Paraburkholderia Symbionts Display Variable Infection Patterns That Are Not Predictive of Amoeba Host Outcomes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria and Amoeba Culture Conditions
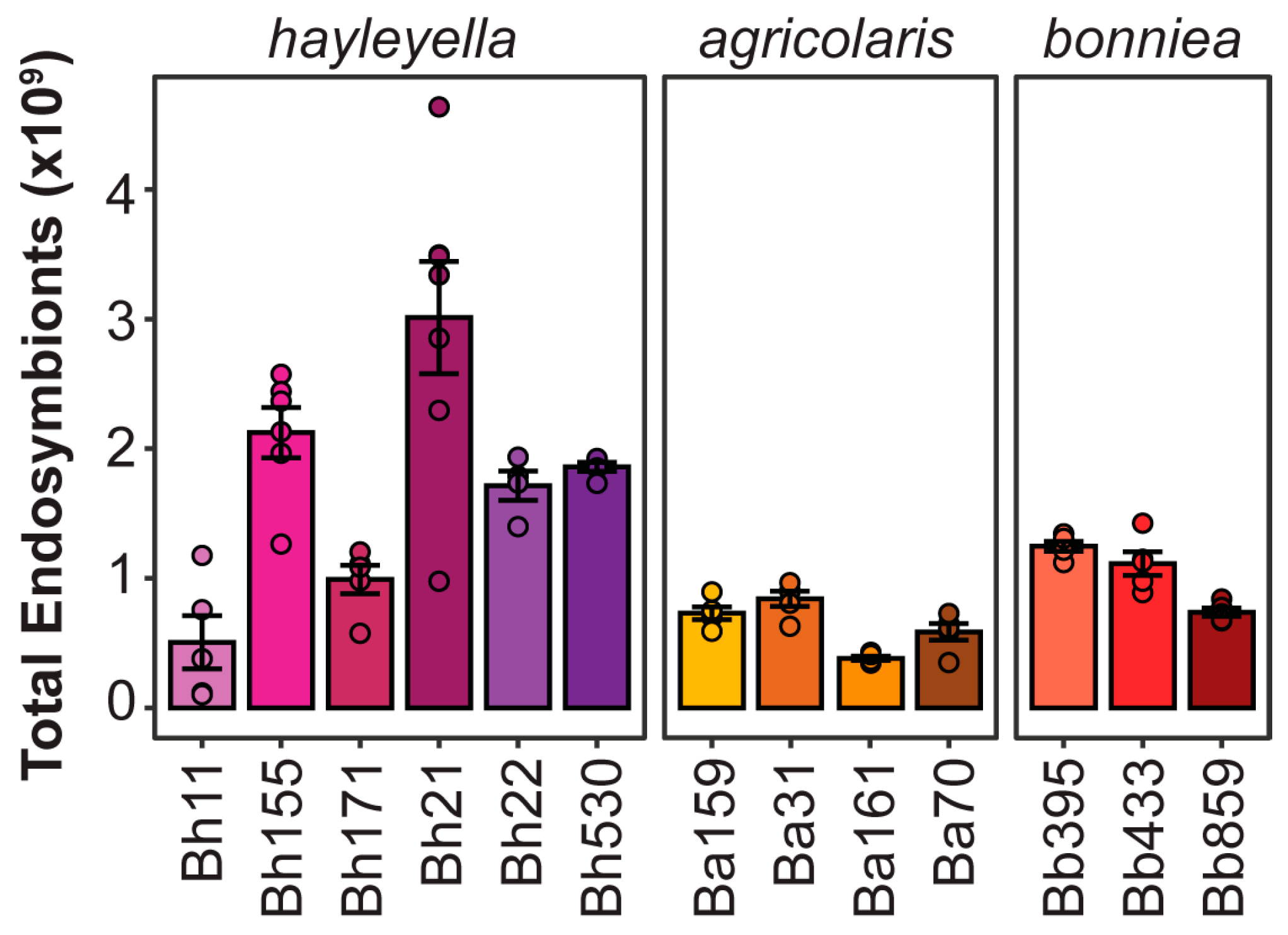
2.2. Confocal Microscopy and Infection Density Assay
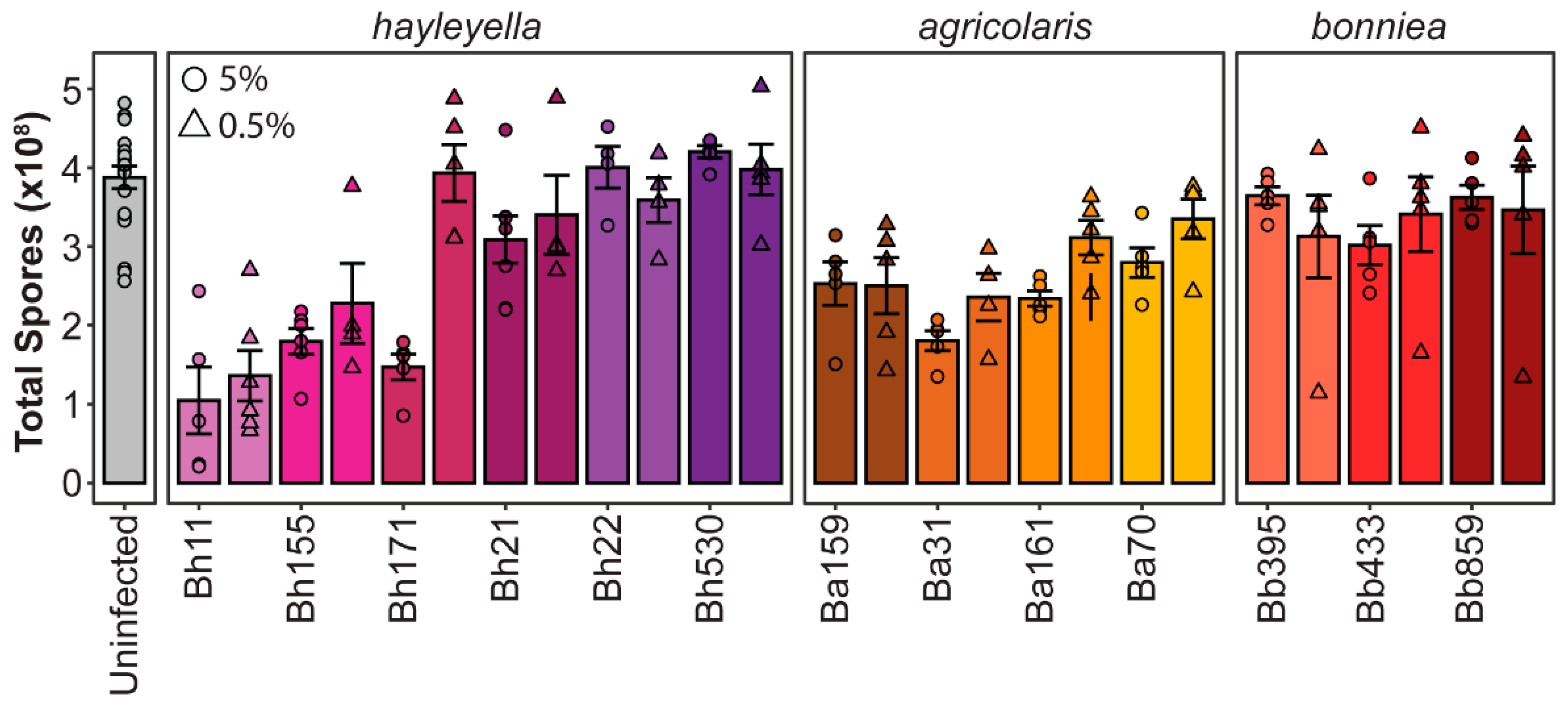
2.3. Total Spore Productivity
2.4. Infection Prevalence
2.5. Infection Stability
2.6. Supernatant Toxicity
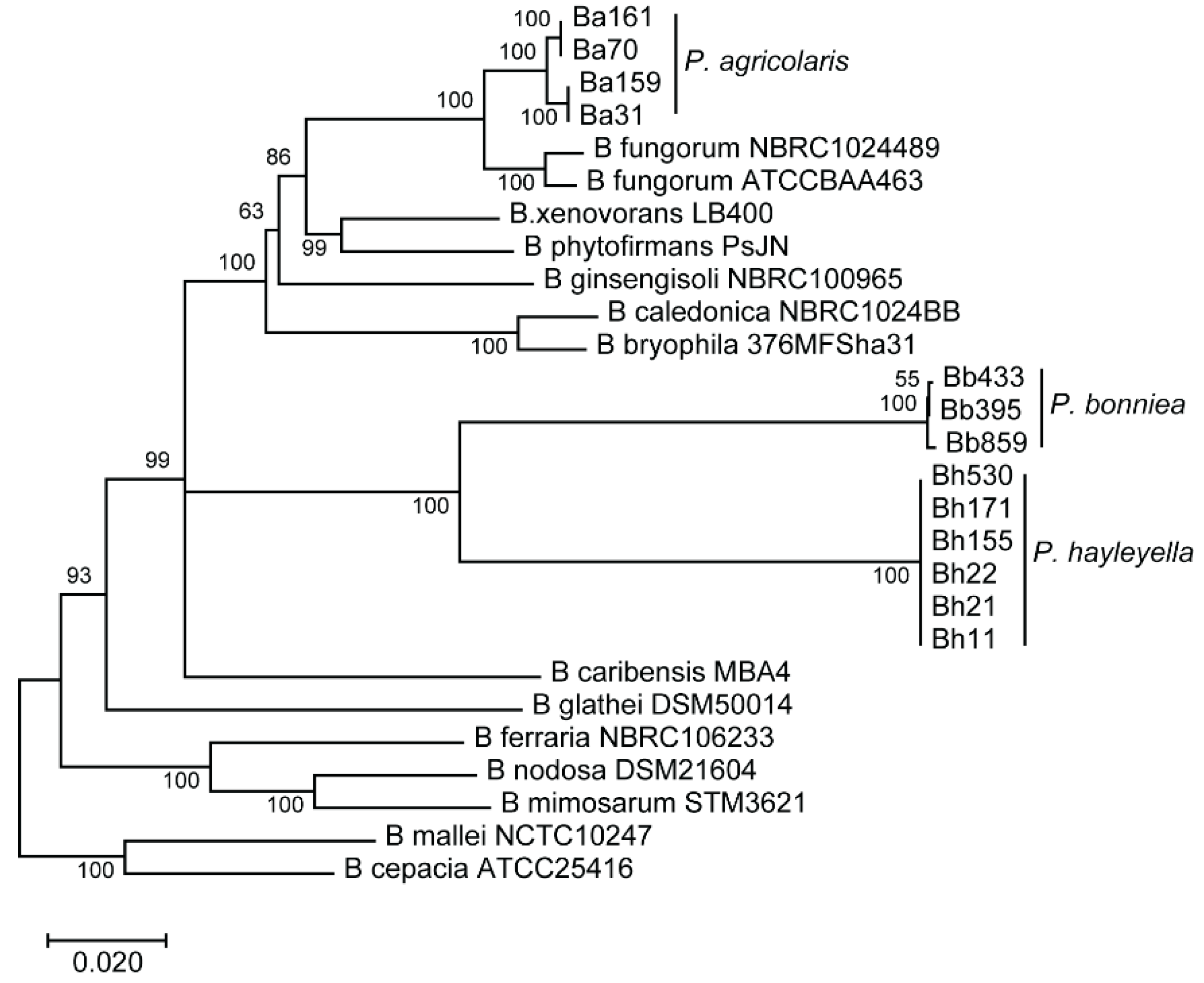
2.7. Phylogenetic Tree Construction
2.8. Statistical Analysis
3. Results
3.1. Different Paraburkholderia Symbionts and Exposure Doses Lead to a Range of Final Infection Patterns
3.2. Final Symbiont Infection Properties do not Strongly Correlate with Host Outcomes
3.3. Secreted Toxins not Detected from P. hayleyella Symbiont Supernatants
3.4. Symbiont Prevalence Plateaus over Multiple Social Cycles, Reaching Similar Levels Regardless of Exposure Dosage
3.5. Symbionts Capable of Achieving High Infection Loads with Minimal Host Damage Should Maximize Symbiont Population Abundance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bozzaro, S.; Bucci, C.; Steinert, M. Phagocytosis and host-pathogen interactions in Dictyostelium with a look at macrophages. Int. Rev. Cell Mol. Biol. 2008, 271, 253–300. [Google Scholar]
- Cardelli, J. Phagocytosis and macropinocytosis in Dictyostelium: Phosphoinositide-based processes, biochemically distinct. Traffic Cph. Den. 2001, 2, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Cosson, P.; Soldati, T. Eat, kill or die: When amoeba meets bacteria. Curr. Opin. Microbiol. 2008, 11, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Nasser, W.; Santhanam, B.; Miranda, E.R.; Parikh, A.; Juneja, K.; Rot, G.; Dinh, C.; Chen, R.; Zupan, B.; Shaulsky, G.; et al. Bacterial discrimination by dictyostelid amoebae reveals the complexity of ancient interspecies interactions. Curr. Biol. 2013, 23, 862–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jousset, A.; Rochat, L.; Scheu, S.; Bonkowski, M.; Keel, C. Predator-Prey Chemical Warfare Determines the Expression of Biocontrol Genes by Rhizosphere-Associated Pseudomonas fluorescens. Appl. Environ. Microbiol. 