Genome-Wide Identification of the CrRLK1L Subfamily and Comparative Analysis of Its Role in the Legume-Rhizobia Symbiosis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of CrRLK1 Subfamily Proteins in 62 Plant Species
2.2. Phylogenetic Analysis of the CrRLK1L Subfamily
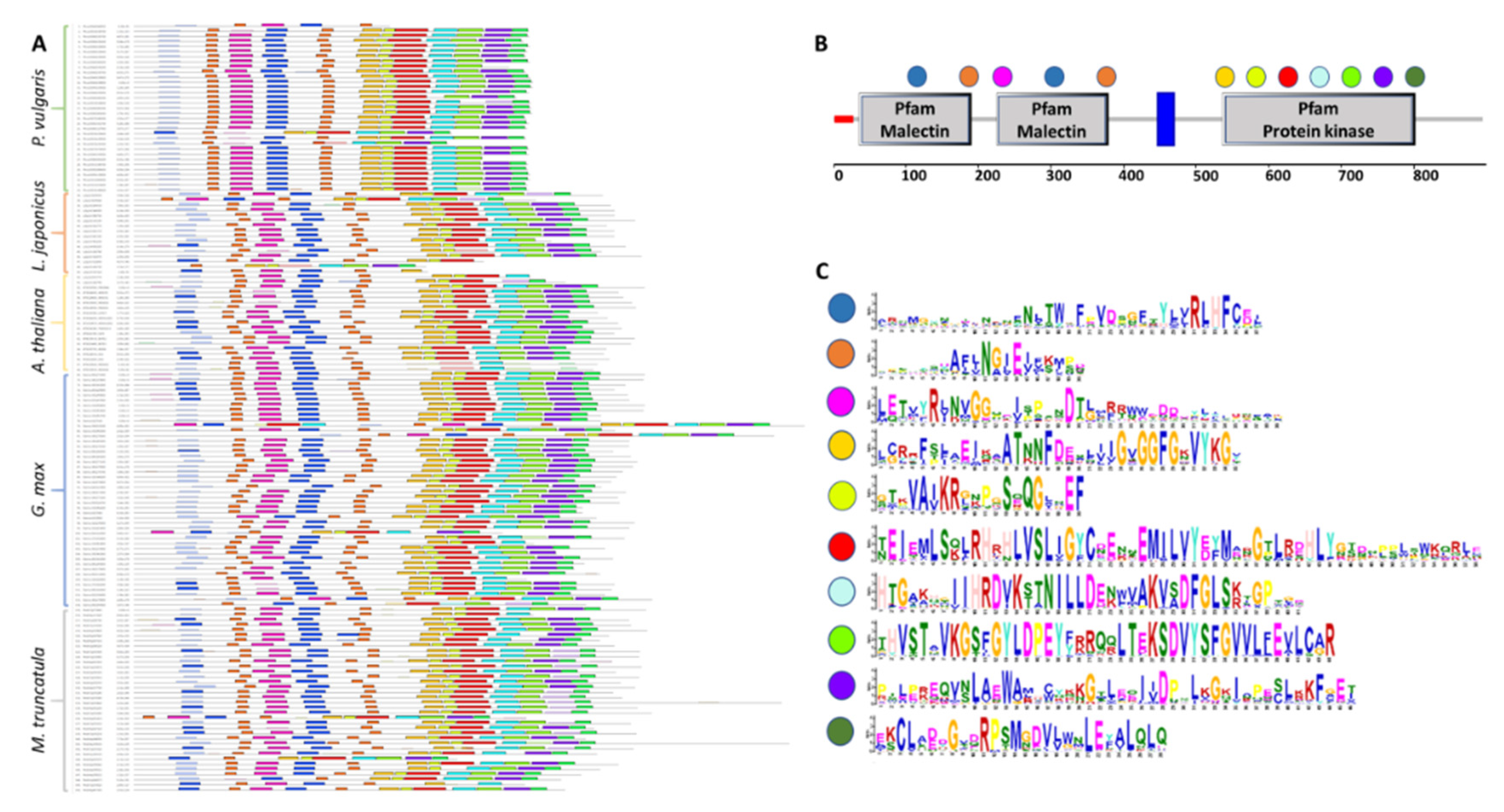
2.3. Analysis of CrRLK1L Protein Motif Conservation in Legumes and A. thaliana
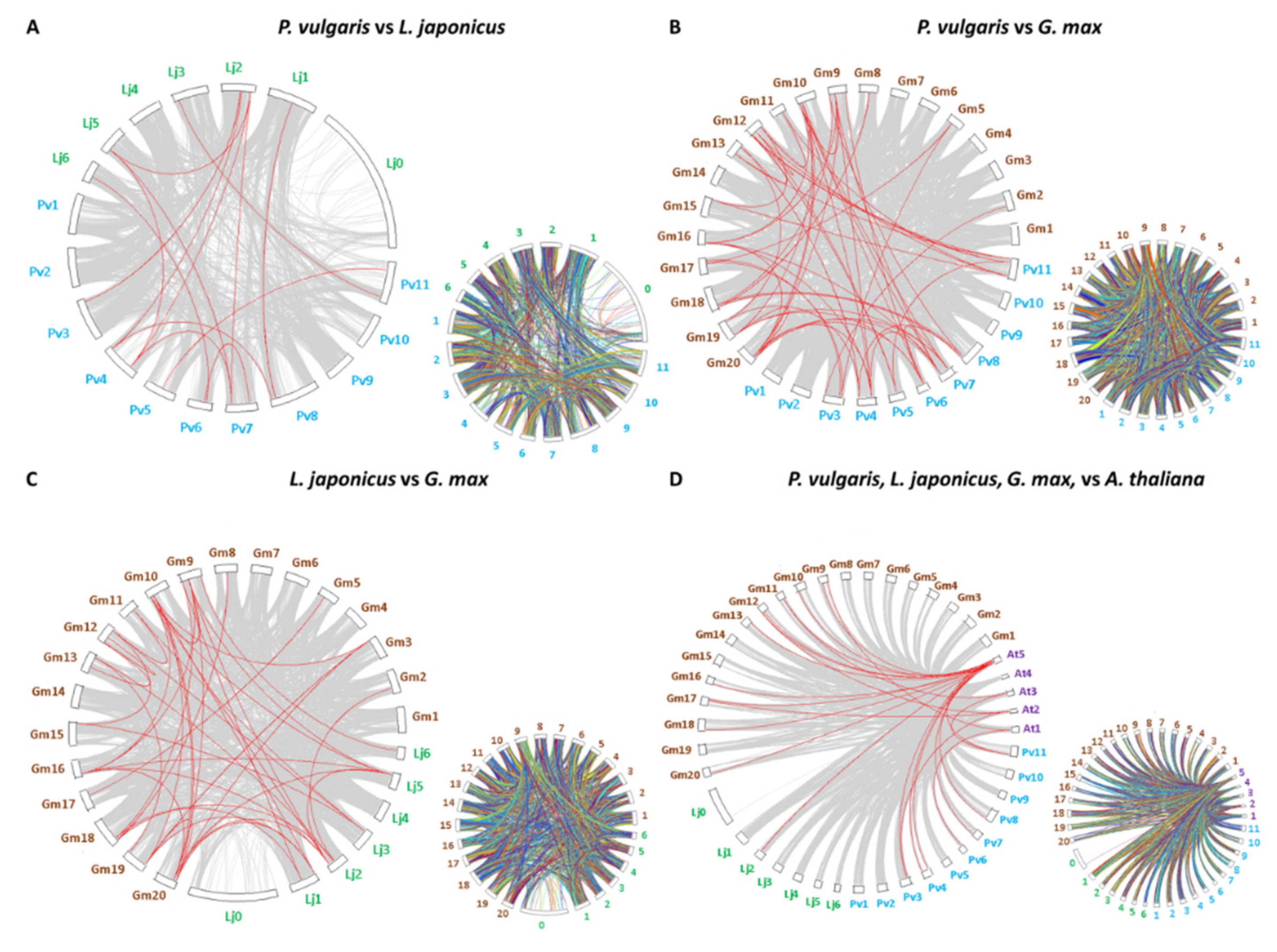
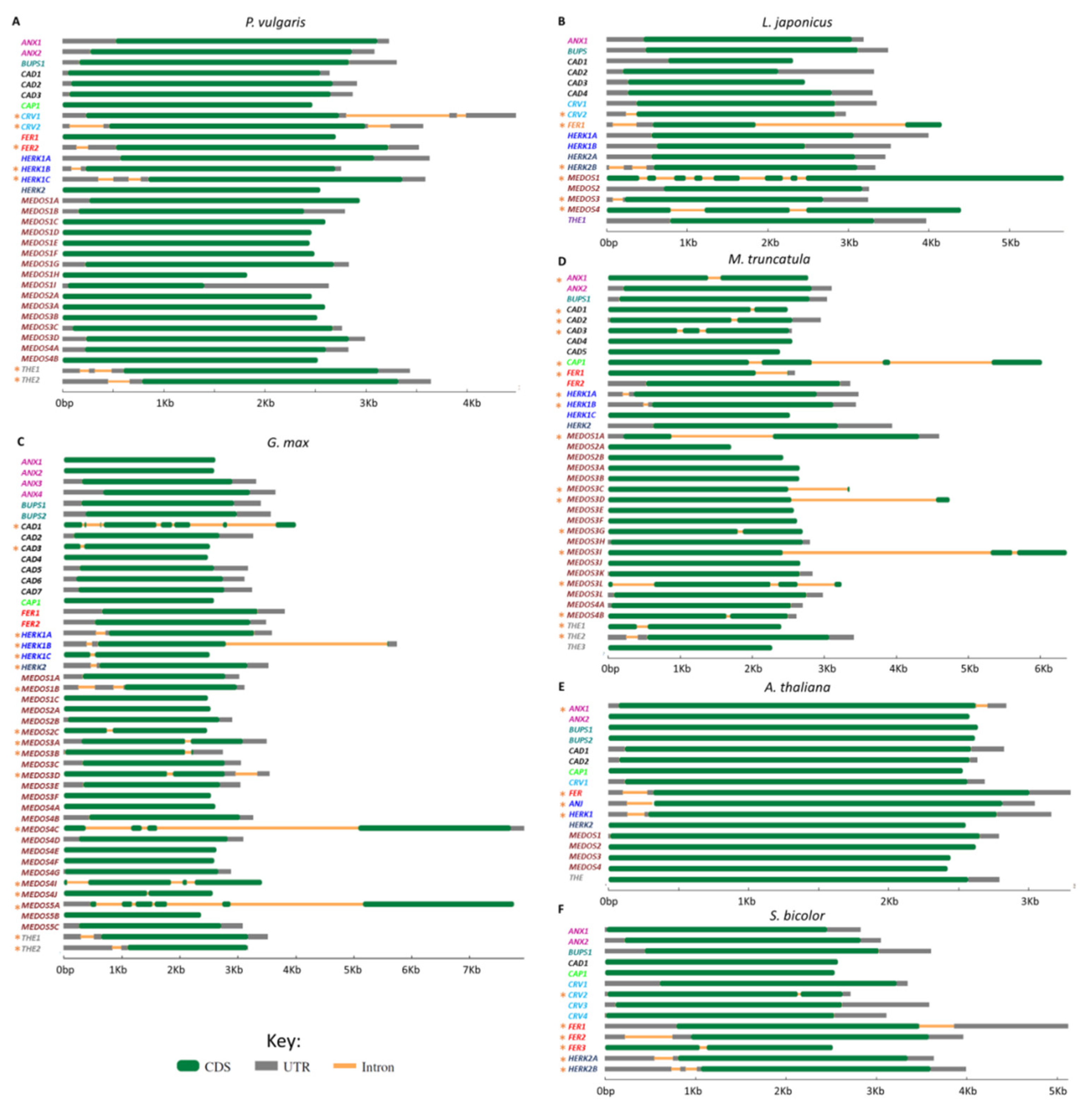
2.4. Gene Structure, Chromosomal Localization, and Synteny Analysis of the CrRLK1L Gene Subfamily of Legumes, A. thaliana, and Sorgum bicolor (L.), Moench
2.5. In Silico Analysis of the CrRLK1L Gene Family Expression in Legumes, A. thaliana, and P. patens
2.6. Plant Growth Conditions and RT-qPCR Assays
3. Results
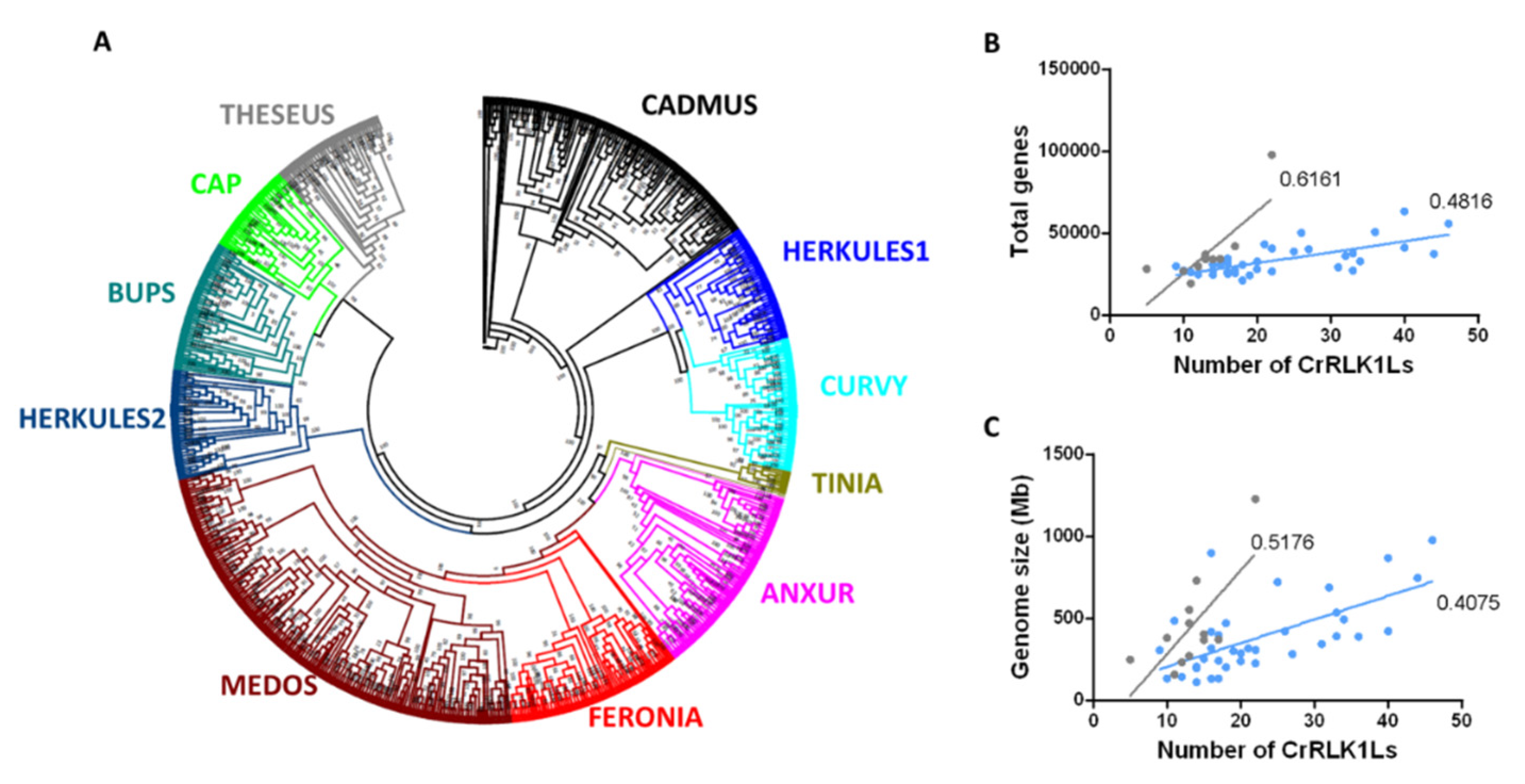
3.1. Identification and Phylogenetic Analysis of CrRLK1L Proteins in Diverse Plant Species
3.2. Phylogenetic Analysis of the CrRLK1L Subfamily in Legumes, A. thaliana, and P. patens
3.3. Features of the CrRLK1L Subfamily Proteins in Legumes and A. thaliana
3.4. Chromosomal Localization and Synteny of CrRLK1L Genes in Legumes and A. thaliana
3.5. Exon–Intron Structure of CrRLK1L Genes in Legumes and A. thaliana
3.6. Analysis of the Expression Patterns of the CrRLK1L Genes in Legumes, A. thaliana, and P. patens
3.6.1. FER Genes Are Broadly Expressed in All Tissues in Four Different Plant Species
3.6.2. ANX, BUPS, and CAP Genes Are Expressed Only in A. thaliana Pollen Tubes and G. max Flowers
3.6.3. HERK and THE Genes Are Expressed in Roots, Leaves, and Pods/Siliques
3.6.4. MEDOS Genes Are Mostly Expressed in Leaves
3.6.5. CRV Gene Expression Is Observed in Roots and Leaves, but Is Absent in G. max
3.6.6. The Tissue Specificity of CAD Gene Expression Is Broader in Legumes Than in A. thaliana
3.6.7. P. patens CrRLK1L Genes Are Widely but Differentially Expressed in All Tissues Tested
3.6.8. Certain CrRLK1L Genes Are Differentially Expressed during Nodulation
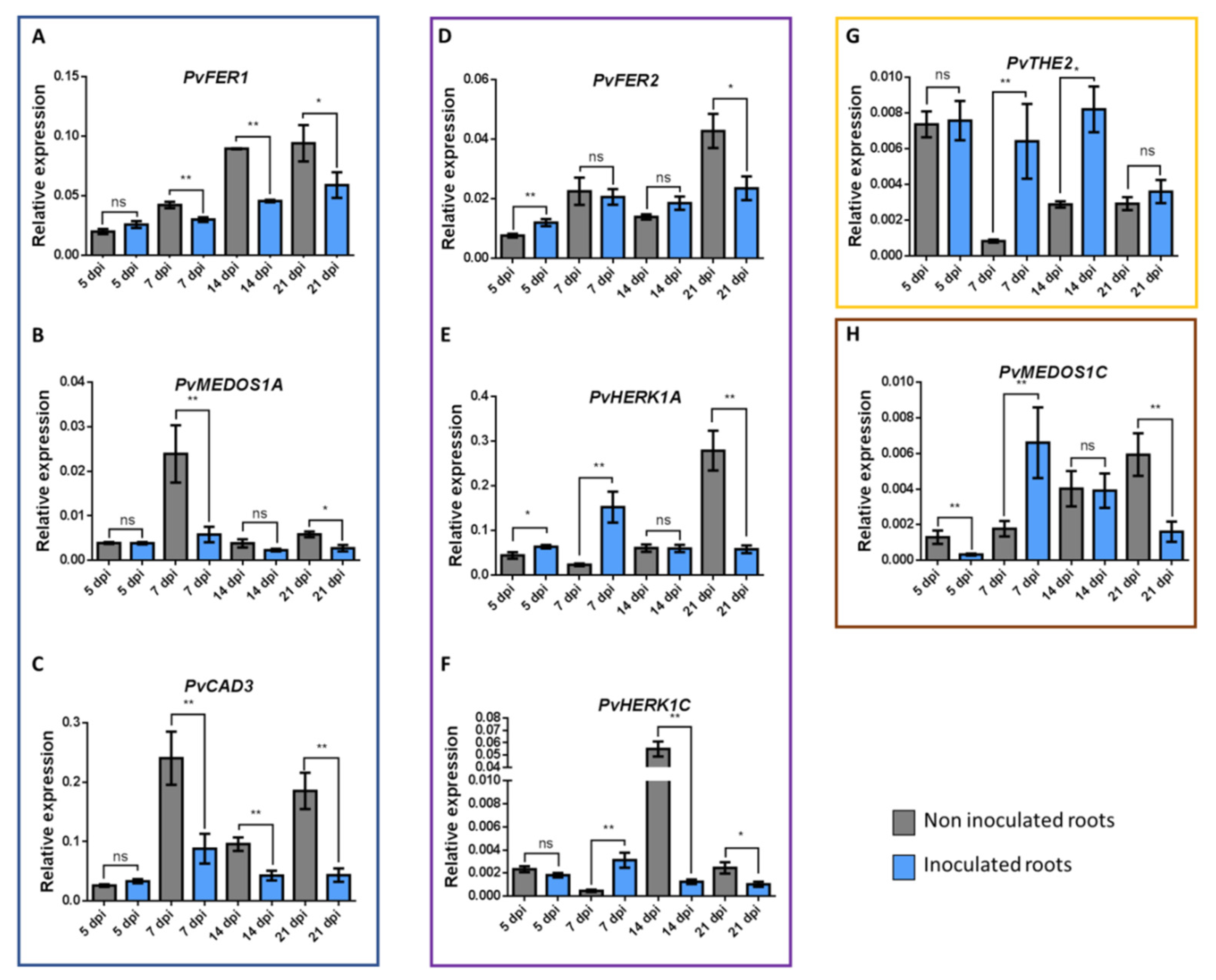
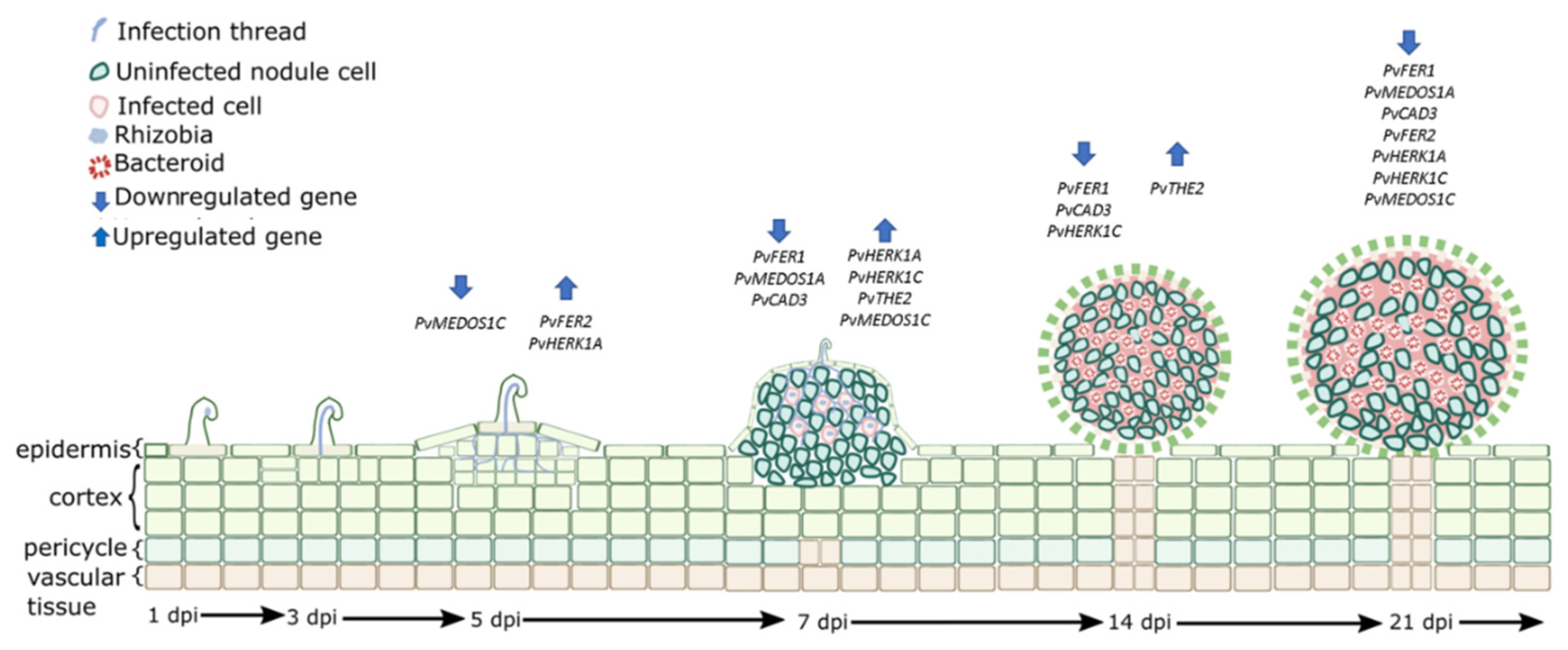
3.7. Expression of CrRLK1L Genes in P. vulgaris Nodules
4. Discussion
4.1. Structural Features of CrRLK1L Genes
4.2. Differences and Similarities in the Expression of CrRLK1L Genes in Legumes and in A. thaliana
4.3. Putative Roles of CrRLK1L Genes during Nodulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shiu, S.-H.; Bleecker, A.B. Receptor-like kinases from Arabidopsis form a monophyletic gene family related to animal receptor kinases. Proc. Natl. Acad. Sci. USA 2001, 98, 10763–10768. [Google Scholar] [CrossRef] [Green Version]
- Schulze-Muth, P.; Irmler, S.; Schröder, G.; Schröder, J. Novel type of receptor-like protein kinase from a higher plant (Catharanthus roseus): cDNA, gene, intramolecular autophosphorylation, and identification of a threonine important for auto- and substrate phosphorylation. J. Biol. Chem. 1996, 271, 26684–26689. [Google Scholar] [CrossRef] [Green Version]
- Boisson-Dernier, A.; Kessler, S.A.; Grossniklaus, U. The walls have ears: The role of plant CrRLK1Ls in sensing and transducing extracellular signals. J. Exp. Bot. 2011, 62, 1581–1591. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Li, L.; Ye, H.; Yu, X.; Algreen, A.; Yin, Y. Three related receptor-like kinases are required for optimal cell elongation in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2009, 106, 7648–7653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DesLauriers, S.D.; Larsen, P.B. Feronia is a Key Modulator of Brassinosteroid and Ethylene Responsiveness in Arabidopsis Hypocotyls. Mol. Plant 2010, 3, 626–640. [Google Scholar] [CrossRef]
- Yu, F.; Qian, L.; Nibau, C.; Duan, Q.; Kita, D.; Levasseur, K.; Li, X.; Lu, C.; Li, H.; Hou, C.; et al. FERONIA receptor kinase pathway suppresses abscisic acid signaling in Arabidopsis by activating ABI2 phosphatase. Proc. Natl. Acad. Sci. USA 2012, 109, 14693–14698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, D.; Yu, F.; Li, J.; Van De Poel, B.; Tan, D.; Li, J.; Liu, Y.; Li, X.; Dong, M.; Chen, L.; et al. FERONIA receptor kinase interacts withS-adenosylmethionine synthetase and suppressesS-adenosylmethionine production and ethylene biosynthesis inArabidopsis. Plant Cell Environ. 2015, 38, 2566–2574. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.; Tang, R.; Zhang, X.; Luan, S.; Yu, F. FERONIA Receptor Kinase at the Crossroads of Hormone Signaling and Stress Responses. Plant Cell Physiol. 2017, 58, 1143–1150. [Google Scholar] [CrossRef]
