Characterization of POU2F1 Gene and Its Potential Impact on the Expression of Genes Involved in Fur Color Formation in Rex Rabbit


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Samples
2.2. Melanocyte Culture and RNA Extraction
2.3. Real-Time qPCR
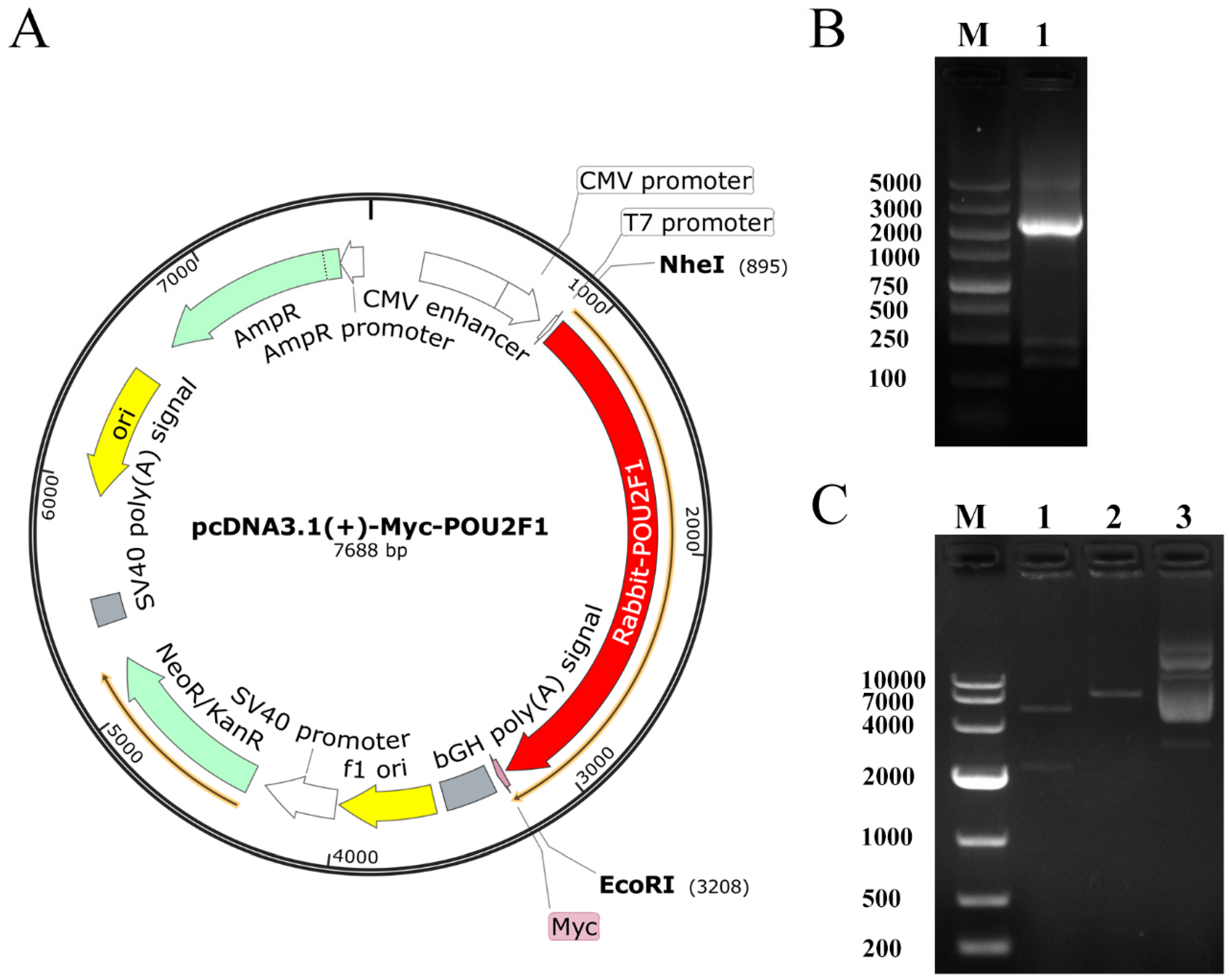
2.4. Construction of pcDNA3.1(+)-Myc-POU2F1 Vector
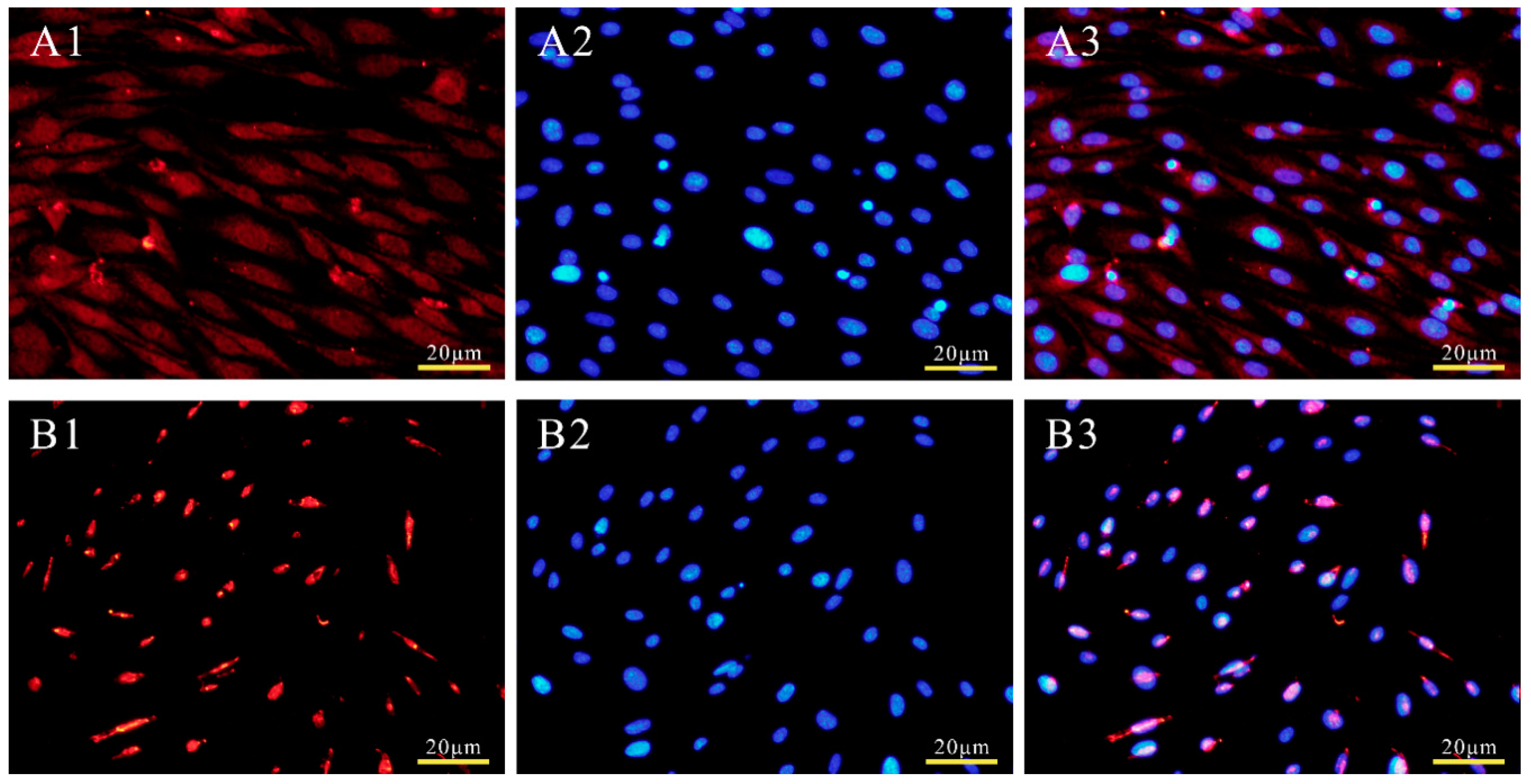
2.5. Subcellular Localization of POU2F1 Protein
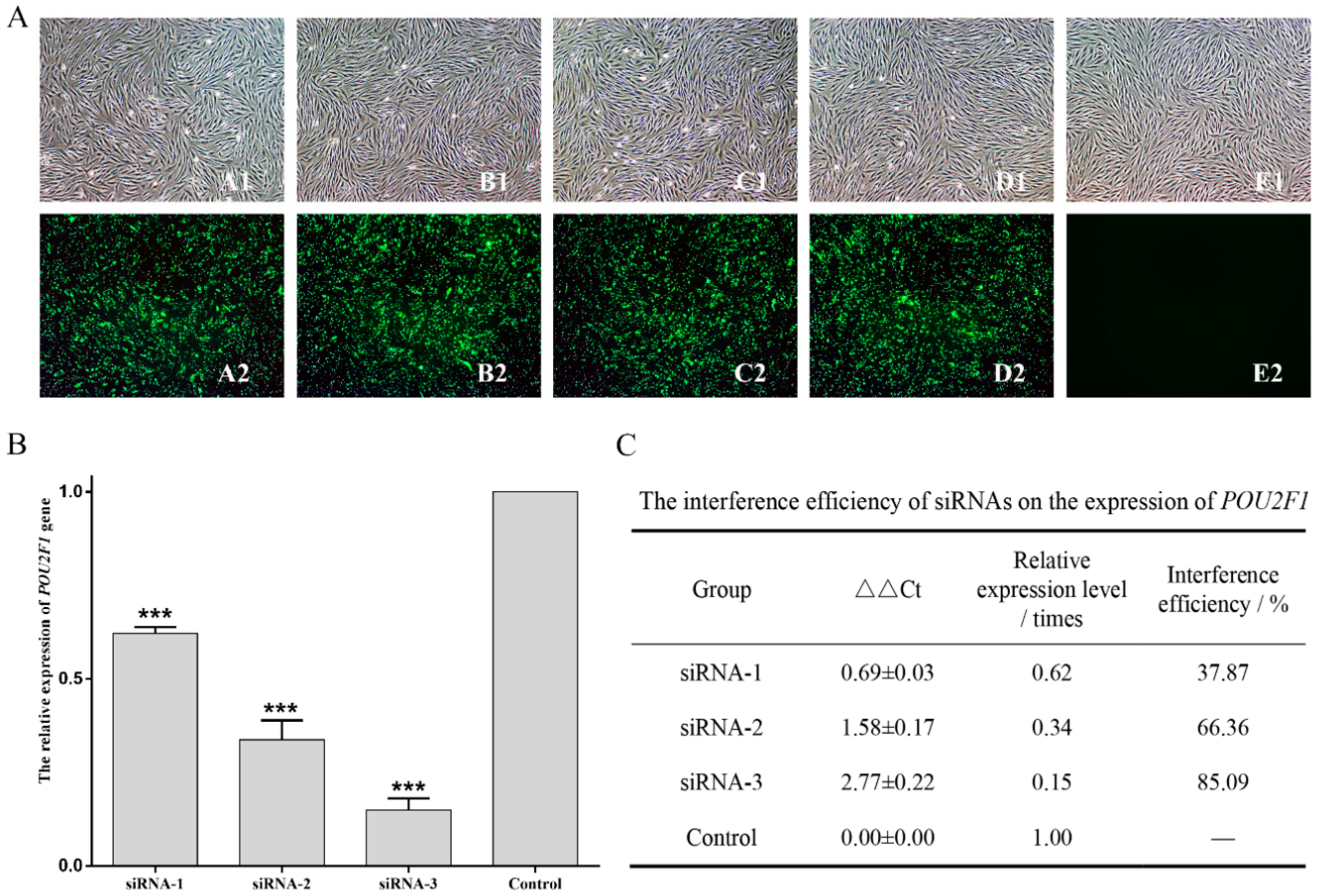
2.6. Inhibition of POU2F1 by siRNA
2.7. Statistical Analysis
3. Results
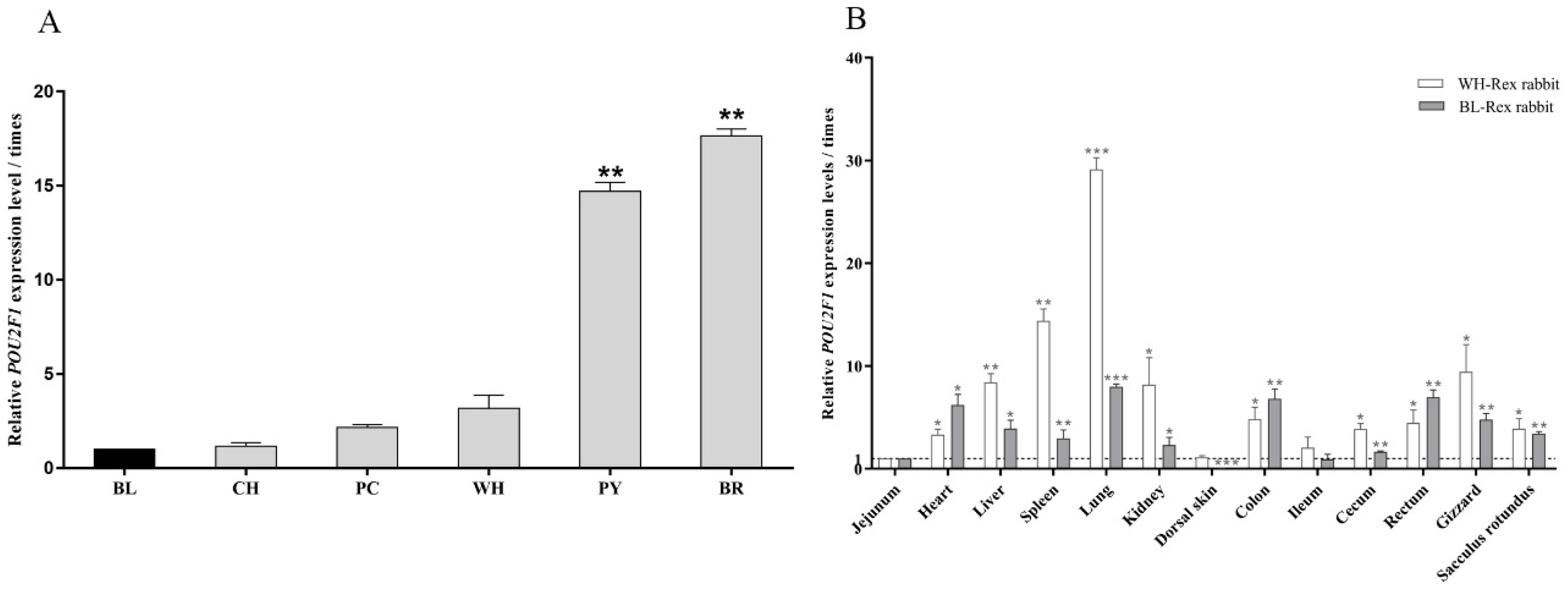
3.1. POU2F1 Gene Expression in Dorsal Skin of Rex Rabbits with Different Fur Colors
3.2. The Tissue Distribution of POU2F1 Gene Expression in WH- and BL-Rex Rabbits
3.3. Subcellular Localization of POU2F1 Protein
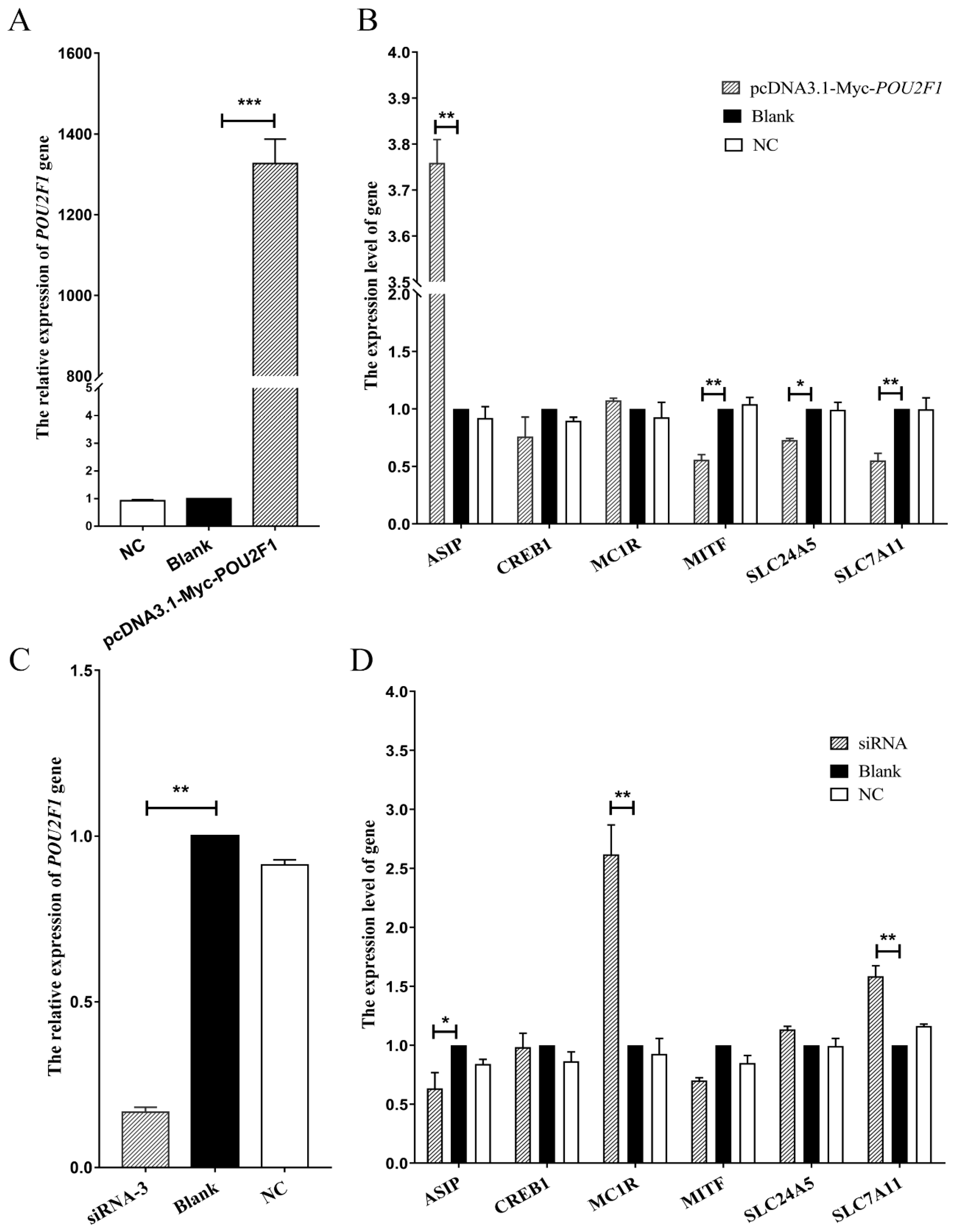
3.4. The Effects of POU2F1 Gene on Other Pigmentation-Related Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tang, J.; Zhang, Y. Color Rabbit Hair Property and Market Foreground. Adv. Mater. Res. 2011, 332, 35–40. [Google Scholar] [CrossRef]
- Kulikov, A.V.; Bazhenova, E.Y.; Kulikova, E.A.; Fursenko, D.V.; Trapezova, L.I.; Terenina, E.E.; Mormede, P.; Popova, N.K.; Trapezov, O.V. Interplay between aggression, brain monoamines and fur color mutation in the American mink. Genes Brain Behav. 2016, 15, 733–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieslak, M.; Reissmann, M.; Hofreiter, M.; Ludwig, A. Colours of domestication. Biol. Rev. 2011, 86, 885–899. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.-S. An Updated Review of Tyrosinase Inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sturm, R.A. Molecular genetics of human pigmentation diversity. Hum. Mol. Genet. 2009, 18, R9–R17. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kanai, Y.; Chairoungdua, A.; Cha, S.H.; Matsuo, H.; Kim, D.K.; Inatomi, J.; Sawa, H.; Ida, Y.; Endou, H. Human cystine/glutamate transporter: cDNA cloning and upregulation by oxidative stress in glioma cells. Biochim. Biophys. Acta (BBA) Biomembr. 