Detection of a Novel, and Likely Ancestral, Tn916-Like Element from a Human Saliva Metagenomic Library


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Strains and Culture Conditions
2.2. Creation of a Functional Metagenomic Library
2.3. Screening of the Functional Metagenomic Library
2.4. Plasmid Extraction
2.5. Sequencing and Analysis
3. Results
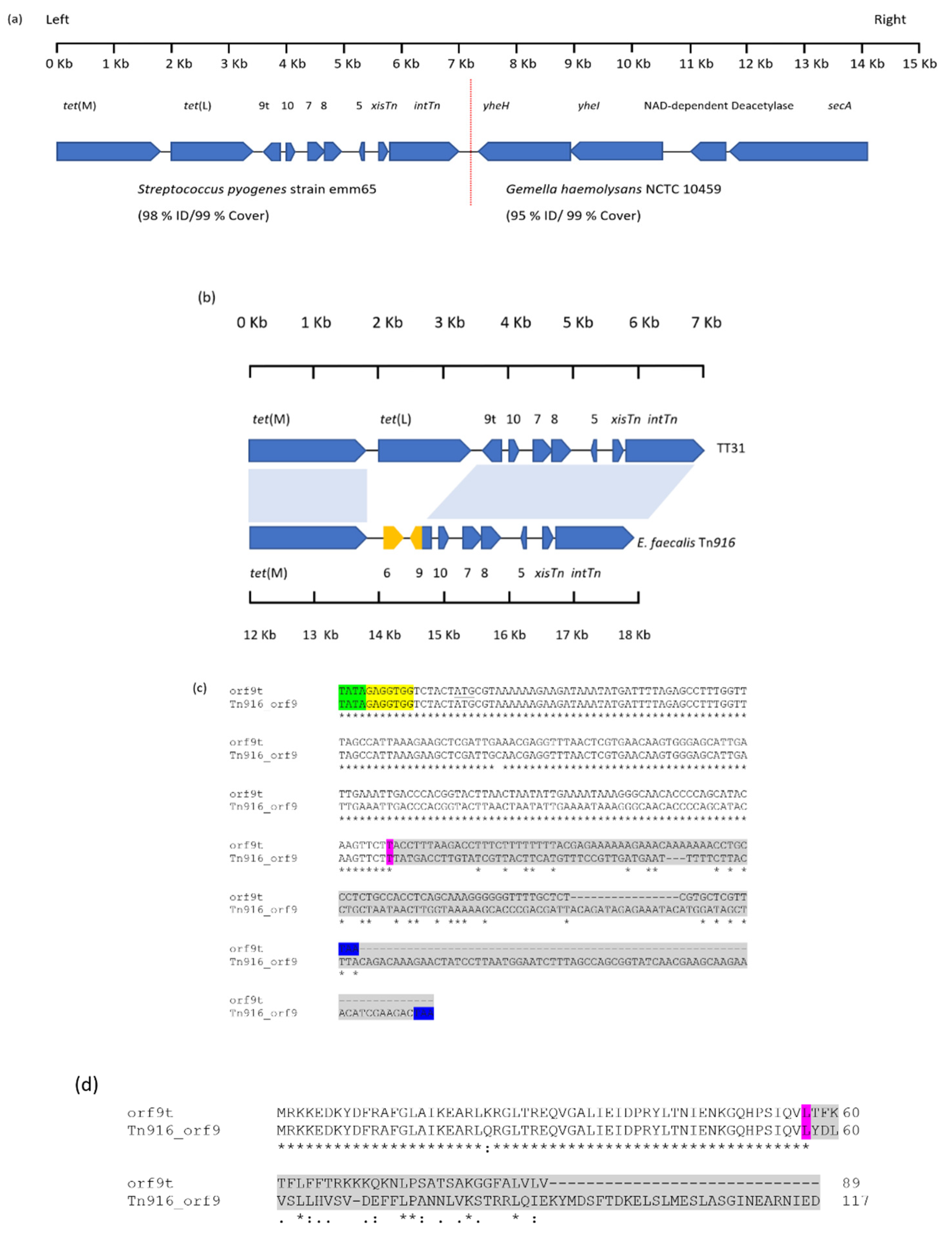
3.1. Sequence Analysis and Gene Annotation of TT31 Insert
3.2. Nucleotide Sequence Homology of the TT31 Insert with Tn916
3.3. TT31 Encodes a Truncated orf9
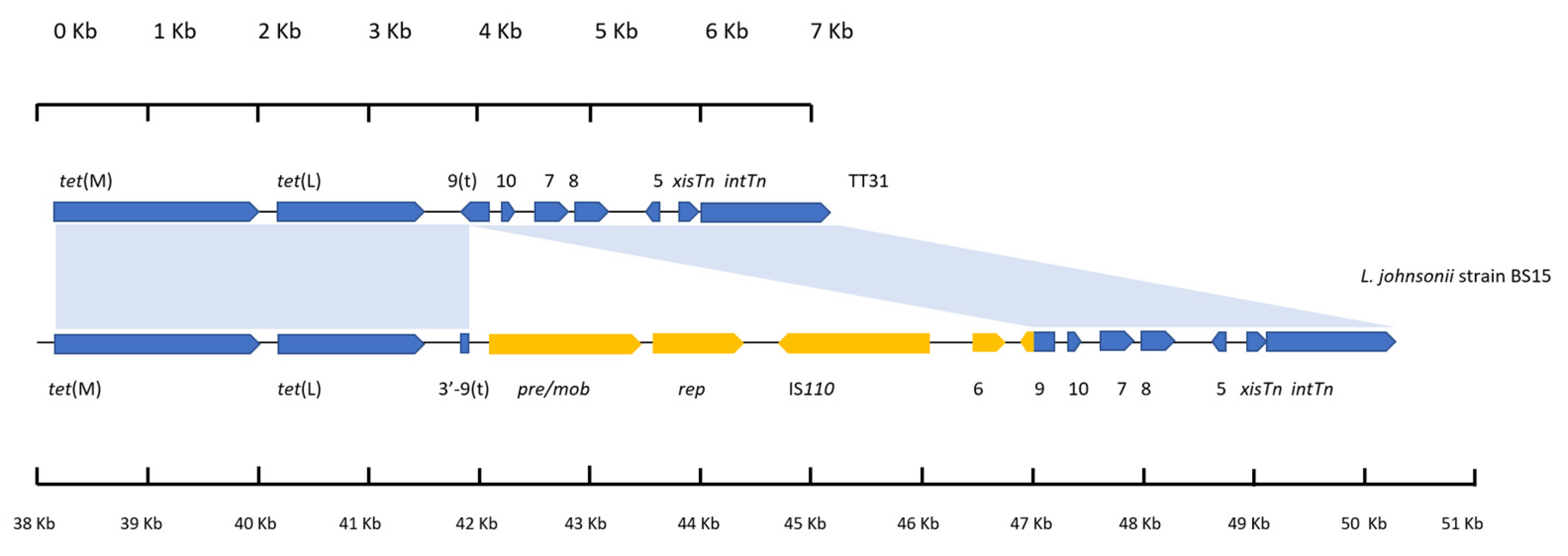
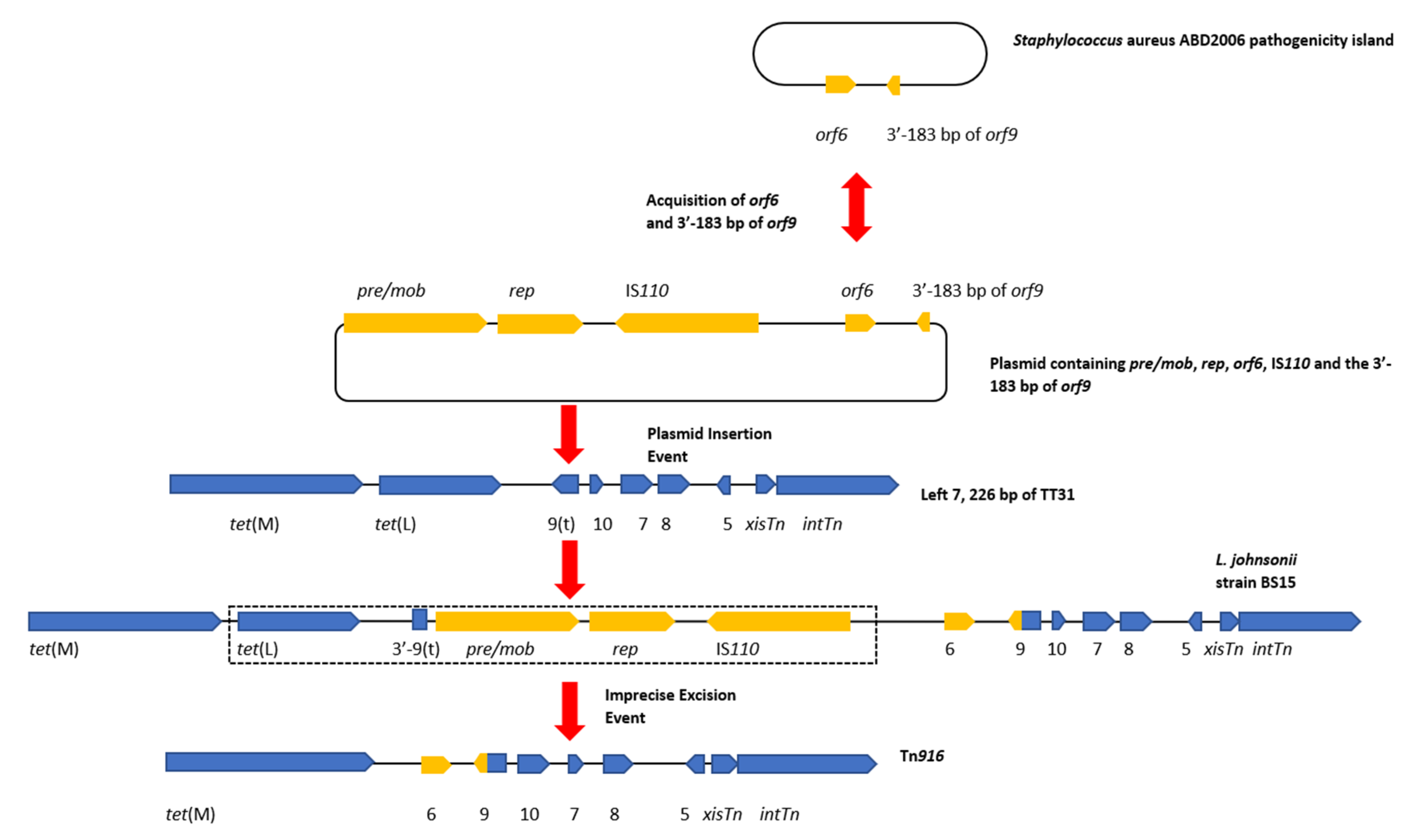
3.4. A Plasmid Insertion Occurred within orf9t in Lactobacillus Johnsonii
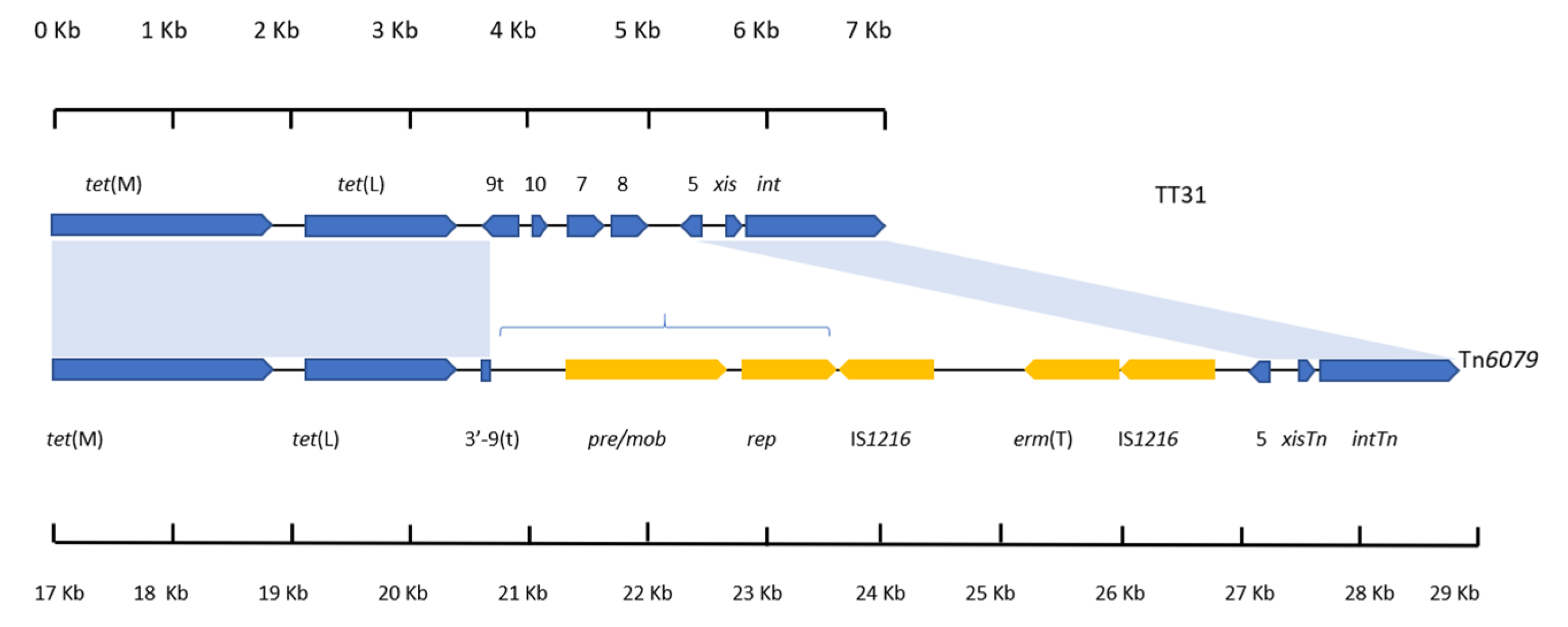
3.5. tet(L) and the 3′-orf9t Sequence Are Present in the TT31 Tn916-Like Element and Tn6079
4. Discussion
4.1. A Partial Tn916-Like Element Was Cloned in TT31
4.2. The Impact of tet(L) Expression on a Tn916-Like Element
4.3. The TT31 Insert Contained a Partial ‘Ancestral’ Tn916-Like Element
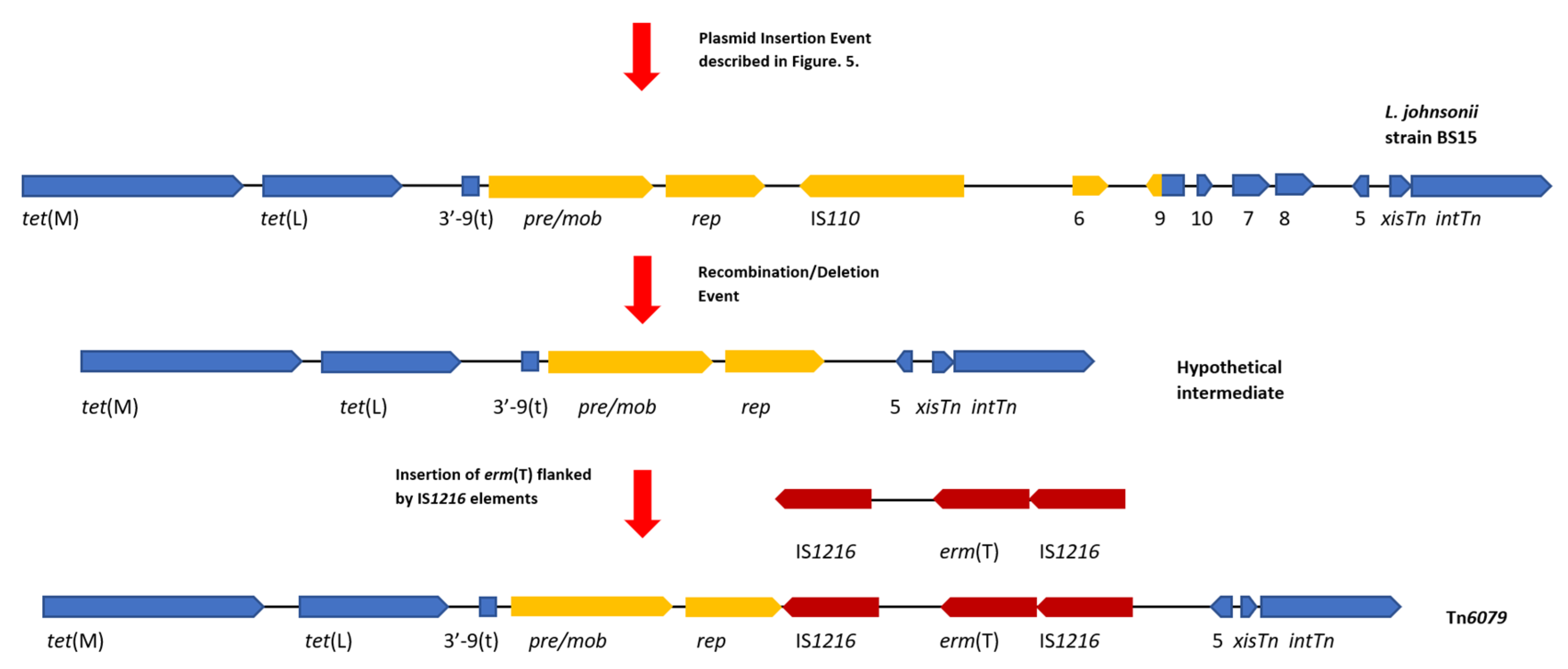
4.4. Tn6079 Descends from the TT31 Tn916-Like Element
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ciric, L.; Mullany, P.; Roberts, A.P. Antibiotic and antiseptic resistance genes are linked on a novel mobile genetic element: Tn6087. J. Antimicrob. Chemother. 2011, 66, 2235–2239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franke, A.; Clewell, D. Evidence for a chromosome-borne resistance transposon (Tn916) in Streptococcus faecalis that is capable of “conjugal” transfer in the absence of a conjugative plasmid. J. Bacteriol. 