Arabidopsis V-ATPase d2 Subunit Plays a Role in Plant Responses to Oxidative Stress

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
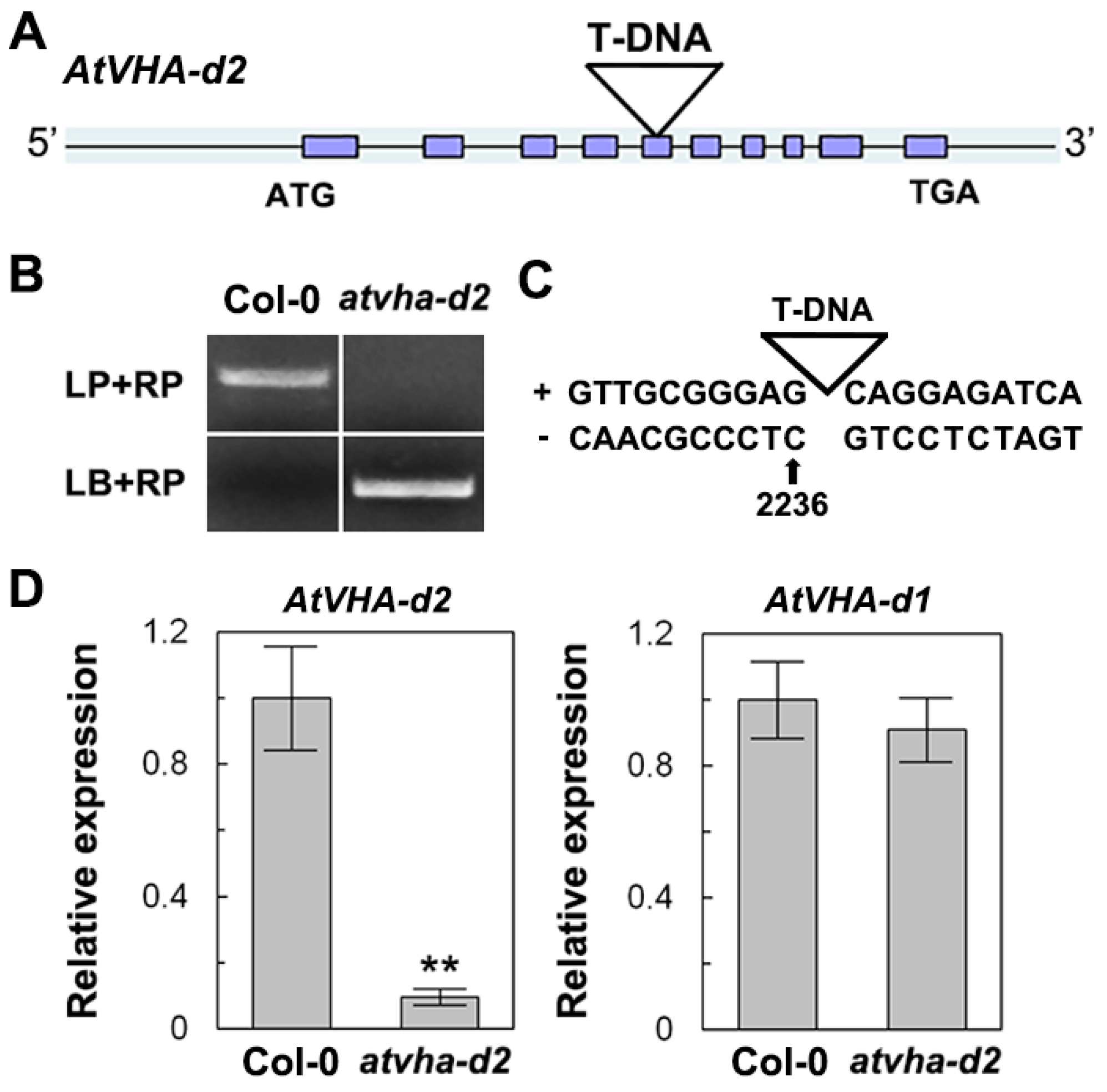
2.2. Vector Construction and Arabidopsis Transformation
2.3. Confocal Laser Scanning Microscopy
2.4. Histochemical β-Glucuronidase Staining
2.5. qPCR Analyses
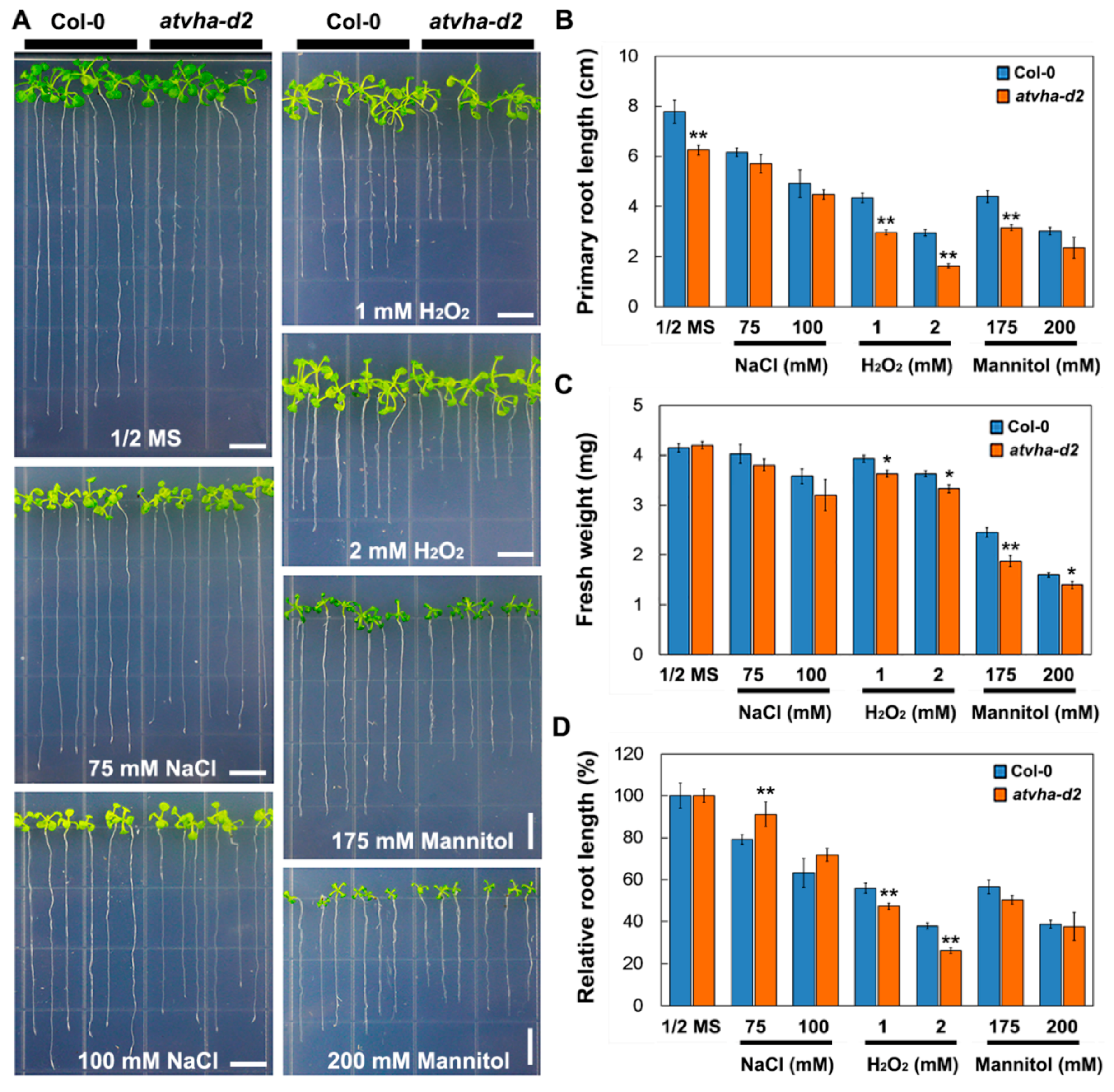
2.6. Stress Tolerant Phenotype
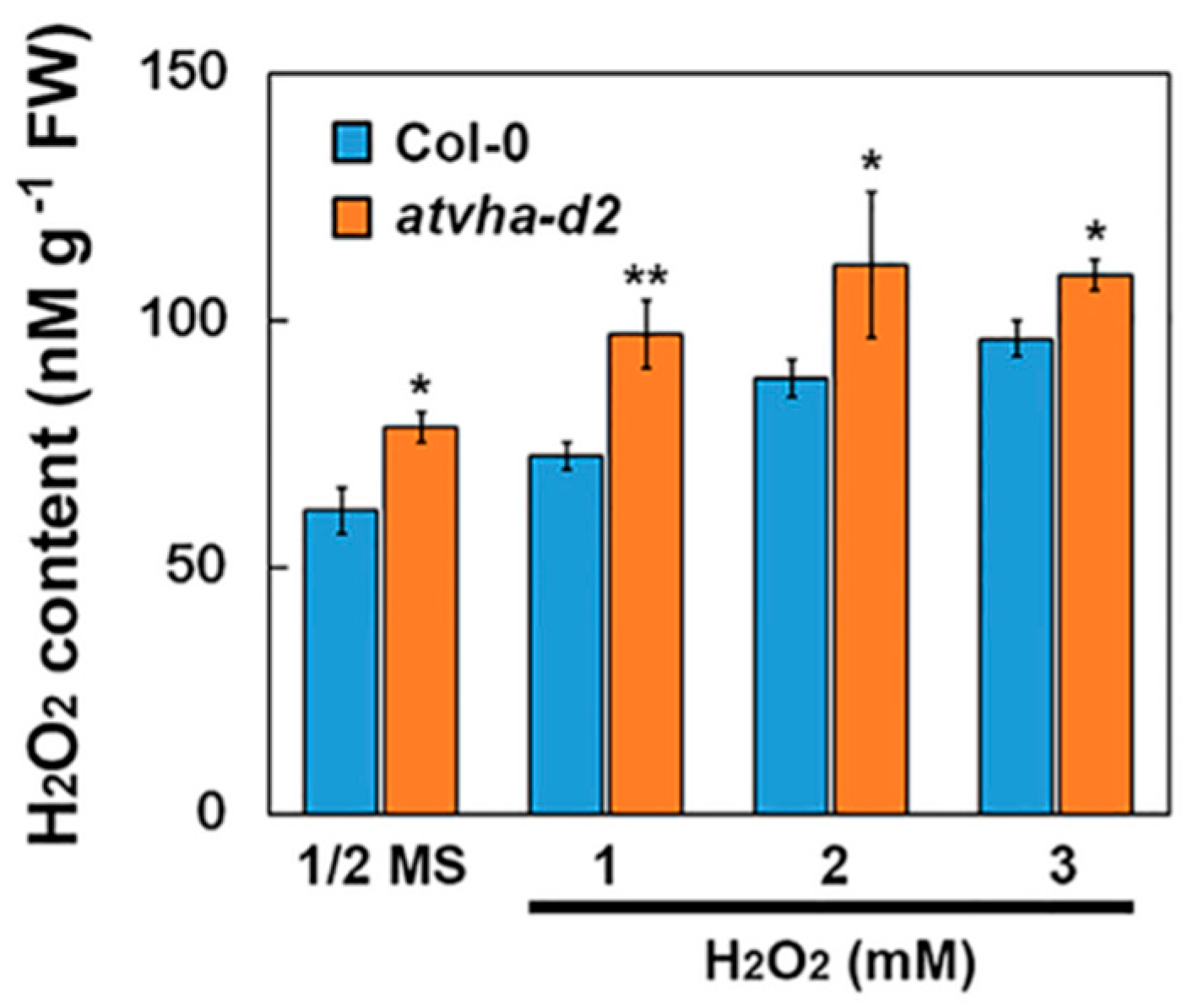
2.7. Hydrogen Peroxide (H2O2) Content Measurement
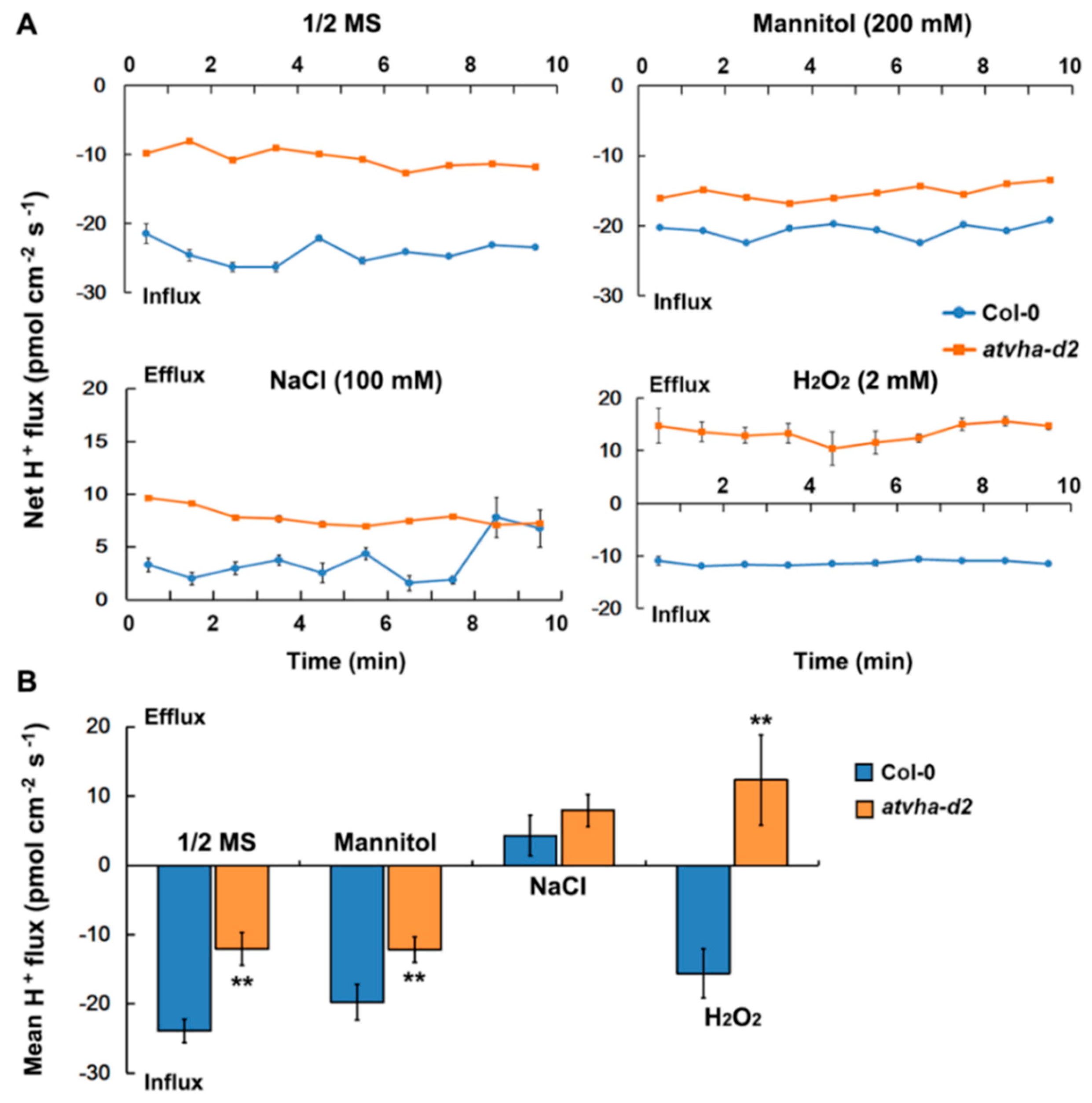
2.8. Net H+ Flux Measurement
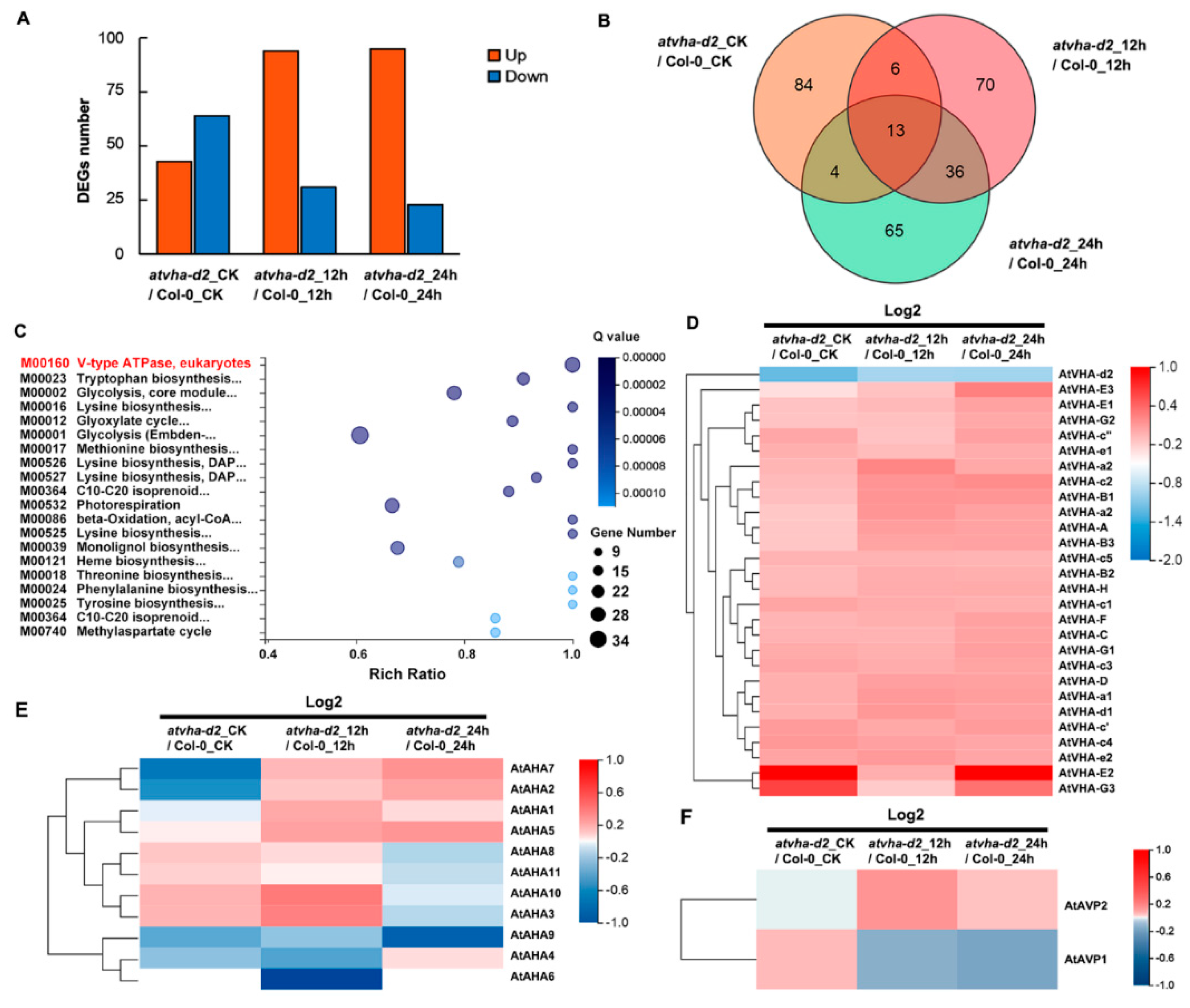
2.9. RNA-seq and DEGs Analysis
2.10. Statistical Analysis
3. Results
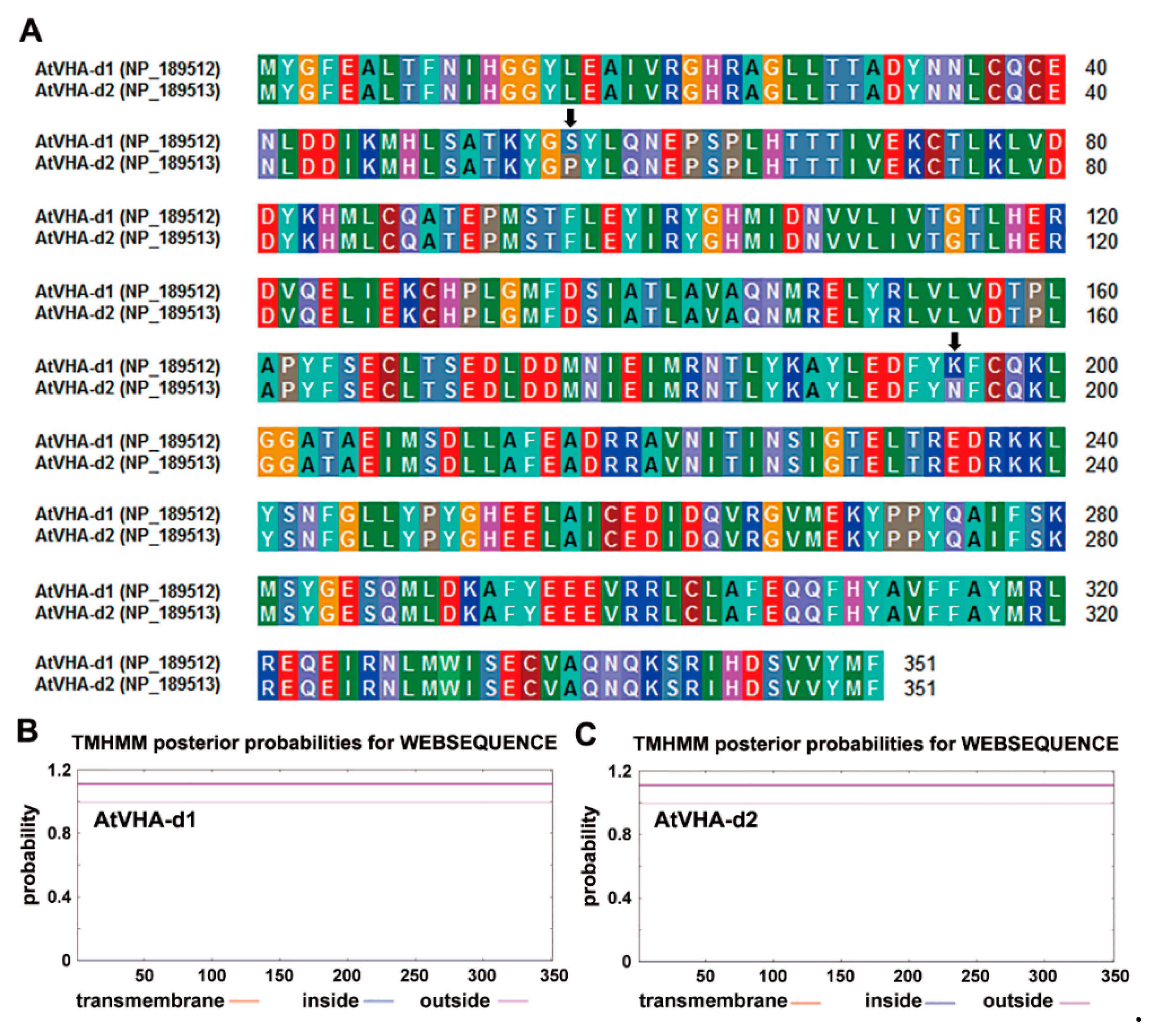
3.1. Tissue Specificity of AtVHA-d Genes Expression
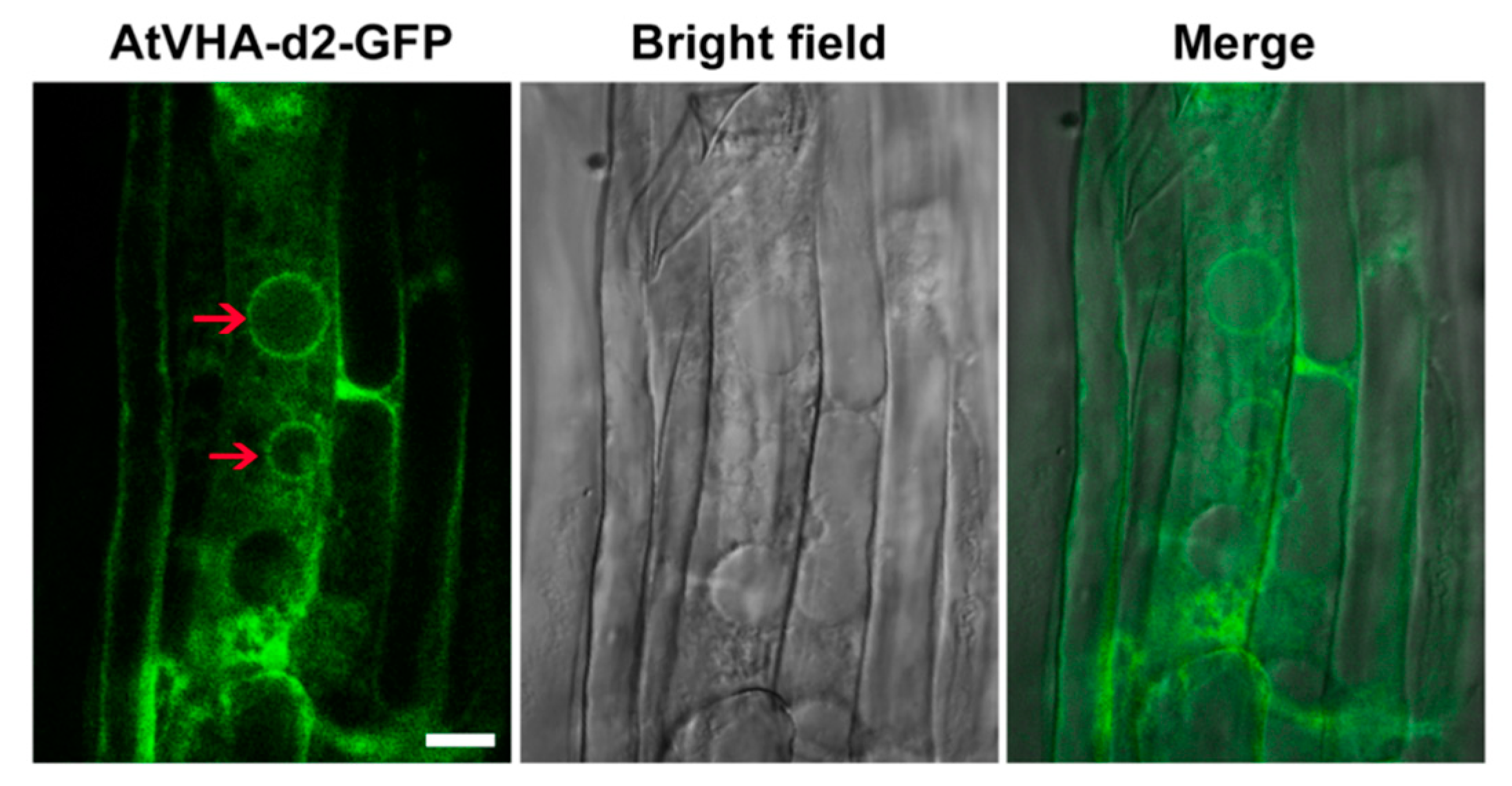
3.2. Subcellular Localization of AtVHA-d2
3.3. Sensitivity of the Atvha-d2 Mutant to Multiple Stresses
3.4. H+ Flux in Root of the Atvha-d2 Mutant under Multiple Stresses
3.5. Identification of DEGs between Atvha-d2 and Wild-Type under Normal and Oxidative Stress Conditions
3.6. Expression of AtAHA and AtAVP Genes in atvha-d2 and Wild-Type under Normal and Oxidative Stress Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gaxiola, R.A.; Palmgren, M.G.; Schumacher, K. Plant proton