Tackling the Molecular Drug Sensitivity in the Sea Louse Caligus rogercresseyi Based on mRNA and lncRNA Interactions



Abstract
:1. Introduction
2. Materials and Methods
2.1. Lice Samples and Culture in Laboratory-Controlled Conditions
2.2. Bioassays for Transcriptome Sequencing
2.3. High-Throughput Sequencing from Lice Exposed to Delousing Drugs
2.4. Transcriptome Analyses of Coding Genes in C. rogercresseyi
2.5. LncRNAs Identification and Characterization from Lice Transcriptome
2.6. Validation of Coding Genes and lncRNAs Transcription Levels
3. Results
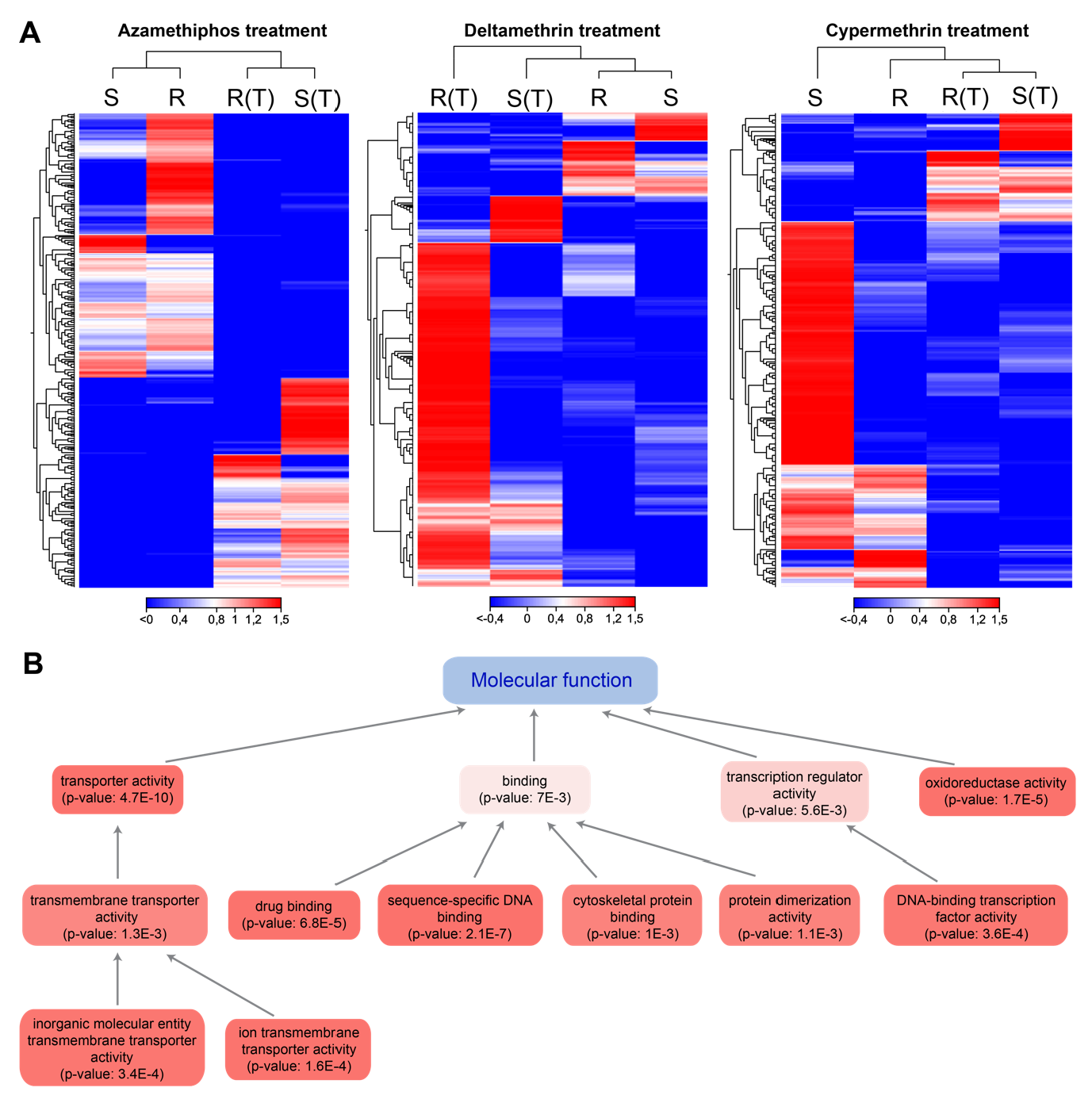