1981, 145, 494–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, A.; Mullany, P. A modular master on the move: The Tn916 family of mobile genetic elements. Trends Microbiol. 2009, 17, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Wright, L.; Grossman, A. Autonomous Replication of the Conjugative Transposon Tn916. J. Bacteriol. 2016, 198, 3355–3366. [Google Scholar] [CrossRef] [Green Version]
- Lunde, T.; Roberts, A.; Al-Haroni, M. Determination of copy number and circularization ratio of Tn916-Tn1545 family of conjugative transposons in oral streptococci by droplet digital PCR. J. Oral Microbiol. 2019, 11, 1552060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochetti, I.; Tili, E.; Mingoia, M.; Varaldo, P.; Montanari, M. erm(B)-carrying elements in tetracycline-resistant pneumococci and correspondence between Tn1545 and Tn6003. Antimicrob. Agents Chemother. 2008, 52, 1285–1290. [Google Scholar] [CrossRef] [Green Version]
- Soge, O.; Beck, N.; White, T.; No, D.; Roberts, M.C. A novel transposon, Tn6009, composed of a Tn916 element linked with a Staphylococcus aureus mer operon. J. Antimicrob. Chemother. 2008, 62, 674–680. [Google Scholar] [CrossRef]
- Flannagan, S.; Zitzow, L.; Su, Y.; Clewell, D. Nucleotide sequence of the 18-kb conjugative transposon Tn916 from Enterococcus faecalis. Plasmid 1994, 32, 350–354. [Google Scholar] [CrossRef] [Green Version]