pumps. FEBS Lett. 2007, 581, 2204–2214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, K. Endomembrane proton pumps: Connecting membrane and vesicle transport. Curr. Opin. Plant Biol. 2006, 9, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, K. pH in the plant endomembrane system-an import and export business. Curr. Opin. Plant Biol. 2014, 22, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zeng, Y.; Zhuang, X.; Sun, L.; Yao, X.; Pimpl, P.; Jiang, L. Organelle pH in the Arabidopsis endomembrane system. Mol. Plant 2013, 6, 1419–1437. [Google Scholar] [CrossRef] [Green Version]
- Forgac, M. Vacuolar ATPases: Rotary proton pumps in physiology and pathophysiology. Nat. Rev. Mol. Cell Biol. 2007, 8, 917–929. [Google Scholar] [CrossRef]
- Schumacher, K.; Krebs, M. The V-ATPase: Small cargo, large effects. Curr. Opin. Plant Biol. 2010, 13, 724–730. [Google Scholar] [CrossRef]
- Schnitzer, D.; Seidel, T.; Sander, T.; Golldack, D.; Dietz, K.J. The cellular energization state affects peripheral stalk stability of plant vacuolar H+-ATPase and impairs vacuolar acidification. Plant Cell Physiol. 2011, 52, 946–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dettmer, J.; Hong-Hermesdorf, A.; Stierhof, Y.D.; Schumacher, K. Vacuolar H+-ATPase activity is required for endocytic and secretory trafficking in Arabidopsis. Plant Cell 2006, 18, 715–730. [Google Scholar] [CrossRef] [Green Version]