2010, 76, 5263–5268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denet, E.; Coupat-Goutaland, B.; Nazaret, S.; Pélandakis, M.; Favre-Bonté, S. Diversity of free-living amoebae in soils and their associated human opportunistic bacteria. Parasitol. Res. 2017, 116, 3151–3162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denoncourt, A.M.; Paquet, V.E.; Charette, S.J. Potential role of bacteria packaging by protozoa in the persistence and transmission of pathogenic bacteria. Front. Microbiol. 2014, 5, 240. [Google Scholar] [CrossRef] [Green Version]
- Molmeret, M.; Horn, M.; Wagner, M.; Santic, M.; Abu Kwaik, Y. Amoebae as training grounds for intracellular bacterial pathogens. Appl. Environ. Microbiol. 2005, 71, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Winiecka-Krusnell, J.; Linder, E. Bacterial infections of free-living amoebae. Res. Microbiol. 2001, 152, 613–619. [Google Scholar] [CrossRef]
- Dunn, J.D.; Bosmani, C.; Barisch, C.; Raykov, L.; Lefrançois, L.H.; Cardenal-Muñoz, E.; López-Jiménez, A.T.; Soldati, T. Eat Prey, Live: Dictyostelium discoideum As a Model for Cell-Autonomous Defenses. Front. Immunol. 2018, 8, 1906. [Google Scholar] [CrossRef] [Green Version]
- Skriwan, C.; Fajardo, M.; Hägele, S.; Horn, M.; Wagner, M.; Michel, R.; Krohne, G.; Schleicher, M.; Hacker, J.; Steinert, M. Various bacterial pathogens and symbionts infect the amoeba Dictyostelium discoideum. Int. J. Med. Microbiol. 2002, 291, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Bozzaro, S.; Eichinger, L. The professional phagocyte Dictyostelium discoideum as a model host for bacterial pathogens. Curr. Drug Targets 2011, 12, 942–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, J.M.; Isberg, R.R. Growth of Legionella pneumophila in Dictyostelium discoideum: A novel system for genetic analysis of host–pathogen interactions. Trends Microbiol. 2000, 8, 478–480. [Google Scholar] [CrossRef]
- Steinert, M. Pathogen–host interactions in Dictyostelium, Legionella, Mycobacterium and other pathogens. Semin. Cell Dev. Biol. 2011, 22, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Thewes, S.; Soldati, T.; Eichinger, L. Editorial: Amoebae as Host Models to Study the Interaction with Pathogens. Front. Cell. Infect. Microbiol. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Kessin, R.H. Dictyostelium: Evolution, Cell Biology, and the Development of Multicellularity; Developmental and Cell Biology Series; Cambridge University Press: Cambridge, UK, 2001; ISBN 978-0-521-58364-0. [Google Scholar]