- Guo, H.; Nolan, T.M.; Song, G.; Liu, S.; Xie, Z.; Chen, J.; Schnable, P.S.; Walley, J.W.; Yin, Y. FERONIA Receptor Kinase Contributes to Plant Immunity by Suppressing Jasmonic Acid Signaling in Arabidopsis thaliana. Curr. Biol. 2018, 28, 3316–3324. [Google Scholar] [CrossRef] [Green Version]
- Duan, Q.; Kita, D.; Li, C.; Cheung, A.Y.; Wu, H.-M. FERONIA receptor-like kinase regulates RHO GTPase signaling of root hair development. Proc. Natl. Acad. Sci. USA 2010, 107, 17821–17826. [Google Scholar] [CrossRef] [Green Version]
- Kessler, S.; Shimosato-Asano, H.; Keinath, N.F.; Wuest, S.E.; Ingram, G.C.; Panstruga, R.; Grossniklaus, U. Conserved Molecular Components for Pollen Tube Reception and Fungal Invasion. Science 2010, 330, 968–971. [Google Scholar] [CrossRef]
- Shen, Q.; Bourdais, G.; Pan, H.; Robatzek, S.; Tang, D. Arabidopsis glycosylphosphatidylinositol-anchored protein LLG1 associates with and modulates FLS2 to regulate innate immunity. Proc. Natl. Acad. Sci. USA 2017, 114, 5749–5754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Estévez, J.M.; Liao, H.; Zhu, Y.; Yang, T.; Li, C.; Wang, Y.; Li, L.; Liu, X.; Pacheco, J.M.; et al. The RALF1–FERONIA Complex Phosphorylates eIF4E1 to Promote Protein Synthesis and Polar Root Hair Growth. Mol. Plant 2020, 13, 698–716. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Chen, W.; Song, L.; Chen, Q.; Zhang, H.; Liao, H.; Zhao, G.; Lin, F.; Zhou, H.; Yu, F. Feronia phosphorylates E3 ubiquitin ligase ATL6 to modulate the stability of 14-3-3 proteins in response to the carbon/nitrogen ratio. J. Exp. Bot. 2019, 70, 6375–6388. [Google Scholar] [CrossRef] [PubMed]
- Huck, N.; Moore, J.M.; Federer, M.; Grossniklaus, U. The Arabidopsis mutant feronia disrupts the female gametophytic control of pollen tube reception. Development 2003, 130, 2149–2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escobar-Restrepo, J.-M.; Huck, N.; Kessler, S.A.; Gagliardini, V.; Gheyselinck, J.; Yang, W.-C.; Grossniklaus, U. The FERONIA Receptor-like Kinase Mediates Male-Female Interactions during Pollen Tube Reception. Science 2007, 317, 656–660. [Google Scholar] [CrossRef]
- Rotman, N.; Gourgues, M.; Guitton, A.-E.; Faure, J.-E.; Berger, F. A Dialogue between the Sirène Pathway in Synergids and the Fertilization Independent Seed Pathway in the Central Cell Controls Male Gamete Release during Double Fertilization in Arabidopsis. Mol. Plant 2008, 1, 659–666. [Google Scholar] [CrossRef]
- Ngo, Q.A.; Vogler, H.; Lituiev, D.S.; Nestorova, A.; Grossniklaus, U. A Calcium Dialog Mediated by the FERONIA Signal Transduction Pathway Controls Plant Sperm Delivery. Dev. Cell 2014, 29, 491–500. [Google Scholar] [CrossRef] [Green Version]
- Kessler, S.A.; Lindner, H.; Jones, D.S.; Grossniklaus, U. Functional analysis of related Cr RLK 1L receptor-like kinases in pollen tube reception. EMBO Rep. 2014, 16, 107–115. [Google Scholar] [CrossRef]
- Stegmann, M.; Monaghan, J.; Smakowska-Luzan, E.; Rovenich, H.; Lehner, A.; Holton, N.; Belkhadir, Y.; Zipfel, C. The receptor kinase FER is a RALF-regulated scaffold controlling plant immune signaling. Science 2017, 355, 287–289. [Google Scholar] [CrossRef] [Green Version]
- Campos, W.F.; Dressano, K.; Ceciliato, P.H.O.; Guerrero-Abad, J.C.; Silva, A.L.; Fiori, C.S.; Canto, A.M.D.; Bergonci, T.; Claus, L.A.N.; Silva-Filho, M.C.; et al. Arabidopsis thaliana rapid alkalinization factor 1–mediated root growth inhibition is dependent on calmodulin-like protein 38. J. Biol. Chem. 2017, 293, 2159–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Li, J.; Huang, Y.; Liu, L.; Li, D.; Chen, L.; Luan, S. FERONIA Receptor Kinase Controls Seed Size in Arabidopsis thaliana. Mol. Plant 2014, 7, 920–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Zayed, O.; Yu, Z.; Jiang, W.; Zhu, P.; Hsu, C.-C.; Zhang, L.; Tao, W.A.; Lozano-Durán, R.; Zhu, J. Leucine-rich repeat extensin proteins regulate plant salt tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, 13123–13128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Wang, L.; Li, C.; Liu, Y.; Zhu, S.; Qi, Y.; Liu, X.; Lin, Q.; Luan, S.; Yu, F. Receptor protein kinase FERONIA controls leaf starch accumulation by interacting with glyceraldehyde-3-phosphate dehydrogenase. Biochem. Biophys. Res. Commun. 