- Krebs, M.; Beyhl, D.; Gorlich, E.; Al-Rasheid, K.A.; Marten, I.; Stierhof, Y.D.; Hedrich, R.; Schumacher, K. Arabidopsis V-ATPase activity at the tonoplast is required for efficient nutrient storage but not for sodium accumulation. Proc. Natl. Acad. Sci. USA 2010, 107, 3251–3256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Scholl, S.; Doering, A.; Zhang, Y.; Irani, N.G.; Rubbo, S.D.; Neumetzler, L.; Krishnamoorthy, P.; Van Houtte, I.; Mylle, E.; et al. V-ATPase activity in the TGN/EE is required for exocytosis and recycling in Arabidopsis. Nat Plants 2015, 1, 15094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, K.; Vafeados, D.; McCarthy, M.; Sze, H.; Wilkins, T.; Chory, J. The Arabidopsis det3 mutant reveals a central role for the vacuolar H(+)-ATPase in plant growth and development. Genes Dev. 1999, 13, 3259–3270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batelli, G.; Verslues, P.E.; Agius, F.; Qiu, Q.; Fujii, H.; Pan, S.; Schumaker, K.S.; Grillo, S.; Zhu, J.K. SOS2 promotes salt tolerance in part by interacting with the vacuolar H+-ATPase and upregulating its transport activity. Mol. Cell. Biol. 2007, 27, 7781–7790. [Google Scholar] [CrossRef] [Green Version]