2001, 1512, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Chintala, S.; Li, W.; Lamoreux, M.L.; Ito, S.; Wakamatsu, K.; Sviderskaya, E.V.; Bennett, D.C.; Park, Y.-M.; Gahl, W.A.; Huizing, M.; et al. Slc7a11 gene controls production of pheomelanin pigment and proliferation of cultured cells. Proc. Natl. Acad. Sci. USA 2005, 102, 10964–10969. [Google Scholar] [CrossRef] [Green Version]
- Li, H.-T.; He, X.; Zhou, Z.; Zhao, S.-H.; Zhang, W.-X.; Liu, G.; Zhao, Z.-S.; Jia, B. Expression levels of Slc7a11 in the skin of Kazakh sheep with different coat colors. Hereditas (Beijing) 2012, 34, 1314–1319. [Google Scholar] [CrossRef]
- Luo, H.; Hou, J.N.; Li, Y.-M.; Yan, S.Q. Cloning and Sequence of MC1R Gene 5′-Flanking Sequence in Rabbit. China Anim. Husb. Vet. Med. 2011, 38, 89–92. [Google Scholar]
- Yang, G.-L.; Fu, D.-L.; Lang, X.; Wang, Y.-T.; Cheng, S.-R.; Fang, S.-L.; Luo, Y.-Z. Mutations in MC1R Gene Determine Black Coat Color Phenotype in Chinese Sheep. Sci. World J. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.C.; Heux, P.; Santos, C.; Arulvasan, W.; Solanky, N.; Carey, M.E.; Gerrelli, D.; Kinsler, V.A.; Etchevers, H.C. Widespread dynamic and pleiotropic expression of the melanocortin-1-receptor (MC1R) system is conserved across chick, mouse and human embryonic development. Birth Defects Res. 2018, 110, 443–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flesher, J.L.; Paterson-Coleman, E.K.; Vasudeva, P.; Ruiz-Vega, R.; Marshall, M.; Pearlman, E.; MacGregor, G.R.; Neumann, J.; Ganesan, A.K. Delineating the role of MITF isoforms in pigmentation and tissue homeostasis. Pigment Cell Melanoma Res. 2019, 33, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Fontanesi, L.; Forestier, L.; Allain, D.; Scotti, E.; Beretti, F.; Deretz-Picoulet, S.; Pecchioli, E.; Vernesi, C.; Robinson, T.; Malaney, J.L.; et