- Chen, G.; Zhuchenko, O.; Kuspa, A. Immune-like phagocyte activity in the social amoeba. Science 2007, 317, 678–681. [Google Scholar] [CrossRef] [Green Version]
- Farinholt, T.; Dinh, C.; Kuspa, A. Microbiome management in the social amoeba Dictyostelium discoideum compared to humans. Int. J. Dev. Biol. 2019, 63, 447–450. [Google Scholar] [CrossRef] [Green Version]
- Peracino, B.; Buracco, S.; Bozzaro, S. The Nramp (Slc11) proteins regulate development, resistance to pathogenic bacteria and iron homeostasis in Dictyostelium discoideum. J. Cell Sci. 2013, 126, 301–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sillo, A.; Bloomfield, G.; Balest, A.; Balbo, A.; Pergolizzi, B.; Peracino, B.; Skelton, J.; Ivens, A.; Bozzaro, S. Genome-wide transcriptional changes induced by phagocytosis or growth on bacteria in Dictyostelium. BMC Genomics 2008, 9, 291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vines, J.H.; King, J.S. The endocytic pathways of Dictyostelium discoideum. Int. J. Dev. Biol. 2019, 63, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Walk, A.; Callahan, J.; Srisawangvong, P.; Leuschner, J.; Samaroo, D.; Cassilly, D.; Snyder, M.L.D. Lipopolysaccharide enhances bactericidal activity in Dictyostelium discoideum cells. Dev. Comp. Immunol. 2011, 35, 850–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brock, D.A.; Douglas, T.E.; Queller, D.C.; Strassmann, J.E. Primitive agriculture in a social amoeba. Nature 2011, 469, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Brock, D.A.; Callison, W.É.; Strassmann, J.E.; Queller, D.C. Sentinel cells, symbiotic bacteria and toxin resistance in the social amoeba Dictyostelium discoideum. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haselkorn, T.S.; DiSalvo, S.; Miller, J.W.; Bashir, U.; Brock, D.A.; Queller, D.C.; Strassmann, J.E. The specificity of Burkholderia symbionts in the social amoeba farming symbiosis: Prevalence, species, genetic and phenotypic diversity. Mol. Ecol. 2019, 28, 847–862. [Google Scholar] [CrossRef] [PubMed]