- Zhou, A.; Liu, E.; Ma, H.; Feng, S.; Gong, S.; Wang, J. NaCl-induced expression of AtVHA-c5 gene in the roots plays a role in response of Arabidopsis to salt stress. Plant Cell Rep. 2018, 37, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Bu, Y.; Takano, T.; Zhang, X.; Liu, S. Conserved V-ATPase c subunit plays a role in plant growth by influencing V-ATPase-dependent endosomal trafficking. Plant Biotechnol. J. 2016, 14, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Milgrom, E.; Diab, H.; Middleton, F.; Kane, P.M. Loss of vacuolar proton-translocating ATPase activity in yeast results in chronic oxidative stress. J. Biol. Chem. 2007, 282, 7125–7136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikawa, H.; Miyazaki, T.; Nakayama, H.; Minematsu, A.; Yamauchi, S.; Yamashita, K.; Takazono, T.; Shimamura, S.; Nakamura, S.; Izumikawa, K.; et al. Roles of vacuolar H+-ATPase in the oxidative stress response of Candida glabrata. FEMS Yeast Res. 2016, 16, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hager, A.; Lanz, C. Essential sulfhydryl groups in the catalytic center of the tonoplast H(+)-ATPase from coleoptiles of Zea mays L. as demonstrated by the biotin-streptavidin-peroxidase system. Planta 1989, 180, 116–122. [Google Scholar] [CrossRef]