al. Characterization of the rabbit agouti signaling protein (ASIP) gene: Transcripts and phylogenetic analyses and identification of the causative mutation of the nonagouti black coat colour. Genomics 2010, 95, 166–175. [Google Scholar] [CrossRef]
- Chen, Y.; Hu, S.; Mu, L.; Zhao, B.; Wang, M.; Yang, N.; Bao, G.; Zhu, C.; Wu, X. Slc7a11 Modulated by POU2F1 is Involved in Pigmentation in Rabbit. Int. J. Mol. Sci. 2019, 20, 2493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, M.C.Y.; Lam, V.K.L.; Tam, C.H.T.; Chan, A.W.; So, W.-Y.; Ma, R.C.; Zee, B.; Waye, M.M.Y.; Mak, W.W.; Hu, C.; et al. Association of the POU class 2 homeobox 1 gene (POU2F1) with susceptibility to Type 2 diabetes in Chinese populations. Diabet. Med. 2010, 27, 1443–1449. [Google Scholar] [CrossRef]
- Kang, J.; Shen, Z.; Lim, J.-M.; Handa, H.; Wells, L.; Tantin, D. Regulation of Oct1/Pou2f1 transcription activity by O-GlcNAcylation. FASEB J. 2013, 27, 2807–2817. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Xiang, L.; Dai, W.; Tang, W.; Wang, J.; Zhang, Y.; Wu, X.; Liu, G.; Chen, Y.; Zhu, H.; et al. The POU2F1/miR-4490/USP22 Axis Regulates Cell Proliferation and Metastasis in Gastric Cancer. Available online: https://ssrn.com/abstract=3384868 (accessed on 5 May 2019). [CrossRef]
- Dey, A.; Sen, S.; Maulik, U. Studying POU2F1 to Unveil the Structural Facet for Pan-Cancer Analysis Considering the Functional Annotations and Sequence-Structure Space Paradigm. bioRxiv 2019, 806489. [Google Scholar]
- Zhang, R.; Lu, H.; Lyu, Y.-Y.; Yang, X.-M.; Zhu, L.-Y.; Yang, G.-D.; Jiang, P.-C.; Re, Y.; Song, W.-W.; Wang, J.-H. E6/E7-P53-POU2F1-CTHRC1 axis promotes cervical cancer metastasis and activates Wnt/PCP pathway. Sci. Rep. 2017, 7, 44744. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Arreguín, K.; Bensard, C.; Schell, J.C.; Swanson, E.; Chen, X.; Rutter, J.; Tantin, D. Oct1/Pou2f1 is selectively required for colon regeneration and regulates colon malignancy. PLoS Genet. 