- Brenciani, A.; Bacciaglia, A.; Vecchi, M.; Vitali, L.; Varaldo, P.; Giovanetti, E. Genetic elements carrying erm(B) in Streptococcus pyogenes and association with tet(M) tetracycline resistance gene. Antimicrob. Agents Chemother. 2007, 51, 1209–1216. [Google Scholar] [CrossRef] [Green Version]
- De Vries, L.; Vallès, Y.; Agersø, Y.; Vaishampayan, P.; García-Montaner, A.; Kuehl, J.; Christensen, H.; Barlow, M.; Francino, M. The gut as reservoir of antibiotic resistance: Microbial diversity of tetracycline resistance in mother and infant. PLoS ONE 2011, 6, e21644. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.; Grossman, A. Integrative and Conjugative Elements (ICEs): What They Do and How They Work. Annu. Rev. Genet. 2015, 49, 577–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDougal, L.; Tenover, F.; Lee, L.; Rasheed, J.; Patterson, J.; Jorgensen, J.; LeBlanc, D. Detection of Tn917-like sequences within a Tn916-like conjugative transposon (Tn3872) in erythromycin-resistant isolates of Streptococcus pneumoniae. Antimicrob. Agents Chemother. 1998, 42, 2312–2318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leon-Sampedro, R.; Fernandez-de-Bobadilla, M.; San Millan, A.; Baquero, F.; Coque, T. Transfer dynamics of Tn6648, a composite integrative conjugative element generated by tandem accretion of Tn5801 and Tn6647 in Enterococcus faecalis. J. Antimicrob. Chemother. 2019, 74, 2517–2523. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Mullany, P. The large resolvase TndX is required and sufficient for integration and excision of derivatives of the novel conjugative transposon Tn5397. J. Bacteriol. 