- Khojandi, N.; Haselkorn, T.S.; Eschbach, M.N.; Naser, R.A.; DiSalvo, S. Intracellular Burkholderia Symbionts induce extracellular secondary infections; driving diverse host outcomes that vary by genotype and environment. ISME J. 2019, 13, 2068–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, L.; Brock, D.A.; Geist, K.S.; Miller, J.W.; Queller, D.C.; Strassmann, J.E.; DiSalvo, S. Symbiont location, host fitness, and possible coadaptation in a symbiosis between social amoebae and bacteria. eLife 2018, 7. [Google Scholar] [CrossRef]
- DiSalvo, S.; Haselkorn, T.S.; Bashir, U.; Jimenez, D.; Brock, D.A.; Queller, D.C.; Strassmann, J.E. Burkholderia bacteria infectiously induce the proto-farming symbiosis of Dictyostelium amoebae and food bacteria. Proc. Natl. Acad. Sci. USA 2015, 112, E5029–E5037. [Google Scholar] [CrossRef] [Green Version]
- Brock, D.A.; Noh, S.; Hubert, A.N.M.; Haselkorn, T.S.; DiSalvo, S.; Suess, M.K.; Bradley, A.S.; Tavakoli-Nezhad, M.; Geist, K.S.; Queller, D.C.; et al. Endosymbiotic adaptations in three new bacterial species associated with Dictyostelium discoideum: Paraburkholderia agricolaris sp. nov., Paraburkholderia hayleyella sp. nov., and Paraburkholderia bonniea sp. nov. PeerJ 2020, 8, e9151. [Google Scholar] [CrossRef]
- Sawana, A.; Adeolu, M.; Gupta, R.S. Molecular signatures and phylogenomic analysis of the genus Burkholderia: Proposal for division of this genus into the emended genus Burkholderia containing pathogenic organisms and a new genus Paraburkholderia gen. nov. harboring environmental species. Front. Genet. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Brock, D.A.; Read, S.; Bozhchenko, A.; Queller, D.C.; Strassmann, J.E. Social amoeba farmers carry defensive symbionts to protect and privatize their crops. Nat. Commun. 2013, 4, 2385. [Google Scholar] [CrossRef]
- Garcia, J.R.; Larsen, T.J.; Queller, D.C.; Strassmann, J.E. Fitness costs and benefits vary for two facultative Burkholderia symbionts of the social amoeba, Dictyostelium discoideum. Ecol. Evol. 2019, 9, 9878–9890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, K.M.; Moran, N.A.; Hunter, M.S. Costs and benefits of a superinfection of facultative symbionts in aphids. Proc. R. Soc. B Biol. Sci. 2006, 273, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Weldon, S.R.; Strand, M.R.; Oliver, K.M. Phage loss and the breakdown of a defensive symbiosis in aphids. