase as a proliferative marker: Clinical relevance in 1,692 primary breast cancer patients. J. Clin. Oncol. 2001, 19, 2778–2787. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.X.; Gao, F.; Luo, J.; Northfelt, D.W.; Goetz, M.; Forero, A.; Hoog, J.; Naughton, M.; Ademuyiwa, F.; Suresh, R.; et al. NeoPalAna: Neoadjuvant Palbociclib, a Cyclin-Dependent Kinase 4/6 Inhibitor, and Anastrozole for Clinical Stage 2 or 3 Estrogen Receptor-Positive Breast Cancer. Clin. Cancer Res. 2017, 23, 4055–4065. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Up-Regulated | Down-Regulated | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Ensembl ID | Gene Symbol|Description | Log2FC | p-Value | Q-value | Ensembl ID | Gene Symbol|Description | Log2FC | p-Value | Q-Value |
| ENSG00000011465 | DCN|decorin | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000211816 | TRAV38-1| | −16.6898 | 5 × 10−5 | 0.00026 |
| ENSG00000011677 | GABRA3|gamma-aminobutyric acid (GABA) A receptor, alpha 3 | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000228340 | MIR646HG|MIR646 host gene | −16.6898 | 5 × 10−5 | 0.00026 |
| ENSG00000012223 | LTF|lactotransferrin | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000259761 | −16.6898 | 5 × 10−5 | 0.00026 | |
| ENSG00000070729 | CNGB1|cyclic nucleotide gated channel beta 1 | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000082482 | KCNK2|potassium channel, two pore domain subfamily K, member 2 | −7.09413 | 5 × 10−5 | 0.00026 |
| ENSG00000079689 | SCGN|secretagogin, EF-hand calcium binding protein | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000182836 | PLCXD3|phosphatidylinositol-specific phospholipase C, X domain containing 3 | −6.53477 | 5 × 10−5 | 0.00026 |
| ENSG00000083782 | EPYC|epiphycan | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000253293 | HOXA10|homeobox A10 | −6.24454 | 5 × 10−5 | 0.00026 |
| ENSG00000086205 | FOLH1|folate hydrolase (prostate-specific membrane antigen) 1///FOLH1|folate hydrolase 1B | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000259527 | LINC00052|long intergenic non-protein coding RNA 52 | −6.17543 | 5 × 10−5 | 0.00026 |
| ENSG00000101441 | CST4|cystatin S | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000006747 | SCIN|scinderin | −5.91229 | 5 × 10−5 | 0.00026 |
| ENSG00000101460 | MAP1LC3A|microtubule-associated protein 1 light chain 3 alpha | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000110492 | MDK|midkine (neurite growth-promoting factor 2) | −5.60036 | 5 × 10−5 | 0.00026 |
| ENSG00000102854 | MSLN|mesothelin | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000140538 | NTRK3|neurotrophic tyrosine kinase, receptor, type 3 | −5.39223 | 5 × 10−5 | 0.00026 |
| ENSG00000104313 | EYA1|EYA transcriptional coactivator and phosphatase 1 | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000102271 | KLHL4|kelch-like family member 4 | −5.2236 | 5 × 10−5 | 0.00026 |
| ENSG00000109846 | CRYAB|crystallin, alpha B | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000105974 | CAV1|caveolin 1, caveolae protein, 22kDa | −5.18256 | 5 × 10−5 | 0.00026 |
| ENSG00000111863 | ADTRP|androgen-dependent TFPI-regulating protein | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000112175 | BMP5|bone morphogenetic protein 5 | −5.08812 | 5 × 10−5 | 0.00026 |