2015, 465, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Liu, X.; Qiang, X.; Li, X.; Li, X.; Zhu, S.; Wang, L.; Wang, Y.; Liao, H.; Luan, S.; et al. EBP1 nuclear accumulation negatively feeds back on FERONIA-mediated RALF1 signaling. PLoS Biol. 2018, 16, e2006340. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yang, T.; Lin, Q.; Wang, B.; Li, X.; Luan, S.; Yu, F. Receptor kinase FERONIA regulates flowering time in Arabidopsis. BMC Plant Biol. 2020, 20, 26. [Google Scholar] [CrossRef] [Green Version]
- Richter, J.; Ploderer, M.; Mongelard, G.; Gutierrez, L.; Hauser, M.-T. Role of CrRLK1L Cell Wall Sensors HERCULES1 and 2, THESEUS1, and FERONIA in Growth Adaptation Triggered by Heavy Metals and Trace Elements. Front. Plant Sci. 2017, 8, 1554. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Xing, J.; Wang, L.; Liu, Y.; Qu, J.; Tan, Y.; Fu, X.; Lin, Q.; Deng, H.; Yu, F. Mutations of two FERONIA-like receptor genes enhance rice blast resistance without growth penalty. J. Exp. Bot. 2020, 71, 2112–2126. [Google Scholar] [CrossRef]
- Jia, M.; Ding, N.; Zhang, Q.; Xing, S.; Wei, L.; Zhao, Y.; Du, P.; Mao, W.; Li, J.; Li, B.; et al. A FERONIA-Like Receptor Kinase Regulates Strawberry (Fragaria × ananassa) Fruit Ripening and Quality Formation. Front. Plant Sci. 2017, 8, 1099. [Google Scholar] [CrossRef]
- Zhu, L.; Chu, L.-C.; Liang, Y.; Zhang, X.-Q.; Chen, L.-Q.; Ye, D. The Arabidopsis CrRLK1L protein kinases BUPS1 and BUPS2 are required for normal growth of pollen tubes in the pistil. Plant J. 2018, 95, 474–486. [Google Scholar] [CrossRef] [Green Version]
- Boisson-Dernier, A.; Roy, S.; Kritsas, K.; Grobei, M.A.; Jaciubek, M.; Schroeder, J.I.; Grossniklaus, U. Disruption of the pollen-expressed FERONIA homologs ANXUR1 and ANXUR2 triggers pollen tube discharge. Development 2009, 136, 3279–3288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boisson-Dernier, A.; Lituiev, D.S.; Nestorova, A.; Franck, C.M.; Thirugnanarajah, S.; Grossniklaus, U. ANXUR receptor-like kinases coordinate cell wall integrity with growth at the pollen tube tip via NADPH oxidases. PLoS Biol. 2013, 11, e1001719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galindo-Trigo, S.; Blanco-Touriñán, N.; DeFalco, T.A.; Wells, E.S.; Gray, J.E.; Zipfel, C.; Smith, L.M. Cr RLK 1L receptor-like kinases HERK 1 and ANJEA are female determinants of pollen tube reception. EMBO Rep. 2019, 21, e48466. [Google Scholar] [CrossRef]
- Hematy, K.; Höfte, H. Novel receptor kinases involved in growth regulation. Curr. Opin. Plant Biol. 2008, 11, 321–328. [Google Scholar] [CrossRef]
- Schoenaers, S.; Balcerowicz, D.; Costa, A.; Vissenberg, K. The Kinase ERULUS Controls Pollen Tube Targeting and Growth in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoenaers, S.; Balcerowicz, D.; Breen, G.; Hill, K.; Zdanio, M.; Mouille, G.; Holman, T.J.; Oh, J.; Wilson, M.H.; Nikonorova, N.; et al. The Auxin-Regulated CrRLK1L Kinase ERULUS Controls Cell Wall Composition during Root Hair Tip Growth. Curr. Biol. 2018, 28, 722–732. [Google Scholar] [CrossRef] [Green Version]
- Gachomo, E.W.; Jno Baptiste, L.; Kefela, T.; Saidel, W.M.; Kotchoni, S.O. The Arabidopsis CURVY1 (CVY1) gene encoding a novel receptor-like protein kinase regulates cell morphogenesis, flowering time and seed production. BMC Plant Biol. 2014, 14, 221. [Google Scholar] [CrossRef] [Green Version]
- Richter, J.; Watson, J.M.; Stasnik, P.; Borowska, M.; Neuhold, J.; Berger, M.; Stolt-Bergner, P.; Schoft, V.; Hauser, M.-T. Multiplex mutagenesis of four clustered CrRLK1L with CRISPR/Cas9 exposes their growth regulatory roles in response to metal ions. Sci. Rep. 2018, 8, 12182. [Google Scholar] [CrossRef]