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weldon, S.R.; Russell, J.A.; Oliver, K.M. More Is Not Always Better: Coinfections with Defensive Symbionts Generate Highly Variable Outcomes. Appl. Environ. Microbiol. 2019, 86, 02537. [Google Scholar] [CrossRef]
- Rock-Blake, R.; McCormick, M.K.; Brooks, H.E.A.; Jones, C.S.; Whigham, D.F. Symbiont abundance can affect host plant population dynamics. Am. J. Bot. 2017, 104, 72–82. [Google Scholar] [CrossRef]
- Vannette, R.L.; Hunter, M.D. Plant defence theory re-examined: Nonlinear expectations based on the costs and benefits of resource mutualisms: Nonlinear effects of mutualism on defence. J. Ecol. 2011, 99, 66–76. [Google Scholar] [CrossRef]
- Vannette, R.L.; Hunter, M.D. Mycorrhizal abundance affects the expression of plant resistance traits and herbivore performance. J. Ecol. 2013, 101, 1019–1029. [Google Scholar] [CrossRef] [Green Version]
- Anselme, C.; Pérez-Brocal, V.; Vallier, A.; Vincent-Monegat, C.; Charif, D.; Latorre, A.; Moya, A.; Heddi, A. Identification of the Weevil immune genes and their expression in the bacteriome tissue. BMC Biol. 2008, 6, 43. [Google Scholar] [CrossRef] [Green Version]
- Minter, E.J.A.; Lowe, C.D.; Sørensen, M.E.S.; Wood, A.J.; Cameron, D.D.; Brockhurst, M.A. Variation and asymmetry in host-symbiont dependence in a microbial symbiosis. BMC Evol. Biol. 2018, 18, 108. [Google Scholar] [CrossRef]
- Lowe, C.D.; Minter, E.J.; Cameron, D.D.; Brockhurst, M.A. Shining a Light on Exploitative Host Control in a Photosynthetic Endosymbiosis. Curr. Biol. 2016, 26, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Schwartzman, J.A.; Ruby, E.G. A conserved chemical dialog of mutualism: Lessons from squid and vibrio. Microbes Infect. 2016, 18, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sørensen, M.E.S.; Lowe, C.D.; Minter, E.J.A.; Wood, A.J.; Cameron, D.D.; Brockhurst, M.A. The role of exploitation in the establishment of mutualistic microbial symbioses. FEMS Microbiol. Lett. 2019, 366, fnz148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamada, N.; Chen, G.Y.; Inohara, N.; Núñez, G. Control of pathogens and pathobionts by the gut microbiota. Nat. Immunol. 