- Feng, Y.; Forgac, M. Cysteine 254 of the 73-kDa A subunit is responsible for inhibition of the coated vesicle (H+)-ATPase upon modification by sulfhydryl reagents. J. Biol. Chem. 1992, 267, 5817–5822. [Google Scholar]
- Dschida, W.J.; Bowman, B.J. The vacuolar ATPase: Sulfite stabilization and the mechanism of nitrate inactivation. J. Biol. Chem. 1995, 270, 1557–1563. [Google Scholar] [CrossRef] [Green Version]
- Seidel, T.; Scholl, S.; Krebs, M.; Rienmuller, F.; Marten, I.; Hedrich, R.; Hanitzsch, M.; Janetzki, P.; Dietz, K.J.; Schumacher, K. Regulation of the V-type ATPase by redox modulation. Biochem. J. 2012, 448, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Thaker, Y.R.; Roessle, M.; Grüber., G. The boxing glove shape of subunit d of the yeast V-ATPase in solution and the importance of disulfide formation for folding of this protein. J. Bioenerg. Biomembr. 2007, 39, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Owegi, M.A.; Pappas, D.L.; Finch, M.W., Jr.; Bilbo, S.A.; Resendiz, C.A.; Jacquemin, L.J.; Warrier, A.; Trombley, J.D.; McCulloch, K.M.; Margalef, K.L.; et al. Identification of a domain in the V0 subunit d that is critical for coupling of the yeast vacuolar proton-translocating ATPase. J. Biol. Chem. 2006, 281, 