2019, 15, e1007687. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Huang, H.; Chen, M.; Huang, J.; Wu, Q.; Yan, G.-R.; Chen, D. POU2F1 over-expression correlates with poor prognoses and promotes cell growth and epithelial-to-mesenchymal transition in hepatocellular carcinoma. Oncotarget 2017, 8, 44082–44095. [Google Scholar] [CrossRef]
- Hong, Y.Z.; Guan, Y.C.; Shi, P.W.; Yu, C.; Jian, P.H. POU2F1 promotes growth and metastasis of hepatocellular carcinoma through the FAT1 signaling pathway. Am. J. Cancer Res. 2017, 7, 1665–1679. [Google Scholar]
- Chen, Y.; Hu, S.; Wang, M.; Zhao, B.; Yang, N.; Li, J.; Chen, Q.; Liu, M.; Zhou, J.; Bao, G.; et al. Characterization and Establishment of an Immortalized Rabbit Melanocyte Cell Line Using the SV40 Large T Antigen. Int. J. Mol. Sci. 2019, 20, 4874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svingen, T.; Letting, H.; Hadrup, N.; Hass, U.; Vinggaard, A.M. Selection of reference genes for quantitative RT-PCR (RT-qPCR) analysis of rat tissues under physiological and toxicological conditions. PeerJ 2015, 3, 855. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.-H.; Cao, Y.-M.; Huang, Y.; Shi, Q.-W.; Guo, J.-H.; Fan, Z.-W.; Li, J.-G.; Chen, B.-W.; Wu, B.-Y. Long non-coding RNA TUG1 contributes to tumorigenesis of human osteosarcoma by sponging miR-9-5p and regulating POU2F1 expression. Tumor Biol. 2016, 37, 15031–15041. [Google Scholar] [CrossRef] [PubMed]
- Batool, A.; Jahan, N.; Sun, Y.; Hanif, A.; Xue, H. Genetic association of IDE, POU2F1, PON1, IL1 alpha and IL1 beta with type 2 diabetes in Pakistani population. Mol. Biol. Rep. 2014, 41, 3063–3069. [Google Scholar] [CrossRef]