2013, 14, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Barber, B.E.; William, T.; Grigg, M.J.; Parameswaran, U.; Piera, K.A.; Price, R.N.; Yeo, T.W.; Anstey, N.M. Parasite Biomass-Related Inflammation, Endothelial Activation, Microvascular Dysfunction and Disease Severity in Vivax Malaria. PLoS Pathog. 2015, 11, e1004558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Georgiadou, A.; Walther, M.; Nwakanma, D.; Stewart, L.B.; Levin, M.; Otto, T.D.; Conway, D.J.; Coin, L.J.; Cunnington, A.J. Integrated pathogen load and dual transcriptome analysis of systemic host-pathogen interactions in severe malaria. Sci. Transl. Med. 2018, 10, eaar3619. [Google Scholar] [CrossRef] [Green Version]
- Bergkvist, J.; Selander, E.; Pavia, H. Induction of toxin production in dinoflagellates: The grazer makes a difference. Oecologia 2008, 156, 147–154. [Google Scholar] [CrossRef]
- Cosson, P.; Zulianello, L.; Join-Lambert, O.; Faurisson, F.; Gebbie, L.; Benghezal, M.; van Delden, C.; Curty, L.K.; Köhler, T. Pseudomonas aeruginosa Virulence Analyzed in a Dictyostelium discoideum Host System. J. Bacteriol. 2002, 184, 3027–3033. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, N.; Hagedorn, M.; Soldati-Favre, D.; Soldati, T. Prison Break: Pathogens’ Strategies To Egress from Host Cells. Microbiol. Mol. Biol. Rev. 2012, 76, 707–720. [Google Scholar] [CrossRef] [Green Version]
- Meena, N.P.; Kimmel, A.R. Quantification of Live Bacterial Sensing for Chemotaxis and Phagocytosis and of Macropinocytosis. Front. Cell. Infect. Microbiol. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Meena, N.P.; Kimmel, A.R. Chemotactic network responses to live bacteria show independence of phagocytosis from chemoreceptor sensing. eLife 2017, 6, e24627. [Google Scholar] [CrossRef]
- Shu, L.; Zhang, B.; Queller, D.C.; Strassmann, J.E. Burkholderia bacteria use chemotaxis to find social amoeba Dictyostelium discoideum hosts. ISME J. 2018, 12, 1977. [Google Scholar] [CrossRef]
- Lelong, E.; Marchetti, A.; Guého, A.; Lima, W.C.; Sattler, N.; Molmeret, M.; Hagedorn, M.; Soldati, T.; Cosson, P. Role of magnesium and a phagosomal P-type ATPase in intracellular bacterial killing: P-type ATPase, magnesium and intracellular killing. Cell. Microbiol. 2011, 13, 246–258. [Google Scholar] [CrossRef] [Green Version]