| ENSG00000113805 | CNTN3|contactin 3 (plasmacytoma associated) | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000159618 | ADGRG5|adhesion G protein-coupled receptor G5 | −4.97744 | 5 × 10−5 | 0.00026 |
| ENSG00000115009 | CCL20|chemokine (C-C motif) ligand 20 | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000174498 | IGDCC3|immunoglobulin superfamily, DCC subclass, member 3 | −4.96488 | 5 × 10−5 | 0.00026 |
| ENSG00000125788 | DEFB126|defensin, beta 126 | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000168269 | FOXI1|forkhead box I1 | −4.95038 | 5 × 10−5 | 0.00026 |
| ENSG00000131095 | GFAP|glial fibrillary acidic protein | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000087077 | TRIP6|thyroid hormone receptor interactor 6 | −4.80366 | 5 × 10−5 | 0.00026 |
| ENSG00000139329 | LUM|lumican | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000134193 | REG4|regenerating islet-derived family, member 4 | −4.705 | 5 × 10−5 | 0.00026 |
| ENSG00000140287 | HDC|histidine decarboxylase | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000003436 | TFPI|tissue factor pathway inhibitor (lipoprotein-associated coagulation inhibitor) | −4.62227 | 5 × 10−5 | 0.00026 |
| ENSG00000146233 | CYP39A1|cytochrome P450, family 39, subfamily A, polypeptide 1 | 18.6778 | 5 × 10−5 | 0.00026 | ENSG00000064205 | WISP2|WNT1 inducible signaling pathway protein 2 | −4.57396 | 5 × 10−5 | 0.00026 |
| KEGG ID | Description | p-value | FDR |
|---|---|---|---|
| 4110 | Cell cycle | 8.73 × 10−8 | 0 |
| 3030 | DNA replication | 1.20 × 10−7 | 0 |
| 4114 | Oocyte meiosis | 6.34 × 10−5 | 0 |
| 4914 | Progesterone-mediated oocyte maturation | 8.36 × 10−5 | 0 |
| 4360 | Axon guidance | 8.76 × 10−5 | 0 |
| 3430 | Mismatch repair | 5.20 × 10−4 | 0.01 |
| 4115 | p53 signaling pathway | 1.11 × 10−3 | 0.01143 |
| 4010 | MAPK signaling pathway | 2.91 × 10−3 | 0.03333 |
| 600 | Sphingolipid metabolism | 3.88 × 10−3 | 0.044 |
| 4145 | Phagosome | 4.78 × 10−3 | 0.05 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lanceta, L.; O'Neill, C.; Lypova, N.; Li, X.; Rouchka, E.; Waigel, S.; Gomez-Gutierrez, J.G.; Chesney, J.; Imbert-Fernandez, Y. Transcriptomic Profiling Identifies Differentially Expressed Genes in Palbociclib-Resistant ER+ MCF7 Breast Cancer Cells. Genes 2020, 11, 467. https://doi.org/10.3390/genes11040467
Lanceta L, O'Neill C, Lypova N, Li X, Rouchka E, Waigel S, Gomez-Gutierrez JG, Chesney J, Imbert-Fernandez Y. Transcriptomic Profiling Identifies Differentially Expressed Genes in Palbociclib-Resistant ER+ MCF7 Breast Cancer Cells. Genes. 2020; 11(4):467. https://doi.org/10.3390/genes11040467
Chicago/Turabian StyleLanceta, Lilibeth, Conor O'Neill, Nadiia Lypova, Xiahong Li, Eric Rouchka, Sabine Waigel, Jorge G. Gomez-Gutierrez, Jason Chesney, and Yoannis Imbert-Fernandez. 2020. "Transcriptomic Profiling Identifies Differentially Expressed Genes in Palbociclib-Resistant ER+ MCF7 Breast Cancer Cells" Genes 11, no. 4: 467. https://doi.org/10.3390/genes11040467
APA StyleLanceta, L., O'Neill, C., Lypova, N., Li, X., Rouchka, E., Waigel, S., Gomez-Gutierrez, J. G., Chesney, J., & Imbert-Fernandez, Y. (2020). Transcriptomic Profiling Identifies Differentially Expressed Genes in Palbociclib-Resistant ER+ MCF7 Breast Cancer Cells. Genes, 11(4), 467. https://doi.org/10.3390/genes11040467