30001–30014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Zhao, Y.J.; Zhao, B.C.; Ge, R.C.; Li, M.; Shen, Y.Z.; Huang, Z.J. Cloning and functional analysis of wheat V-H+-ATPase subunit genes. Plant Mol. Biol. 2009, 69, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Sze, H.; Schumacher, K.; Muller, M.L.; Padmanaban, S.; Taiz, L. A simple nomenclature for a complex proton pump: VHA genes encode the vacuolar H(+)-ATPase. Trends Plant Sci. 2002, 7, 157–161. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Chen, S.; Dai, S.; Wang, R.; Li, N.; Shen, X.; Zhou, X.; Lu, C.; Zheng, X.; Hu, Z.; et al. NaCl-induced alternations of cellular and tissue ion fluxes in roots of salt-resistant and salt-sensitive poplar species. Plant Physiol. 2009, 149, 1141–1153. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Zhao, J.; Feng, S.; Qiao, K.; Gong, S.; Wang, J.; Zhou, A. Heterologous expression of nitrate assimilation related-protein DsNAR2.1/NRT3.1 affects uptake of nitrate and ammonium in nitrogen-starved Arabidopsis. Int. J. Mol. Sci. 2020, 21, 4027. [Google Scholar] [CrossRef]
- Zhou, A.; Ma, H.; Liu, E.; Jiang, T.; Feng, S.; Gong, S.; Wang, J. Transcriptome sequencing of Dianthus spiculifolius and analysis of the genes involved in responses to combined cold and drought stress. Int. J. Mol. Sci. 2017, 18, 849. [Google Scholar] [CrossRef]