- Lamothe, J.; Thyssen, S.; Valvano, M.A. Burkholderia cepacia complex isolates survive intracellularly without replication within acidic vacuoles of Acanthamoeba polyphaga. Cell. Microbiol. 2004, 6, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Helaine, S.; Thompson, J.A.; Watson, K.G.; Liu, M.; Boyle, C.; Holden, D.W. Dynamics of intracellular bacterial replication at the single cell level. Proc. Natl. Acad. Sci. USA 2010, 107, 3746–3751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, A.; Mikkelsen, H.; Jungersen, G. Intracellular Pathogens: Host Immunity and Microbial Persistence Strategies. J. Immunol. Res. 2019, 2019, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Tresse, E.; Giusti, C.; Kosta, A.; Luciani, M.; Golstein, P. Chapter 23 Autophagy and Autophagic Cell Death in Dictyostelium. Methods Enzymol. 2008, 451, 343–358. [Google Scholar]
- Levraud, J.-P.; Adam, M.; Luciani, M.-F.; de Chastellier, C.; Blanton, R.L.; Golstein, P. Dictyostelium cell death. J. Cell Biol. 2003, 160, 1105–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giusti, C.; Luciani, M.-F.; Ravens, S.; Gillet, A.; Golstein, P. Autophagic Cell Death in Dictyostelium Requires the Receptor Histidine Kinase DhkM. Mol. Biol. Cell 2010, 21, 1825–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnoult, D.; Tatischeff, I.; Estaquier, J.; Girard, M.; Sureau, F.; Tissier, J.P.; Grodet, A.; Dellinger, M.; Traincard, F.; Kahn, A.; et al. On the Evolutionary Conservation of the Cell Death Pathway: Mitochondrial Release of an Apoptosis-inducing Factor during Dictyostelium discoideum Cell Death. Mol. Biol. Cell 2001, 12, 3016–3030. [Google Scholar] [CrossRef] [Green Version]
- Calvo-Garrido, J.; Carilla-Latorre, S.; Kubohara, Y.; Santos-Rodrigo, N.; Mesquita, A.; Soldati, T.; Golstein, P.; Escalante, R. Autophagy in Dictyostelium: Genes and pathways, cell death and infection. Autophagy 2010, 6, 686–701. [Google Scholar] [CrossRef] [Green Version]