2000, 182, 6577–6583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brouwer, M.; Mullany, P.; Roberts, A.P. Characterization of the conjugative transposon Tn6000 from Enterococcus casseliflavus 664.1H1 (formerly Enterococcus faecium 664.1H1). FEMS Microbiol. Lett. 2010, 309, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Sebaihia, M.; Wren, B.; Mullany, P.; Fairweather, N.; Minton, N.; Stabler, R.; Thomson, N.; Roberts, A.; Cerdeno-Tarraga, A.; Wang, H.; et al. The multidrug-resistant human pathogen Clostridium difficile has a highly mobile, mosaic genome. Nat. Genet. 2006, 38, 779–786. [Google Scholar] [CrossRef]
- Rice, L.; Carias, L.; Marshall, S.; Rudin, S.; Hutton-Thomas, R. Tn5386, a novel Tn916-like mobile element in Enterococcus faecium D344R that interacts with Tn916 to yield a large genomic deletion. J. Bacteriol. 2005, 19, 6668–6677. [Google Scholar] [CrossRef] [Green Version]
- Wozniak, R.; Fouts, D.; Spagnoletti, M.; Colombo, M.; Ceccarelli, D.; Garriss, G.; Dery, C.; Burrus, V.; Waldor, M. Comparative ICE genomics: Insights into the evolution of the SXT/R391 family of ICEs. PLoS Genet. 2009, 5, e1000786. [Google Scholar] [CrossRef] [Green Version]
- Burrus, V.; Pavlovic, G.; Decaris, B.; Guédon, G. The ICESt1 element of Streptococcus thermophilus belongs to a large family of integrative and conjugative elements that exchange modules and change their specificity of integration. Plasmid 2002, 48, 77–97. [Google Scholar] [CrossRef]