- Zhou, A.; Sun, H.; Dai, S.; Feng, S.; Zhang, J.; Gong, S.; Wang, J. Identification of transcription factors involved in the regulation of flowering in Adonis amurensis through combined RNA-seq transcriptomics and iTRAQ proteomics. Genes 2019, 10, 305. [Google Scholar] [CrossRef] [Green Version]
- Padmanaban, S.; Lin, X.; Perera, I.; Kawamura, Y.; Sze, H. Differential expression of vacuolar H+-ATPase subunit c genes in tissues active in membrane trafficking and their roles in plant growth as revealed by RNAi. Plant Physiol. 2004, 134, 1514–1526. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Ni, Z.; Zhang, H.; Chen, Q.; Gao, W.; Cai, Y.; Li, M.; Sun, G.; Qu, Y.Y. The Gene encoding subunit A of the vacuolar H(+)-ATPase from cotton plays an important role in conferring tolerance to water deficit. Front. Plant Sci. 2018, 9, 758. [Google Scholar] [CrossRef]
- Haruta, M.; Burch, H.L.; Nelson, R.B.; Barrett-Wilt, G.; Kline, K.G.; Mohsin, S.B.; Young, J.C.; Otegui, M.S.; Sussman, M.R. Molecular characterization of mutant Arabidopsis plants with reduced plasma membrane proton pump activity. J. Biol. Chem. 2010, 285, 17918–17929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, I.; Tchieu, J.; Sussman, M.R.; Boutry, M.; Palmgren, M.G.; Gribskov, M.; Harper, J.F.; Axelsen, K.B. Genomic comparison of P-type ATPase ion pumps in Arabidopsis and rice. Plant Physiol. 