- Haruta, M.; Sabat, G.; Stecker, K.; Minkoff, B.B.; Sussman, M.R. A Peptide Hormone and Its Receptor. Science 2014, 343, 408–411. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.; Bergonci, T.; Zhao, Y.; Zou, Y.; Du, S.; Liu, M.-C.; Luo, X.; Ruan, H.; García-Valencia, L.E.; Zhong, S.; et al. Arabidopsispollen tube integrity and sperm release are regulated by RALF-mediated signaling. Science 2017, 358, 1596–1600. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Kurten, E.L.; Monshausen, G.; Hummel, G.M.; Gilroy, S.; Baldwin, I.T. NaRALF, a peptide signal essential for the regulation of root hair tip apoplastic pH inNicotiana attenuata, is required for root hair development and plant growth in native soils. Plant J. 2007, 52, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Covey, P.A.; Subbaiah, C.C.; Parsons, R.L.; Pearce, G.; Lay, F.T.; Anderson, M.A.; Ryan, C.A.; Bedinger, P.A. A pollen-specific RALF from tomato that regulates pollen tube elongation. Plant Physiol. 2010, 153, 703–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Shi, F. Evolution of the RALF Gene Family in Plants: Gene Duplication and Selection Patterns. Evol. Bioinform. 2012, 8, 271–292. [Google Scholar] [CrossRef]
- Campbell, L.; Turner, S.R. A Comprehensive Analysis of RALF Proteins in Green Plants Suggests There Are Two Distinct Functional Groups. Front. Plant Sci. 2017, 8, 403. [Google Scholar] [CrossRef]
- Combier, J.-P.; Küster, H.; Journet, E.-P.; Hohnjec, N.; Gamas, P.; Niebel, A. Evidence for the Involvement in Nodulation of the Two Small Putative Regulatory Peptide-Encoding GenesMtRALFL1andMtDVL1. Mol. Plant Microbe Interact. 2008, 21, 1118–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, J.J.; Luckow, M.A. The Rest of the Iceberg. Legume Diversity and Evolution in a Phylogenetic Context. Plant Physiol. 2003, 131, 900–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, B.; Mens, C.; Hastwell, A.; Zhang, M.; Su, H.; Jones, C.M.; Chu, X.; Gresshoff, P.M. Legume nodulation: The host controls the party. Plant Cell Environ. 2018, 42, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Goodstein, D.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.H.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2011, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Mun, T.; Bachmann, A.; Gupta, V.; Stougaard, J.; Andersen, S.U. Lotus Base: An integrated information portal for the model legume Lotus japonicus. Sci. Rep. 2016, 6, 39447. [Google Scholar] [CrossRef] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O.; Dufayard, J.-F. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Bodén, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, USA, 2005; pp. 571–608. [Google Scholar] [CrossRef]
- Wolfe, D.; Dudek, S.; Ritchie, M.; Pendergrass, S.A. Visualizing genomic information across chromosomes with PhenoGram. BioData Min. 2013, 6, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- O’Rourke, J.A.; Iniguez, L.P.; Fu, F.; Bucciarelli, B.; Miller, S.S.; Jackson, S.A.; McClean, P.E.; Li, J.; Dai, X.; Zhao, P.X.; et al. An RNA-Seq based gene expression atlas of the common bean. BMC Genom. 2014, 15, 866. [Google Scholar] [CrossRef] [Green Version]
- Benedito, V.A.; Torres-Jerez, I.; Murray, J.D.; Andriankaja, A.; Allen, S.; Kakar, K.; Wandrey, M.; Verdier, J.; Zuber, H.; Ott, T.; et al. A gene expression atlas of the model legume Medicago truncatula. Plant J. 2008, 55, 504–513. [Google Scholar] [CrossRef]
- He, J.; Benedito, V.A.; Wang, M.; Murray, J.D.; Zhao, P.X.; Tang, Y.; Udvardi, M.K. The Medicago truncatula gene expression atlas web server. BMC Bioinform. 2009, 10, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, M.; Davison, T.S.; Henz, S.R.; Pape, U.J.; Demar, M.; Vingron, M.; Schölkopf, B.; Weigel, D.; Lohmann, J.U. A gene expression map of Arabidopsis thaliana development. Nat. Genet. 2005, 37, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Libault, M.; Farmer, A.; Joshi, T.; Takahashi, K.; Langley, R.J.; Franklin, L.D.; He, J.; Xu, D.; May, G.; Stacey, G. An integrated transcriptome atlas of the crop model Glycine max, and its use in comparative analyses in plants. Plant J. 2010, 63, 86–99. [Google Scholar] [CrossRef]
- Ortiz-Ramírez, C.; Hernandez-Coronado, M.; Thamm, A.; Catarino, B.; Wang, M.; Dolan, L.; Feijó, J.A.A.; Becker, J.D.D. A Transcriptome Atlas of Physcomitrella patens Provides Insights into the Evolution and Development of Land Plants. Mol. Plant 2016, 9, 205–220. [Google Scholar] [CrossRef] [Green Version]
- Warnes, G.R.; Bolker, B.; Huber, W.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S. Gplots: Various R Programming Tools for Plotting Data. Available online: https://CRAN.R-project.org/package=gplots (accessed on 12 October 2019).