- Mesquita, A.; Cardenal-Muñoz, E.; Dominguez, E.; Muñoz-Braceras, S.; Nuñez-Corcuera, B.; Phillips, B.A.; Tábara, L.C.; Xiong, Q.; Coria, R.; Eichinger, L.; et al. Autophagy in Dictyostelium: Mechanisms, regulation and disease in a simple biomedical model. Autophagy 2017, 13, 24–40. [Google Scholar] [CrossRef] [Green Version]
- Escoll, P.; Rolando, M.; Buchrieser, C. Modulation of Host Autophagy during Bacterial Infection: Sabotaging Host Munitions for Pathogen Nutrition. Front. Immunol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Newton, I.L.G.; Clark, M.E.; Kent, B.N.; Bordenstein, S.R.; Qu, J.; Richards, S.; Kelkar, Y.D.; Werren, J.H. Comparative Genomics of Two Closely Related Wolbachia with Different Reproductive Effects on Hosts. Genome Biol. Evol. 2016, 8, 1526–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldo, L.; Dunning Hotopp, J.C.; Jolley, K.A.; Bordenstein, S.R.; Biber, S.A.; Choudhury, R.R.; Hayashi, C.; Maiden, M.C.J.; Tettelin, H.; Werren, J.H. Multilocus Sequence Typing System for the Endosymbiont Wolbachia pipientis. Appl. Environ. Microbiol. 2006, 72, 7098–7110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaltenpoth, M.; Flórez, L.V. Versatile and Dynamic Symbioses Between Insects and Burkholderia Bacteria. Annu. Rev. Entomol. 2020, 65, 145–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miller, J.W.; Bocke, C.R.; Tresslar, A.R.; Schniepp, E.M.; DiSalvo, S. Paraburkholderia Symbionts Display Variable Infection Patterns That Are Not Predictive of Amoeba Host Outcomes. Genes 2020, 11, 674. https://doi.org/10.3390/genes11060674
Miller JW, Bocke CR, Tresslar AR, Schniepp EM, DiSalvo S. Paraburkholderia Symbionts Display Variable Infection Patterns That Are Not Predictive of Amoeba Host Outcomes. Genes. 2020; 11(6):674. https://doi.org/10.3390/genes11060674
Chicago/Turabian StyleMiller, Jacob W., Colleen R. Bocke, Andrew R. Tresslar, Emily M. Schniepp, and Susanne DiSalvo. 2020. "Paraburkholderia Symbionts Display Variable Infection Patterns That Are Not Predictive of Amoeba Host Outcomes" Genes 11, no. 6: 674. https://doi.org/10.3390/genes11060674
APA StyleMiller, J. W., Bocke, C. R., Tresslar, A. R., Schniepp, E. M., & DiSalvo, S. (2020). Paraburkholderia Symbionts Display Variable Infection Patterns That Are Not Predictive of Amoeba Host Outcomes. Genes, 11(6), 674. https://doi.org/10.3390/genes11060674