2003, 132, 618–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, D.H.; Lee, S.Y.; Bressan, R.A.; Yun, D.J.; Bohnert, H.J. Intracellular consequences of SOS1 deficiency during salt stress. J. Exp. Bot. 2010, 61, 1205–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, S.; Peng, Y.; Liu, E.; Ma, H.; Qiao, K.; Zhou, A.; Liu, S.; Bu, Y. Arabidopsis V-ATPase d2 Subunit Plays a Role in Plant Responses to Oxidative Stress. Genes 2020, 11, 701. https://doi.org/10.3390/genes11060701
Feng S, Peng Y, Liu E, Ma H, Qiao K, Zhou A, Liu S, Bu Y. Arabidopsis V-ATPase d2 Subunit Plays a Role in Plant Responses to Oxidative Stress. Genes. 2020; 11(6):701. https://doi.org/10.3390/genes11060701
Chicago/Turabian StyleFeng, Shuang, Yun Peng, Enhui Liu, Hongping Ma, Kun Qiao, Aimin Zhou, Shenkui Liu, and Yuanyuan Bu. 2020. "Arabidopsis V-ATPase d2 Subunit Plays a Role in Plant Responses to Oxidative Stress" Genes 11, no. 6: 701. https://doi.org/10.3390/genes11060701
APA StyleFeng, S., Peng, Y., Liu, E., Ma, H., Qiao, K., Zhou, A., Liu, S., & Bu, Y. (2020). Arabidopsis V-ATPase d2 Subunit Plays a Role in Plant Responses to Oxidative Stress. Genes, 11(6), 701. https://doi.org/10.3390/genes11060701