- Islas, T.; Guillén, G.; Alvarado-Affantranger, X.; Lara-Flores, M.; Sánchez, F.; Villanueva, M.A. PvRACK1 Loss-of-Function Impairs Cell Expansion and Morphogenesis in Phaseolus vulgaris L. Root Nodules. Mol. Plant-Microbe Interact. 2011, 24, 819–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, Q.-N.; Lee, Y.-S.; Cho, L.-H.; Jeong, H.-J.; An, G.; Jung, K.-H. Genome-wide identification and analysis of Catharanthus roseus RLK1-like kinases in rice. Planta 2014, 241, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Niu, E.; Cai, C.; Zheng, Y.; Shang, X.; Fang, L.; Guo, W. Genome-wide analysis of CrRLK1L gene family in Gossypium and identification of candidate CrRLK1L genes related to fiber development. Mol. Genet. Genom. 2016, 291, 1137–1154. [Google Scholar] [CrossRef]
- Kou, X.; Qi, K.; Qiao, X.; Yin, H.; Liu, X.; Zhang, S.; Wu, J. Evolution, expression analysis, and functional verification of Catharanthus roseus RLK1-like kinase (CrRLK1L) family proteins in pear (Pyrus bretchneideri). Genomic 2017, 109, 290–301. [Google Scholar] [CrossRef]
- Morris, J.L.; Puttick, M.N.; Clark, J.W.; Edwards, D.; Kenrick, P.; Pressel, S.; Wellman, C.H.; Yang, Z.; Schneider, H.; Donoghue, P.C.J. The timescale of early land plant evolution. Proc. Natl. Acad. Sci. USA 2018, 115, E2274–E2283. [Google Scholar] [CrossRef] [Green Version]
- Niklas, K.J.; Kutschera, U. The evolution of the land plant life cycle. New Phytol. 2009, 185, 27–41. [Google Scholar] [CrossRef]
- Chang, C.-C.; Chen, H.-L.; Li, W.-H.; Chaw, S.-M. Dating the monocot-dicot divergence and the origin of core eudicots using whole chloroplast genomes. J. Mol. Evol. 2004, 58, 424–441. [Google Scholar] [CrossRef]
- Jiao, X.; Leebens-Mack, J.; Ayyampalayam, S.; Bowers, J.; McKain, M.R.; McNeal, J.R.; Rolf, M.; Ruzicka, D.R.; Wafula, E.K.; Wickett, N.J.; et al. A genome triplication associated with early diversification of the core eudicots. Genome Biol. 2012, 13, R3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, M.; Conery, J.S. The Evolutionary Fate and Consequences of Duplicate Genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [Green Version]
- Coghlan, A.; Eichler, E.E.; Oliver, S.G.; Paterson, A.H.; Stein, L. Chromosome evolution in eukaryotes: A multi-kingdom perspective. Trends Genet. 2005, 21, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Bowers, J.; Chapman, B.; Rong, J.; Paterson, A.H. Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature 2003, 422, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Freeling, M. Bias in Plant Gene Content Following Different Sorts of Duplication: Tandem, Whole-Genome, Segmental, or by Transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; Tang, H.; Tan, X.; Ficklin, S.P.; Feltus, F.A.; Paterson, A.H. Modes of Gene Duplication Contribute Differently to Genetic Novelty and Redundancy, but Show Parallels across Divergent Angiosperms. PLoS ONE 2011, 6, e28150. [Google Scholar] [CrossRef] [Green Version]
- Jami, S.K.; Clark, G.B.; Ayele, B.T.; Ashe, P.; Kirti, P.B. Genome-wide Comparative Analysis of Annexin Superfamily in Plants. PLoS ONE 2012, 7, e47801. [Google Scholar] [CrossRef] [Green Version]
- Carle-Urioste, J.C.; Brendel, V.; Walbot, V. A combinatorial role for exon, intron and splice site sequences in splicing in maize. Plant J. 1997, 11, 1253–1263. [Google Scholar] [CrossRef]
- Mattick, J.S.; Gagen, M.J. The evolution of controlled multitasked gene networks: The role of introns and other noncoding RNAs in the development of complex organisms. Mol. Biol. Evol. 2001, 18, 1611–1630. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, S.; Meshorer, E.; Ast, G. Chromatin organization marks exon-intron structure. Nat. Struct. Mol. Biol. 2009, 16, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhang, Y.; Zhang, W.; Yang, S.; Chen, J.-Q.; Tian, D. Patterns of exon-intron architecture variation of genes in eukaryotic genomes. BMC Genom. 2009, 10, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiraga, J.; Mackiewicz, P.; Mackiewicz, D.; Kowalczuk, M.; Biecek, P.; Polak, N.; Smolarczyk, K.; Dudek, M.R.; Cebrat, S. The relationships between the isoelectric point and: Length of proteins, taxonomy and ecology of organisms. BMC Genom. 2007, 8, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, J.J.; Egan, A.N. Dating the origins of polyploidy events. New Phytol. 2009, 186, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Schranz, M.E. Network approaches for plant phylogenomic synteny analysis. Curr. Opin. Plant Biol. 2017, 36, 129–134. [Google Scholar] [CrossRef]
- Schwartz, R.; Ting, C.S.; King, J. Whole Proteome pI Values Correlate with Subcellular Localizations of Proteins for Organisms within the Three Domains of Life. Genome Res. 2001, 11, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Weiller, G.F.; Caraux, G.; Silvester, N. The modal distribution of protein isoelectric points reflects amino acid properties rather than sequence evolution. Proteomics 2004, 4, 943–949. [Google Scholar] [CrossRef]
- Yoder, J.B. Understanding the coevolutionary dynamics of mutualism with population genomics. Am. J. Bot. 2016, 103, 1742–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Moura, G.G.D.; Remigi, P.; Masson-Boivin, C.; Capela, D. Experimental Evolution of Legume Symbionts: What Have We Learnt? Genes 2020, 11, 339. [Google Scholar] [CrossRef] [Green Version]
- De Mita, S.; Streng, A.; Bisseling, T.; Geurts, R. Evolution of a symbiotic receptor through gene duplications in the legume-rhizobium mutualism. New Phytol. 2013, 201, 961–972. [Google Scholar] [CrossRef]
- Patriarca, E.J.; Tatè, R.; Ferraioli, S.; Iaccarino, M. Organogenesis of Legume Root Nodules. Int. Rev. Cytol. 2004, 234, 201–262. [Google Scholar] [CrossRef] [PubMed]
- Monahan-Giovanelli, H.; Pinedo, C.A.; Gage, D.J. Architecture of Infection Thread Networks in Developing Root Nodules Induced by the Symbiotic Bacterium Sinorhizobium meliloti on Medicago truncatula. Plant Physiol. 2006, 140, 661–670. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, B.; Indrasumunar, A.; Hayashi, S.; Lin, M.-H.; Lin, Y.-H.; Reid, D.E.; Gresshoff, P.M. Molecular Analysis of Legume Nodule Development and Autoregulation. J. Integr. Plant Biol. 2010, 52, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Popp, C.; Ott, T. Regulation of signal transduction and bacterial infection during root nodule symbiosis. Curr. Opin. Plant Biol. 2011, 14, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.T.; Schilderink, S.; Moling, S.; Deinum, E.E.; Kondorosi, E.; Franssen, H.; Kulikova, O.; Niebel, A.; Bisseling, T. Fate map of Medicago truncatula root nodules. Development 2014, 141, 3517–3528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limpens, E.; Ivanov, S.; Van Esse, G.W.; Voets, G.; Fedorova, E.; Bisseling, T. Medicago N2-Fixing Symbiosomes Acquire the Endocytic Identity Marker Rab7 but Delay the Acquisition of Vacuolar Identity. Plant Cell 2009, 21, 2811–2828. [Google Scholar] [CrossRef] [Green Version]