- Pavlovic, G.; Burrus, V.; Gintz, B.; Decaris, B.; Guedon, G. Evolution of genomic islands by deletion and tandem accretion by site-specific recombination: ICESt1-related elements from Streptococcus thermophilus. Microbiology 2004, 150, 759–774. [Google Scholar] [CrossRef] [Green Version]
- Ciric, L.; Ellatif, M.; Sharma, P.; Patel, R.; Song, X.; Mullany, P.; Roberts, A. Tn916-like elements from human, oral, commensal streptococci possess a variety of antibiotic and antiseptic resistance genes. Int. J. Antimicrob. Agents 2012, 39, 360–361. [Google Scholar] [CrossRef] [PubMed]
- Santoro, F.; Vianna, M.; Roberts, A. Variation on a theme; an overview of the Tn916/Tn1545 family of mobile genetic elements in the oral and nasopharyngeal streptococci. Front. Microbiol. 2014, 5, 535. [Google Scholar] [CrossRef] [PubMed]
- Wade, W.; Thompson, H.; Rybalka, A.; Vartoukian, S. Uncultured Members of the Oral Microbiome. J. Calif. Dent. Assoc. 2016, 44, 447–456. [Google Scholar] [PubMed]
- Reynolds, L.; Roberts, A.; Anjum, M. Efflux in the Oral Metagenome: The Discovery of a Novel Tetracycline and Tigecycline ABC Transporter. Front. Microbiol. 2017, 7, 1923. [Google Scholar] [CrossRef] [Green Version]
- Tansirichaiya, S.; Reynolds, L.J.; Cristarella, G.; Wong, L.C.; Rosendahl, K.; Roberts, A.P. Reduced Susceptibility to Antiseptics Is Conferred by Heterologous Housekeeping Genes. Microb. Drug Resist. 2018, 24, 105–112. [Google Scholar] [CrossRef]
- Roberts, A.; Mullany, P. Tn916-like genetic elements: A diverse group of modular mobile elements conferring antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 856–871. [Google Scholar] [CrossRef] [Green Version]