- Sharpe, D.J.; Orr, K.S.; Moran, M.; White, S.J.; McQuaid, S.; Lappin, T.R.; Thompson, A.; James, J. POU2F1 activity regulates HOXD10 and HOXD11 promoting a proliferative and invasive phenotype in Head and Neck cancer. Oncotarget 2014, 5, 8803–8815. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, Y.; Sun, X.; Mei, C.; Zha, X. MiR-449a promotes liver cancer cell apoptosis by down-regulation of Calpain6 and POU2F1. Oncotarget 2015, 7, 13491–13501. [Google Scholar] [CrossRef] [Green Version]
- Moreno, J.A.; Lambros, M.P.; Ouchi, K.; Kuwahara, Y.; Iehara, T.; Konishi, E.; Hosoi, H. Abstract 3209: The expression of SLC7A11 transporter in lung and pancreatic cancer tissues at different stages of development. Tumor Biol. 2015, 75, 3209. [Google Scholar] [CrossRef]
- Ji, X.; Qian, J.; Rahman, S.M.J.; Siska, P.J.; Zou, Y.; Harris, B.K.; Hoeksema, M.D.; Trenary, I.A.; Heidi, C.; Eisenberg, R.; et al. xCT (SLC7A11)-mediated metabolic reprogramming promotes non-small cell lung cancer progression. Oncogene 2018, 37, 5007–5019. [Google Scholar] [CrossRef]
- Gao, J.; Chen, Y.; Yang, Y.; Liang, J.; Xie, J.; Liu, J.; Li, S.; Zheng, G.; Xie, L.; Zhang, R. The transcription factor Pf-POU3F4 regulates expression of the matrix protein genes Aspein and Prismalin-14 in pearl oyster (Pinctada fucata). FEBS J. 2016, 283, 1962–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, A.; Zang, D.-J.; Zhang, Z.; Liu, X.-Z.; He, X.; Yang, L.; Wang, Y.; Zhou, Z.; Zhang, M.-R.; Dai, L.-L.; et al. Exome Sequencing Identifies SLC24A5 as a Candidate Gene for Nonsyndromic Oculocutaneous Albinism. J. Investig. Dermatol. 2013, 133, 1834–1840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginger, R.S.; Askew, S.E.; Ogborne, R.M.; Wilson, S.; Ferdinando, D.; Dadd, T.; Smith, A.M.; Kazi, S.; Szerencsei, R.T.; Winkfein, R.J.; et al. SLC24A5 Encodes atrans-Golgi Network Protein with Potassium-dependent Sodium-Calcium Exchange Activity That Regulates Human Epidermal Melanogenesis. J. Biol. Chem. 2007, 283, 5486–5495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamason, R.; Mohideen, M.