- Gavrin, A.; Chiasson, D.; Ovchinnikova, E.; Kaiser, B.N.; Bisseling, T.; Fedorova, E.E. VAMP721a and VAMP721d are important for pectin dynamics and release of bacteria in soybean nodules. New Phytol. 2016, 210, 1011–1021. [Google Scholar] [CrossRef] [Green Version]
- Cheung, A.Y.; Wu, H.-M. THESEUS 1, FERONIA and relatives: A family of cell wall-sensing receptor kinases? Curr. Opin. Plant Biol. 2011, 14, 632–641. [Google Scholar] [CrossRef]
- Mecchia, M.A.; Santos-Fernandez, G.; Duss, N.N.; Somoza, S.C.; Boisson-Dernier, A.; Gagliardini, V.; Martinez-Bernardini, A.; Fabrice, T.; Ringli, C.; Muschietti, J.; et al. RALF4/19 peptides interact with LRX proteins to control pollen tube growth inArabidopsis. Science 2017, 358, 1600–1603. [Google Scholar] [CrossRef] [Green Version]
- Vogler, H.; Santos-Fernandez, G.; Mecchia, M.A.; Grossniklaus, U. To preserve or to destroy, that is the question: The role of the cell wall integrity pathway in pollen tube growth. Curr. Opin. Plant Biol. 2019, 52, 131–139. [Google Scholar] [CrossRef]
- Kuppusamy, K.T.; Ivashuta, S.; Bucciarelli, B.; Vance, C.P.; Gantt, J.S.; VandenBosch, K.A. Knockdown of CELL DIVISION CYCLE16 reveals an inverse relationship between lateral root and nodule numbers and a link to auxin in Medicago truncatula. Plant Physiol. 2009, 151, 1155–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzaki, T.; Yano, K.; Ito, M.; Umehara, Y.; Suganuma, N.; Kawaguchi, M. Positive and negative regulation of cortical cell division during root nodule development in Lotus japonicus is accompanied by auxin response. Development 2012, 139, 3997–4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, D.; Stacey, N.; Liu, C.-W.; Wen, J.; Mysore, K.S.; Torres-Jerez, I.; Vernié, T.; Tadege, M.; Zhou, C.; Wang, Z.-Y.; et al. Rhizobial infection is associated with the development of peripheral vasculature in nodules of Medicago truncatula. Plant Physiol. 2013, 162, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sogawa, A.; Yamazaki, A.; Yamasaki, H.; Komi, M.; Manabe, T.; Tajima, S.; Hayashi, M.; Nomura, M. SNARE Proteins LjVAMP72a and LjVAMP72b Are Required for Root Symbiosis and Root Hair Formation in Lotus japonicus. Front. Plant Sci. 2019, 9, 1992. [Google Scholar] [CrossRef] [PubMed]
- Foucher, F.; Kondorosi, E. Cell cycle regulation in the course of nodule organogenesis in Medicago. Plant Mol. Biol. 2000, 43, 773–786. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C.; Oldroyd, G.E.D. Plant signalling in symbiosis and immunity. Nature 2017, 543, 328–336. [Google Scholar] [CrossRef]
- Mergaert, P.; Uchiumi, T.; Alunni, B.; Evanno, G.; Cheron, A.; Catrice, O.; Mausset, A.-E.; Barloy-Hubler, F.; Galibert, F.; Kondorosi, A.; et al. Eukaryotic control on bacterial cell cycle and differentiation in the Rhizobium-legume symbiosis. Proc. Natl. Acad. Sci. USA 2006, 103, 5230–5235. [Google Scholar] [CrossRef] [Green Version]
- Ott, T.; Sullivan, J.; James, E.K.; Flemetakis, E.; Günther, C.S.; Gibon, Y.; Ronson, C.; Udvardi, M.K. Absence of Symbiotic Leghemoglobins Alters Bacteroid and Plant Cell Differentiation During Development ofLotus japonicusRoot Nodules. Mol. Plant-Microbe Interact. 2009, 22, 800–808. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Rubio, M.C.; Xin, X.; Zhang, B.; Fan, Q.; Wang, Q.; Ning, G.; Becana, M.; Duanmu, D. CRISPR/Cas9 knockout of leghemoglobin genes in Lotus japonicus uncovers their synergistic roles in symbiotic nitrogen fixation. New Phytol. 2019, 224, 818–832. [Google Scholar] [CrossRef]
- Torres, M.A.; Jones, J.D.; Dangl, J.L. Pathogen-induced, NADPH oxidase–derived reactive oxygen intermediates suppress spread of cell death in Arabidopsis thaliana. Nat. Genet. 2005, 37, 1130–1134. [Google Scholar] [CrossRef]
- Lohar, D.P.; Haridas, S.; VandenBosch, K.A.; Gantt, J.S. A transient decrease in reactive oxygen species in roots leads to root hair deformation in the legume-rhizobia symbiosis. New Phytol. 2006, 173, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Marino, D.; Andrio, E.; Danchin, E.G.J.; Oger, E.; Gucciardo, S.; Lambert, A.; Puppo, A.; Pauly, N. A Medicago truncatula NADPH oxidase is involved in symbiotic nodule functioning. New Phytol. 2010, 189, 580–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montiel, J.; Nava, N.; Cárdenas, L.; Sanchez-Lopez, R.; Arthikala, M.-K.; Santana, O.; Sánchez, F.; Quinto, C. A Phaseolus vulgaris NADPH Oxidase Gene is Required for Root Infection by Rhizobia. Plant Cell Physiol. 2012, 53, 1751–1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthikala, M.-K.; Montiel, J.; Sanchez-Lopez, R.; Nava, N.; Cárdenas, L.; Quinto, C. Respiratory Burst Oxidase Homolog Gene A Is Crucial for Rhizobium Infection and Nodule Maturation and Function in Common Bean. Front. Plant Sci. 2017, 8, 2003. [Google Scholar] [CrossRef] [Green Version]
- Montiel, J.; Fonseca-García, C.; Quinto, C. Phylogeny and Expression of NADPH Oxidases during Symbiotic Nodule Formation. Agriculture 2018, 8, 179. [Google Scholar] [CrossRef] [Green Version]
- Montiel, J.; Arthikala, M.-K.; Cardenas, L.; Quinto, C. Legume NADPH Oxidases Have Crucial Roles at Different Stages of Nodulation. Int. J. Mol. Sci. 2016, 17, 680. [Google Scholar] [CrossRef] [Green Version]
- Khadri, M.; Tejera, N.A.; Plá, C.L. Alleviation of Salt Stress in Common Bean (Phaseolus vulgaris) by Exogenous Abscisic Acid Supply. J. Plant Growth Regul. 2006, 25, 110–119. [Google Scholar] [CrossRef]
- Ding, Y.; Kaló, P.; Yendrek, C.; Sun, J.; Liang, Y.; Marsh, J.F.; Harris, J.M.; Oldroyd, G.E.D. Abscisic Acid Coordinates Nod Factor and Cytokinin Signaling during the Regulation of Nodulation in Medicago truncatula. Plant Cell 2008, 20, 2681–2695. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Mitchell, D.M.; Harris, J.M. Abscisic acid rescues the root meristem defects of the Medicago truncatula latd mutant. Dev. Biol. 2007, 304, 297–307. [Google Scholar] [CrossRef] [Green Version]
- Biswas, B.; Chan, P.K.; Gresshoff, P.M. A Novel ABA Insensitive Mutant of Lotus japonicus with a Wilty Phenotype Displays Unaltered Nodulation Regulation. Mol. Plant 2009, 2, 487–499. [Google Scholar] [CrossRef]
- Hunter, W.J. Influence of Root-Applied Epibrassinolide and Carbenoxolone on the Nodulation and Growth of Soybean (Glycine max L.) Seedlings. J. Agron. Crop Sci. 2001, 186, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, B.; Ross, J.J.; Reid, J.B. Nodulation Phenotypes of Gibberellin and Brassinosteroid Mutants of Pea1. Plant Physiol. 2005, 138, 2396–2405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vardhini, B.V.; Rao, S.S.R. Effect of brassionosteriods on nodulation and nitrogenase activity in groundnut (Arachis hypogaea L.). Plant Growth Regul. 1999, 28, 165–167. [Google Scholar] [CrossRef]