- Zolezzi, P.; Cepero, P.; Ruiz, J.; Laplana, L.; Calvo, C.; Gómez-Lus, R. Molecular epidemiology of macrolide and tetracycline resistances in commensal Gemella sp. isolates. Antimicrob. Agents Chemother. 2007, 51, 1487–1490. [Google Scholar] [CrossRef] [Green Version]
- Cerdá Zolezzi, P.; Laplana, L.; Calvo, C.; Cepero, P.; Erazo, M.; Gómez-Lus, R. Molecular basis of resistance to macrolides and other antibiotics in commensal viridans group streptococci and Gemella spp. and transfer of resistance genes to Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2004, 48, 3462–3467. [Google Scholar] [CrossRef] [Green Version]
- Olsvik, B.; Olsen, I.; Tenover, F. Detection of tet(M) and tet(O) using the polymerase chain reaction in bacteria isolated from patients with periodontal disease. Oral Microbiol. Immunol. 1995, 10, 87–92. [Google Scholar] [CrossRef]
- Su, Y.; He, P.; Clewell, D. Characterization of the tet(M) determinant of Tn916: Evidence for regulation by transcription attenuation. Antimicrob. Agents Chemother. 1992, 36, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Celli, J.; Trieu-Cuot, P. Circularization of Tn916 is required for expression of the transposon-encoded transfer functions: Characterization of long tetracycline-inducible transcripts reading through the attachment site. Mol. Microbiol. 1998, 28, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Marra, D.; Scott, J. Regulation of excision of the conjugative transposon Tn916. Mol. Microbiol. 1999, 31, 609–621. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Lekshmi, M.; Parvathi, A.; Ojha, M.; Wenzel, N.; Varela, M. Functional and Structural Roles of the Major Facilitator Superfamily Bacterial Multidrug Efflux Pumps. Microorganisms 2020, 8, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, M. Update on acquired tetracycline resistance genes. FEMS Microbiol. Lett. 2005, 245, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.; Hill, C. Surviving the acid test: Responses of gram-positive bacteria to low pH. Microbiol. Mol. Biol. Rev. 2003, 67, 429–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.; Guffanti, A.; Bechhofer, D.; Krulwich, T. Tet(L) and Tet(K) tetracycline-divalent metal/H+ antiporters: Characterization of multiple catalytic modes and a mutagenesis approach to differences in their efflux substrate and coupling ion preferences. J. Bacteriol. 2002, 184, 4722–4732. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Deikus, G.; Powers, B.; Shelden, V.; Krulwich, T.; Bechhofer, D. Adaptive gene expression in Bacillus subtilis strains deleted for tetL. J. Bacteriol. 2006, 188, 7090–7100. [Google Scholar] [CrossRef] [Green Version]
- Laverde Gomez, J.; Hendrickx, A.; Willems, R.; Top, J.; Sava, I.; Huebner, J.; Witte, W.; Werner, G. Intra- and interspecies genomic transfer of the Enterococcus faecalis pathogenicity island. PLoS ONE 2011, 29, e16720. [Google Scholar] [CrossRef] [Green Version]