-A.P.; Mest, J.R.; Wong, A.C.; Norton, H.L.; Aros, M.C.; Jurynec, M.; Mao, X.; Humphreville, V.R.; Humbert, J.E.; et al. SLC24A5, a Putative Cation Exchanger, Affects Pigmentation in Zebrafish and Humans. Science 2005, 310, 1782–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goding, C.R. Mitf from neural crest to melanoma: Signal transduction and transcription in the melanocyte lineage. Genes Dev. 2000, 14, 1712–1728. [Google Scholar] [PubMed]
- Bauer, G.L.; Praetorius, C.; Bergsteinsdóttir, K.; Hallsson, J.H.; Gísladóttir, B.K.; Schepsky, A.; Swing, D.A.; O’Sullivan, T.N.; Arnheiter, H.; Bismuth, K.; et al. The Role of MITF Phosphorylation Sites During Coat Color and Eye Development in Mice Analyzed by Bacterial Artificial Chromosome Transgene Rescue. Genetics 2009, 183, 581–594. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, L.C.; Hamer, M.A.; Gunn, D.A.; Deelen, J.; Lall, J.S.; Van Heemst, D.; Uh, H.-W.; Hofman, A.; Uitterlinden, A.G.; Griffiths, C.E.M.; et al. A Genome-Wide Association Study Identifies the Skin Color Genes IRF4, MC1R, ASIP, and BNC2 Influencing Facial Pigmented Spots. J. Investig. Dermatol. 2015, 135, 1735–1742. [Google Scholar] [CrossRef] [Green Version]
- Landi, M.T.; Kanetsky, P.A.; Tsang, S.; Gold, B.; Munroe, D.; Rebbeck, T.; Swoyer, J.; Ter-Minassian, M.; Hedayati, M.; Grossman, L.; et al. MC1R, ASIP, and DNA Repair in Sporadic and Familial Melanoma in a Mediterranean Population. J. Natl. Cancer Inst. 2005, 97, 998–1007. [Google Scholar] [CrossRef] [Green Version]
- Fontanesi, L.; Dall’Olio, S.; Beretti, F.; Portolano, B.; Russo, V. Coat colours in the Massese sheep breed are associated with mutations in the agouti signalling protein (ASIP) and melanocortin 1 receptor (MC1R) genes. Animal 2010, 5, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Yang, M.; Yue, Y.; Guo, T.; Liu, J.; Niu, C.; Yang, B. Analysis of agouti signaling protein (ASIP) gene polymorphisms and association with coat color in Tibetan sheep (Ovis aries). Genet. Mol. Res. 2015, 14, 1200–1209. [Google Scholar] [CrossRef]