- Upreti, K.; Murti, G. Effects of Brassmosteroids on Growth, Nodulation, Phytohormone Content and Nitrogenase Activity in French Bean Under Water Stress. Biol. Plant. 2004, 48, 407–411. [Google Scholar] [CrossRef]
- Sun, J.; Cardoza, V.; Mitchell, D.M.; Bright, L.; Oldroyd, G.; Harris, J.M. Crosstalk between jasmonic acid, ethylene and Nod factor signaling allows integration of diverse inputs for regulation of nodulation. Plant J. 2006, 46, 961–970. [Google Scholar] [CrossRef]
- Nakagawa, T.; Kawaguchi, M. Shoot-applied MeJA Suppresses Root Nodulation in Lotus japonicus. Plant Cell Physiol. 2006, 47, 176–180. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, A.; Suriyagoda, L.; Shigeyama, T.; Tominaga, A.; Sasaki, M.; Hiratsuka, Y.; Yoshinaga, A.; Arima, S.; Agarie, S.; Sakai, T.; et al. Lotus japonicus nodulation is photomorphogenetically controlled by sensing the red/far red (R/FR) ratio through jasmonic acid (JA) signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 16837–16842. [Google Scholar] [CrossRef] [Green Version]
- Guinel, F.C.; Geil, R.D. A model for the development of the rhizobial and arbuscular mycorrhizal symbioses in legumes and its use to understand the roles of ethylene in the establishment of these two symbioses. Can. J. Bot. 2002, 80, 695–720. [Google Scholar] [CrossRef] [Green Version]
- Gresshoff, P.M.; Lohar, D.; Chan, P.-K.; Biswas, B.; Jiang, Q.; Reid, D.E.; Ferguson, B.; Stacey, G. Genetic analysis of ethylene regulation of legume nodulation. Plant Signal. Behav. 2009, 4, 818–823. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Plant Species | Mutant/RNAi | Phenotype | Reference |
|---|---|---|---|---|
| FERONIA | A. thaliana | fer | PT overgrowth, multiple PT reach one ovules | [15,16,17,18,19] |
| fer | Collapsed, burst and short RH | [10] | ||
| fer | Resistance to Powdery mildew infection, increased susceptibility to Pseudomonas syringae pv. tomaeum DC3000 | [11,20] | ||
| fer | Ethylene hypersensitivity, brassinosteroid insensitivity, abscisic acid hypersensitivity, an increase of s-adenosyl methionine synthesis, inhibition of jasmonic acid responses | [5,6,7,9] | ||
| FER RNAi | Dwarf phenotype | [4,21] | ||
| fer | Larger seed size | [22] | ||
| fer | Salt hypersensitivity | [23] | ||
| fer | Increased starch accumulation in a sucrose medium. Hypersensitivity to high carbon/nitrogen ratio | [14,24] | ||
| fer | Reduced induction of ErbB3-binding protein 1, alteration of ribosome synthesis | [13,25] | ||
| fer | Delay in the flowering time under long day condition | [26] | ||
| fer | Hypersensitivity to nickel, tolerance to cadmium, coper, zinc, and lead | [27] | ||
| Oriza sativa L. | flr2 | Enhanced resistance to Magnaporthe oryzae infection | [28] | |
| flr11 | Enhanced resistance to M. oryzae infection | [28] | ||
| Fragaria x ananassa | FaMRL47RNAi | Fruit ripening acceleration | [29] | |
| BUPS1/2 | A. thaliana | bups1 | PT overgrowth | [30] |
| bups1 bups2 | Enhanced bups1 phenotype: PT overgrowth | |||
| ANXUR1/2 | A. thaliana | anx1 anx2 | Reduced fertility, PT burst | [19,31,32] |
| HERKULES1 | A. thaliana | herk1 the | Dwarf plants | [19,27,33] |
| herk1 | Tolerance to cadmium, coper, nickel, and zinc | |||
| herk1 | PT overgrowth | |||
| HERKULES2 | A. thaliana | herk2 | Tolerance to cadmium, coper, nickel, and lead | [27] |
| ANJEA | A. thaliana | herk anj | PT overgrowth | [33] |
| THESEUS1 | A. thaliana | the prc | Rescues of hypocotyl growth but without prc cellulose deficiency phenotype | [34] |
| the | Hypersensitivity to lead and zinc, tolerance to nickel | [27] | ||
| herk1 the | Dwarf plants | [4] | ||
| CAP | A. thaliana | cap | Altered PT growth in low calcium | [35] |
| cap | RH bursting and bulging | [36] | ||
| CURVY | A. thaliana | crv | Distortion of trichomes, altered pavement morphology | [37] |
| MEDOS1-4 | A. thaliana | med1,2,3,4 | Reduced growth in presence of metal ions | [38] |
| Gene ID * | Gene Name | CDS Length, bp | Protein Length, aa | iP | Molecular Weight, kDa |
|---|---|---|---|---|---|
| P. vulgaris | |||||
| Phvul.006G102700 | ANX1 | 2589 | 862 | 5.71 | 95.82 |
| Phvul.007G188300 | ANX2 | 2589 | 862 | 5.6 | 96 |
| Phvul.011G210400 | BUPS | 2670 | 889 | 5.41 | 97.19 |
| Phvul.003G188000 | CAP | 2469 | 822 | 5.91 | 92.06 |
| Phvul.004G109500 | CRV1 | 2505 | 834 | 5.49 | 92.38 |
| Phvul.007G074000 | CRV2 | 2535 | 844 | 5.7 | 93.64 |
| Phvul.008G081000 | FER1 | 2700 | 899 | 5.99 | 98.37 |
| Phvul.008G082400 | FER2 | 2697 | 898 | 6.67 | 98.1 |
| Phvul.005G139800 | HERK1A | 2514 | 837 | 5.64 | 92.51 |
| Phvul.008G000200 | HERK1B | 2472 | 823 | 5.38 | 92.18 |
| Phvul.011G069600 | HERK1C | 2514 | 837 | 5.79 | 92.94 |
| Phvul.006G127900 | HERK2 | 2547 | 848 | 7.3 | 93.01 |
| Phvul.004G038800 | MEDOS1A | 2676 | 891 | 7.29 | 99.43 |
| Phvul.004G039200 | MEDOS1B | 2235 | 744 | 7.05 | 83.82 |
| Phvul.004G039600 | MEDOS1C | 2598 | 865 | 5.78 | 96.98 |
| Phvul.004G039700 | MEDOS1D | 2406 | 801 | 5.79 | 90.35 |
| Phvul.004G039800 | MEDOS1E | 2442 | 813 | 5.99 | 92.04 |
| Phvul.004G039900 | MEDOS1F | 2490 | 829 | 6.51 | 93.64 |
| Phvul.004G040000 | MEDOS1G | 2460 | 819 | 7.31 | 92.32 |
| Phvul.004G040300 | MEDOS1H | 1824 | 607 | 6.26 | 68.13 |
| Phvul.004G040901 | MEDOS1I | 1353 | 450 | 8.46 | 50.59 |
| Phvul.004G039400 | MEDOS2A | 2463 | 820 | 5.99 | 92.31 |
| Phvul.008G030200 | MEDOS3A | 2595 | 864 | 6.15 | 96.81 |
| Phvul.008G030400 | MEDOS3B | 2517 | 838 | 6.24 | 93.86 |
| Phvul.008G030700 | MEDOS3C | 2577 | 858 | 6.28 | 96.37 |
| Phvul.008G030800 | MEDOS3D | 2601 | 866 | 4.83 | 97.08 |
| Phvul.003G038700 | MEDOS4A | 2385 | 794 | 8.67 | 89.03 |
| Phvul.003G038800 | MEDOS4B | 2523 | 840 | 5.95 | 93.94 |
| Phvul.005G085600 | THE1 | 2523 | 840 | 5.93 | 92.73 |
| Phvul.011G148700 | THE2 | 2538 | 845 | 5.87 | 93.05 |
| Phvul.003G239300 | CAD1 | 2499 | 832 | 6.63 | 93.06 |
| Phvul.003G239400 | CAD2 | 2589 | 862 | 8.33 | 96.18 |
| Phvul.003G239500 | CAD3 | 2586 | 861 | 5.86 | 96.32 |
| G. max | |||||
| Glyma.03G247800 | ANXUR1 | 2610 | 869 | 5.24 | 96.22 |
| Glyma.10G163200 | ANXUR2 | 2589 | 862 | 5.67 | 95.95 |
| Glyma.19G245800 | ANXUR3 | 2601 | 866 | 5.31 | 95.71 |
| Glyma.20G225800 | ANXUR4 | 2532 | 843 | 5.8 | 93.73 |
| Glyma.12G235900 | BUPS1 | 2637 | 878 | 5.77 | 96.33 |
| Glyma.13G201400 | BUPS2 | 2610 | 869 | 5.85 | 95.29 |
| Glyma.17G102600 | CAP1 | 2586 | 861 | 6.55 | 95.69 |
| Glyma.09G273300 | FERONIA1 | 2691 | 896 | 5.64 | 98.07 |
| Glyma.18G215800 | FERONIA2 | 2685 | 894 | 5.66 | 97.76 |
| Glyma.12G074600 | HERKULES1A | 2514 | 837 | 5.86 | 92.74 |
| Glyma.15G042900 | HERKULES1B | 2226 | 741 | 7.93 | 81.93 |
| Glyma.U033500 | HERKULES1C | 2436 | 811 | 6.5 | 89.91 |
| Glyma.09G024700 | HERKULES2 | 2559 | 852 | 5.59 | 93.59 |
| Glyma.02G121900 | MEDOS1A | 2463 | 820 | 8.23 | 92.15 |