- Croucher, N.; Harris, S.; Fraser, C.; Quail, M.; Burton, J.; van der Linden, M.; McGee, L.; von Gottberg, A.; Song, J.; Ko, K.; et al. Rapid pneumococcal evolution in response to clinical interventions. Science 2011, 331, 430–434. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, Y.; Wu, C.; Shen, Z.; Schwarz, S.; Du, X.; Dai, L.; Zhang, W.; Zhang, Q.; Shen, J. First report of the multidrug resistance gene cfr in Enterococcus faecalis of animal origin. Antimicrob. Agents Chemother. 2012, 56, 1650–1654. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Lin, D.; Yan, G.; Ye, X.; Wu, S.; Guo, Y.; Zhu, D.; Hu, F.; Zhang, Y.; Wang, F.; et al. vanM, a new glycopeptide resistance gene cluster found in Enterococcus faecium. Antimicrob. Agents Chemother. 2010, 54, 4643–4647. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reynolds, L.J.; Anjum, M.F.; Roberts, A.P. Detection of a Novel, and Likely Ancestral, Tn916-Like Element from a Human Saliva Metagenomic Library. Genes 2020, 11, 548. https://doi.org/10.3390/genes11050548
Reynolds LJ, Anjum MF, Roberts AP. Detection of a Novel, and Likely Ancestral, Tn916-Like Element from a Human Saliva Metagenomic Library. Genes. 2020; 11(5):548. https://doi.org/10.3390/genes11050548
Chicago/Turabian StyleReynolds, Liam J., Muna F. Anjum, and Adam P. Roberts. 2020. "Detection of a Novel, and Likely Ancestral, Tn916-Like Element from a Human Saliva Metagenomic Library" Genes 11, no. 5: 548. https://doi.org/10.3390/genes11050548
APA StyleReynolds, L. J., Anjum, M. F., & Roberts, A. P. (2020). Detection of a Novel, and Likely Ancestral, Tn916-Like Element from a Human Saliva Metagenomic Library. Genes, 11(5), 548. https://doi.org/10.3390/genes11050548