- Rieder, S.; Taourit, S.; Mariat, D.; Langlois, B.; Guérin, G. Mutations in the agouti (ASIP), the extension (MC1R), and the brown (TYRP1) loci and their association to coat color phenotypes in horses (Equus caballus). Mamm. Genome 2001, 12, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Kreuzer, S. A Mutation in the Equine SLC24A5 Gene Is Associated With A Dilution Of Black Horses. In Proceedings of the World Congress on Genetics Applied to Livestock Production, Leipzig, Germany, 1–6 August 2010. [Google Scholar]
- Haase, B.; Signer-Hasler, H.; Binns, M.M.; Obexer-Ruff, G.; Hauswirth, R.; Bellone, R.R.; Burger, D.; Rieder, S.; Wade, C.M.; Leeb, T. Accumulating Mutations in Series of Haplotypes at the KIT and MITF Loci Are Major Determinants of White Markings in Franches-Montagnes Horses. PLoS ONE 2013, 8, e75071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, H.; Ren, J.; Ding, N.; Xiao, S.; Huang, L. Genetic variation within coat color genes of MC1R and ASIP in Chinese brownish red Tibetan pigs. Anim. Sci. J. 2010, 81, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Bradley, B.J.; Gerald, M.S.; Widdig, A.; Mundy, N.I. Coat Color Variation and Pigmentation Gene Expression in Rhesus Macaques (Macaca mulatta). J. Mamm. Evol. 2012, 20, 263–270. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | F/R | Sequences of Primers (5′-3′) | Accession No. |
|---|---|---|---|
| MITF | F | AGCTTGCCATGTCCAAACCAG | XM_008260927.1 |
| R | TTCATACTTGGGCACTCGCTCT | ||
| MC1R | F | CCTCGGCACCCCACTTGCAG | NW_003159591.1 |
| R | CAGCACCTCCTTGAGCGTCCTG | ||
| SLC24A5 | F | CTGTCCCTCATGAAGACTACCGTT | XM_002717863.2 |
| R | ACACCACCTATGTTCAACTGGC | ||
| ASIP | F | CTGTGCTTCCTCACTGCCTATAGCC | NM_001122939.1 |
| R | TTCAGCGCCACAATGGAGACCGAA | ||