| Glyma.02G122000 | MEDOS1B | 1944 | 647 | 5.81 | 72.73 |
| Glyma.02G196000 | MEDOS1C | 2481 | 826 | 5.83 | 93.46 |
| Glyma.08G248900 | MEDOS2A | 2529 | 842 | 6.25 | 92.88 |
| Glyma.08G249200 | MEDOS2B | 2616 | 871 | 6.24 | 96.62 |
| Glyma.08G249400 | MEDOS2C | 2373 | 790 | 6.02 | 88.62 |
| Glyma.13G054400 | MEDOS3A | 2691 | 896 | 6.11 | 99.53 |
| Glyma.13G053800 | MEDOS3B | 2109 | 702 | 6.44 | 77.97 |
| Glyma.13G053700 | MEDOS3C | 2460 | 819 | 5.63 | 91.47 |
| Glyma.13G053600 | MEDOS3D | 2685 | 894 | 5.9 | 99.35 |
| Glyma.13G054200 | MEDOS3E | 2364 | 787 | 8.48 | 88.74 |
| Glyma.13G054300 | MEDOS3F | 2535 | 844 | 5.98 | 94.104 |
| Glyma.18G269900 | MEDOS4A | 2610 | 869 | 6.26 | 97.24 |
| Glyma.18G270100 | MEDOS4B | 2607 | 868 | 6.09 | 97.39 |
| Glyma.18G270600 | MEDOS4C | 3372 | 1123 | 5.77 | 124.54 |
| Glyma.18G270700 | MEDOS4D | 2574 | 857 | 6.25 | 95.77 |
| Glyma.18G270900 | MEDOS4E | 2628 | 875 | 5.82 | 97.45 |
| Glyma.18G271000 | MEDOS4F | 2592 | 863 | 5.9 | 96.86 |
| Glyma.18G271100 | MEDOS4G | 2652 | 883 | 6.02 | 98.05 |
| Glyma.18G270800 | MEDOS4I | 2730 | 909 | 5.98 | 102.67 |
| Glyma.18G271200 | MEDOS4J | 2550 | 849 | 5.83 | 95.14 |
| Glyma.19G033100 | MEDOS5A | 3561 | 1186 | 6.49 | 133.95 |
| Glyma.U027000 | MEDOS5B | 2364 | 787 | 8.48 | 88.76 |
| Glyma.U027100 | MEDOS5C | 2460 | 819 | 5.75 | 91.75 |
| Glyma.12G148200 | THESEUS1 | 2541 | 846 | 5.68 | 93.19 |
| Glyma.12G220400 | THESEUS2 | 2070 | 689 | 6.44 | 75.72 |
| Glyma.05G099900 | CAD1 | 2382 | 793 | 6.3 | 88.42 |
| Glyma.05G100000 | CAD2 | 2517 | 838 | 5.58 | 94.02 |
| Glyma.09G133000 | CAD3 | 2457 | 818 | 7.04 | 99.92 |
| Glyma.10G231500 | CAD4 | 2481 | 826 | 7.94 | 91.86 |
| Glyma.16G179600 | CAD5 | 2322 | 773 | 8.81 | 86.06 |
| Glyma.17G166200 | CAD6 | 2523 | 840 | 5.8 | 93.93 |
| Glyma.20G162300 | CAD7 | 2523 | 840 | 8.17 | 93.19 |
| A. thaliana | |||||
| AT3G04690 | ANX1 | 2837 | 895 | 6.47 | 98.16 |
| AT5G28680 | ANX2 | 2577 | 850 | 6.54 | 94.06 |
| AT4G39110 | BUPS1 | 2637 | 858 | 5.76 | 94.31 |
| AT2G21480 | BUPS2 | 2616 | 873 | 5.66 | 97.18 |
| AT5G61350 | CAP | 2529 | 880 | 5.92 | 97.96 |
| AT2G39360 | CRV | 2683 | 815 | 6.13 | 91.33 |
| AT3G51550 | FER | 3298 | 830 | 5.82 | 91.48 |
| AT5G59700 | HERK/ANJ | 3041 | 849 | 5.76 | 93.96 |
| AT3G46290 | HERK1 | 3158 | 855 | 5.91 | 93.31 |
| AT1G30570 | HERK2 | 2550 | 842 | 6.16 | 92.7 |
| AT5G39000 | MEDO2 | 2622 | 878 | 5.75 | 96.52 |
| AT5G38990 | MEDOS1 | 2785 | 871 | 5.51 | 95.95 |
| AT5G39020 | MEDOS3 | 2442 | 829 | 6.5 | 91.97 |
| AT5G39030 | MEDOS4 | 2421 | 824 | 5.65 | 91.84 |
| AT5G54380 | THE1 | 2789 | 834 | 5.7 | 93.39 |
| AT5G24010 | CAD1 | 2821 | 813 | 7.6 | 90.64 |
| AT2G23200 | CAD2 | 2633 | 806 | 5.97 | 90.68 |
| L. japonicus | |||||
| Lj1g3v4996200 | ANXUR | 2592 | 863 | 5.46 | 95.4 |
| Lj0g3v0115159 | BUPS | 2643 | 880 | 5.93 | 96.29 |
| Lj3g3v3639930 | CURVY1 | 2472 | 823 | 6.08 | 90.91 |
| Lj3g3v3639940 | CURVY2 | 2121 | 706 | 5.98 | 77.77 |
| Lj1g3v2533770 | FERONIA | 2094 | 697 | 5.96 | 75.96 |
| Lj0g3v0249939 | HERKULES1A | 2514 | 837 | 5.41 | 91.74 |
| Lj3g3v3132890 | HERKULES1B | 1848 | 615 | 6.61 | 67.66 |
| Lj6g3v1641160 | HERKULES2A | 2532 | 843 | 5.77 | 92.66 |
| Lj6g3v1641170 | HERKULES2B | 2532 | 843 | 5.77 | 92.66 |
| Lj2g3v1102970 | MEDOS1 | 2676 | 891 | 6.75 | 97.76 |
| Lj2g3v1226730 | MEDOS2 | 1542 | 513 | 7.59 | 58.1 |
| Lj2g3v1226740 | MEDOS3 | 2466 | 821 | 6.93 | 92.49 |
| Lj2g3v1226750 | MEDOS4 | 2277 | 758 | 5.49 | 84.89 |
| Lj0g3v0346559 | THESEUS | 2535 | 844 | 5.54 | 92.59 |
| Lj0g3v0151929 | CAD1 | 1554 | 517 | 8.87 | 57.31 |
| Lj2g3v0322770 | CAD2 | 2493 | 830 | 6.46 | 92.25 |
| Lj2g3v1902230 | CAD3 | 2169 | 722 | 8.75 | 80.67 |
| Lj5g3v1988700 | CAD4 | 2535 | 844 | 6.98 | 94.3 |
| M. truncatula | |||||
| Medtr1g080740 | ANX1 | 2607 | 868 | 6.15 | 96.86 |
| Medtr7g115300 | ANX2 | 2619 | 872 | 5.32 | 97.04 |
| Medtr8g037700 | BUPS1 | 2406 | 801 | 5.94 | 96.47 |
| Medtr4g109010 | CAP1 | 3459 | 1152 | 6.5 | 129.96 |
| Medtr4g111925 | FER1 | 2106 | 701 | 6.5 | 76.58 |
| Medtr7g073660 | FER2 | 2700 | 899 | 5.89 | 97.98 |
| Medtr4g061930 | HERK1A | 2523 | 840 | 5.82 | 93.13 |
| Medtr2g096160 | HERK1B | 2544 | 847 | 5.57 | 92.93 |
| Medtr4g061833 | HERK1C | 2523 | 840 | 5.82 | 93.13 |
| Medtr2g030310 | HERK2 | 2628 | 875 | 5.89 | 96.34 |
| Medtr6g015805 | MEDOS1A | 2703 | 900 | 6.8 | 100.32 |
| Medtr5g047120 | MEDOS2A | 2430 | 809 | 7.09 | 92.07 |
| Medtr5g047070 | MEDOS2B | 1707 | 568 | 6.23 | 64.53 |
| Medtr7g015390 | MEDOS3A | 2670 | 889 | 6.25 | 101.42 |
| Medtr7g015550 | MEDOS3B | 2667 | 888 | 5.92 | 100.81 |
| Medtr7g015670 | MEDOS3C | 2679 | 892 | 5.75 | 101.65 |
| Medtr7g015510 | MEDOS3D | 2667 | 888 | 5.84 | 100.38 |
| Medtr7g015240 | MEDOS3E | 2529 | 842 | 6.16 | 96.25 |
| Medtr7g015280 | MEDOS3F | 2577 | 858 | 6.44 | 97.98 |
| Medtr7g015420 | MEDOS3G | 3417 | 1138 | 6.02 | 130.78 |
| Medtr4g052290 | MEDOS3H | 2658 | 885 | 7.8 | 101.29 |
| Medtr7g015250 | MEDOS3I | 2733 | 910 | 6.16 | 103.04 |
| Medtr7g015310 | MEDOS3J | 2622 | 873 | 6.08 | 98.98 |
| Medtr7g015230 | MEDOS3K | 2652 | 883 | 6.91 | 100.7 |
| Medtr7g015320 | MEDOS3L | 2637 | 878 | 7.58 | 99.19 |
| Medtr7g015620 | MEDOS3M | 2064 | 687 | 8.81 | 78.9 |
| Medtr5g047060 | MEDOS4A | 2502 | 833 | 5.21 | 94.17 |
| Medtr5g047110 | MEDOS4B | 2454 | 817 | 5.5 | 92.57 |
| Medtr6g048090 | OG1 | 2385 | 794 | 6.84 | 88.69 |
| Medtr1g100110 | OG2 | 2445 | 814 | 6.57 | 91.66 |
| Medtr2g080220 | THE1 | 2532 | 843 | 5.61 | 92.73 |
| Medtr4g095042 | CAD1 | 2556 | 851 | 5.85 | 94.87 |
| Medtr4g095012 | CAD2 | 2460 | 819 | 6.12 | 91.27 |
| Medtr4g095032 | CAD3 | 2361 | 786 | 5.82 | 87.7 |
| Medtr1g040073 | CAD4 | 2256 | 751 | 6.23 | 83.97 |
| Medtr8g467150 | CAD5 | 2277 | 758 | 7.33 | 85.16 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solis-Miranda, J.; Fonseca-García, C.; Nava, N.; Pacheco, R.; Quinto, C. Genome-Wide Identification of the CrRLK1L Subfamily and Comparative Analysis of Its Role in the Legume-Rhizobia Symbiosis. Genes 2020, 11, 793. https://doi.org/10.3390/genes11070793
Solis-Miranda J, Fonseca-García C, Nava N, Pacheco R, Quinto C. Genome-Wide Identification of the CrRLK1L Subfamily and Comparative Analysis of Its Role in the Legume-Rhizobia Symbiosis. Genes. 2020; 11(7):793. https://doi.org/10.3390/genes11070793
Chicago/Turabian StyleSolis-Miranda, Jorge, Citlali Fonseca-García, Noreide Nava, Ronal Pacheco, and Carmen Quinto. 2020. "Genome-Wide Identification of the CrRLK1L Subfamily and Comparative Analysis of Its Role in the Legume-Rhizobia Symbiosis" Genes 11, no. 7: 793. https://doi.org/10.3390/genes11070793
APA StyleSolis-Miranda, J., Fonseca-García, C., Nava, N., Pacheco, R., & Quinto, C. (2020). Genome-Wide Identification of the CrRLK1L Subfamily and Comparative Analysis of Its Role in the Legume-Rhizobia Symbiosis. Genes, 11(7), 793. https://doi.org/10.3390/genes11070793