| CREB1 | F | CCTCCCCAGCACTTCCTACACA | XM_008259050.1 |
| R | TTCAGCTCCTCAATCAGCGTCT | ||
| SLC7A11 | F | TCACCATTGGCTACGTGCT | NC_013683.1 |
| R | GCCACAAAGATCGGAACTGCT | ||
| POU2F1 | F | GCAGCAGCAGCAGATTCAAGAATG | XM_017345713.1 |
| R | GTGTGCCTGTGTTGCCATCTCC | ||
| GAPDH | F | CACCAGGGCTGCTTTTAACTCT | NM_001082253.1 |
| R | CTTCCCGTTCTCAGCCTTGACC | ||
| β-actin | F | CGGCACCAGGGCGTGAT | NM_001101683 |
| R | CGCTTGCTCTGGGCCTCGT | ||
| HPRT1 | F | CGTCGAGGACTTGGAAAGGG | NM_001105671.1 |
| R | TTGAGCACACAGAGGGCTAC |
| siRNAs | S/A | Sequences of Primers (5′-3′) |
|---|---|---|
| siRNA1 | S | GGAAGAGCCCAGUGACCUUTT |
| A | AAGGUCACUGGGCUCUUCCTT | |
| siRNA2 | S | CCUUGAACCUCAGCUUUAATT |
| A | UUAAAGCUGAGGUUCAAGGTT | |
| siRNA3 | S | GCCUCCACCUCCGAGACAUTT |
| A | AUGUCUCGGAGGUGGAGGCTT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, N.; Zhao, B.; Hu, S.; Bao, Z.; Liu, M.; Chen, Y.; Wu, X. Characterization of POU2F1 Gene and Its Potential Impact on the Expression of Genes Involved in Fur Color Formation in Rex Rabbit. Genes 2020, 11, 575. https://doi.org/10.3390/genes11050575
Yang N, Zhao B, Hu S, Bao Z, Liu M, Chen Y, Wu X. Characterization of POU2F1 Gene and Its Potential Impact on the Expression of Genes Involved in Fur Color Formation in Rex Rabbit. Genes. 2020; 11(5):575. https://doi.org/10.3390/genes11050575
Chicago/Turabian StyleYang, Naisu, Bohao Zhao, Shuaishuai Hu, Zhiyuan Bao, Ming Liu, Yang Chen, and Xinsheng Wu. 2020. "Characterization of POU2F1 Gene and Its Potential Impact on the Expression of Genes Involved in Fur Color Formation in Rex Rabbit" Genes 11, no. 5: 575. https://doi.org/10.3390/genes11050575
APA StyleYang, N., Zhao, B., Hu, S., Bao, Z., Liu, M., Chen, Y., & Wu, X. (2020). Characterization of POU2F1 Gene and Its Potential Impact on the Expression of Genes Involved in Fur Color Formation in Rex Rabbit. Genes, 11(5), 575. https://doi.org/10.3390/genes11050575