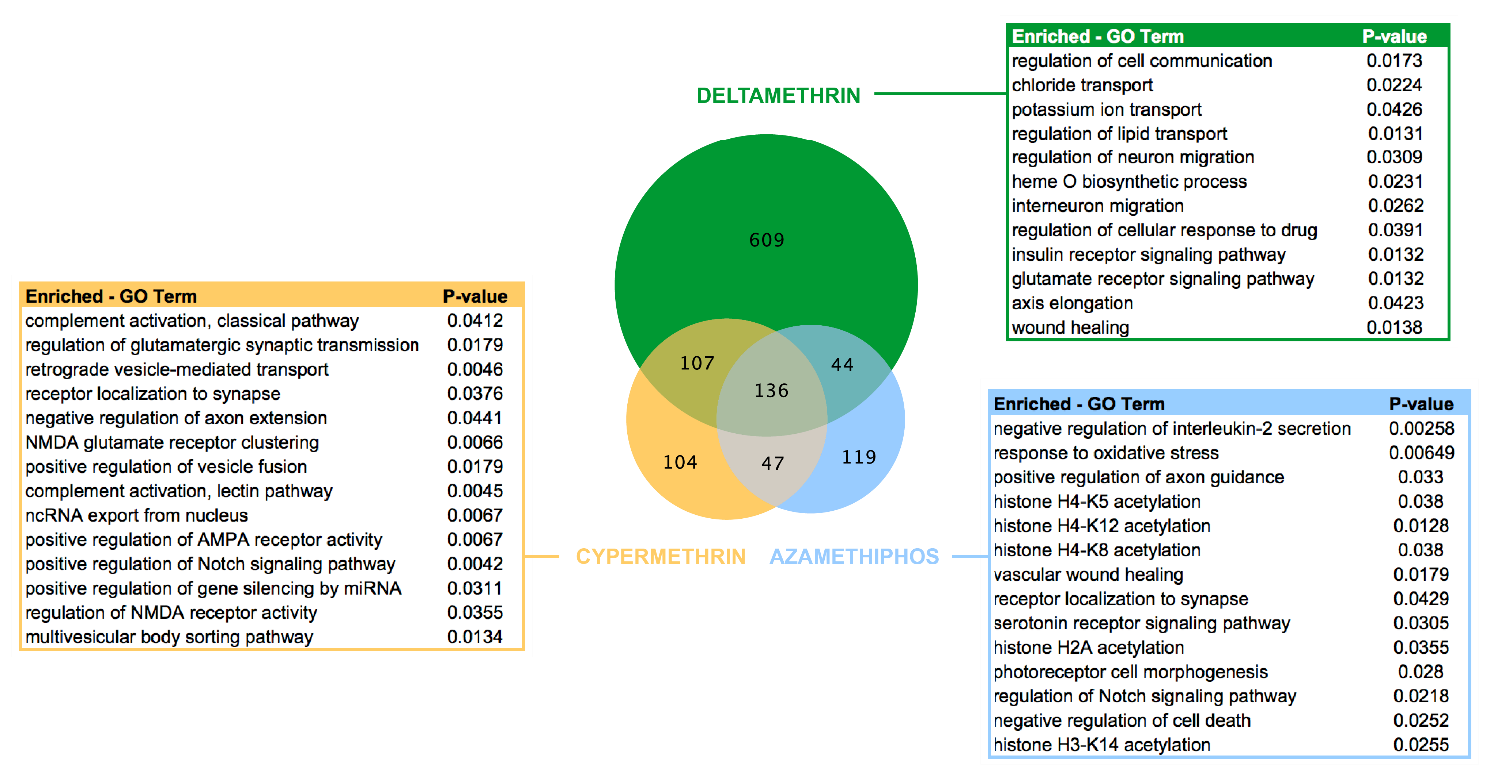
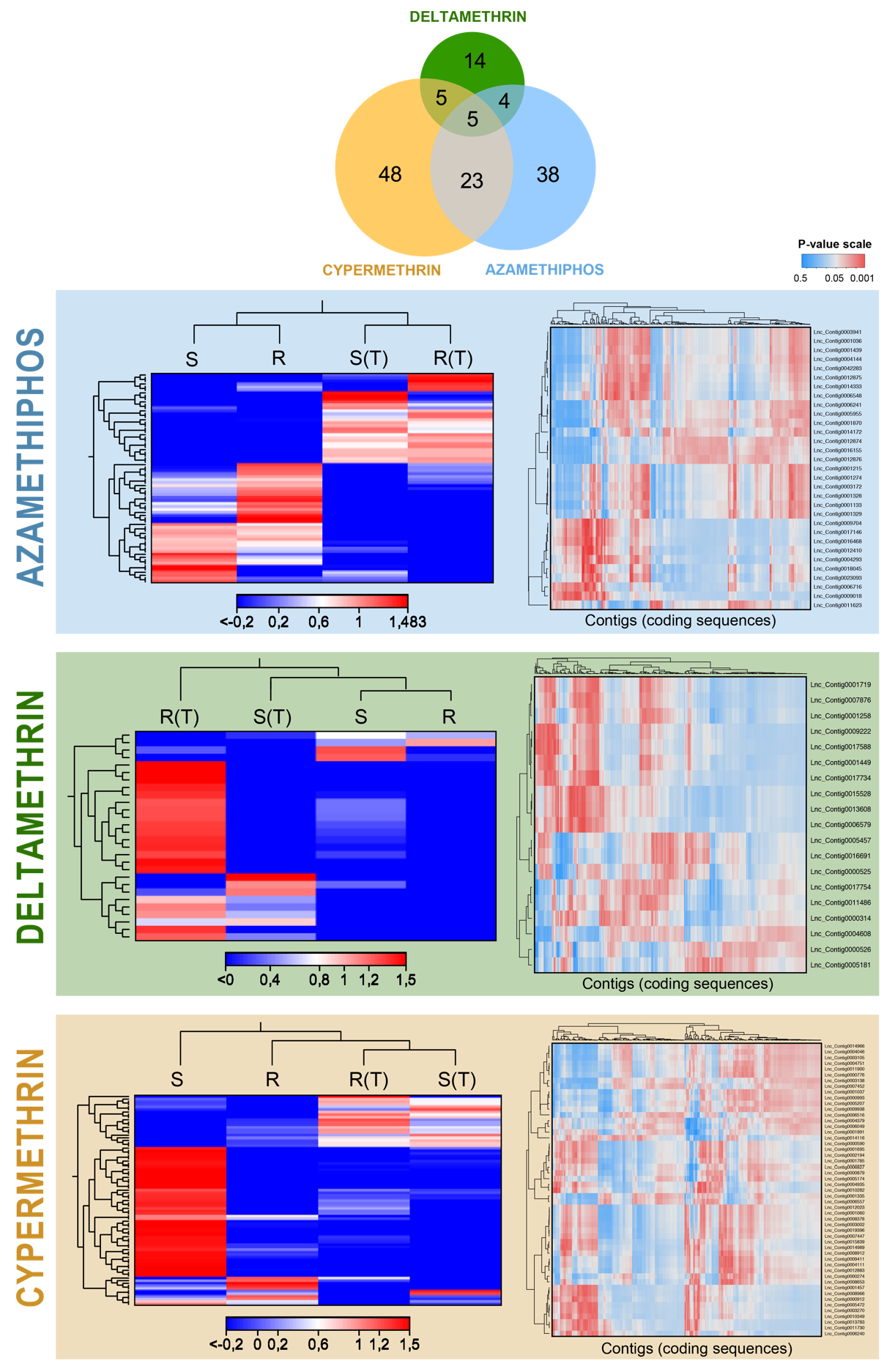
3.1. Coding Transcriptomes Associated with Drug Exposure
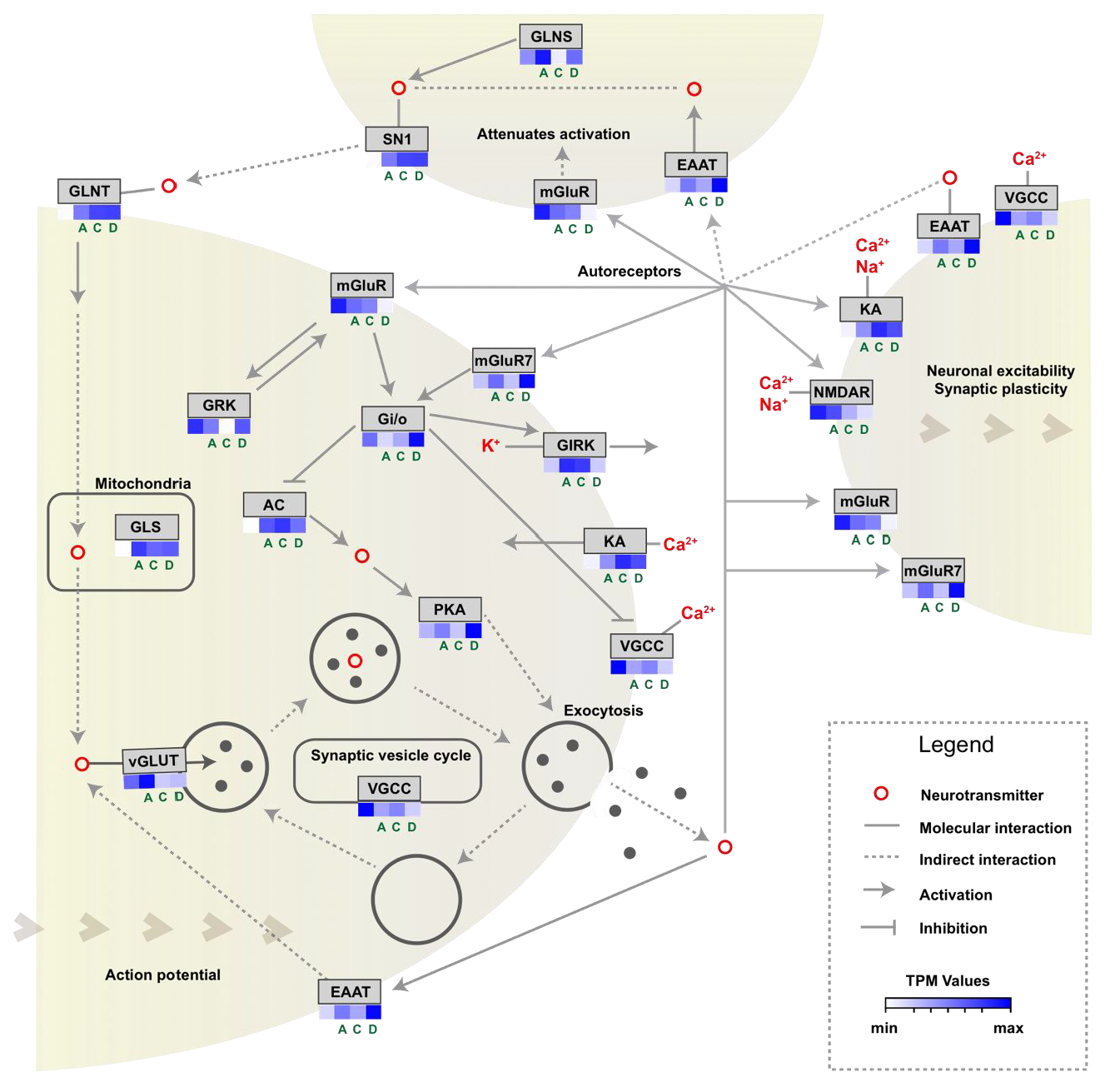
3.2. Glutamatergic Synapse Pathway
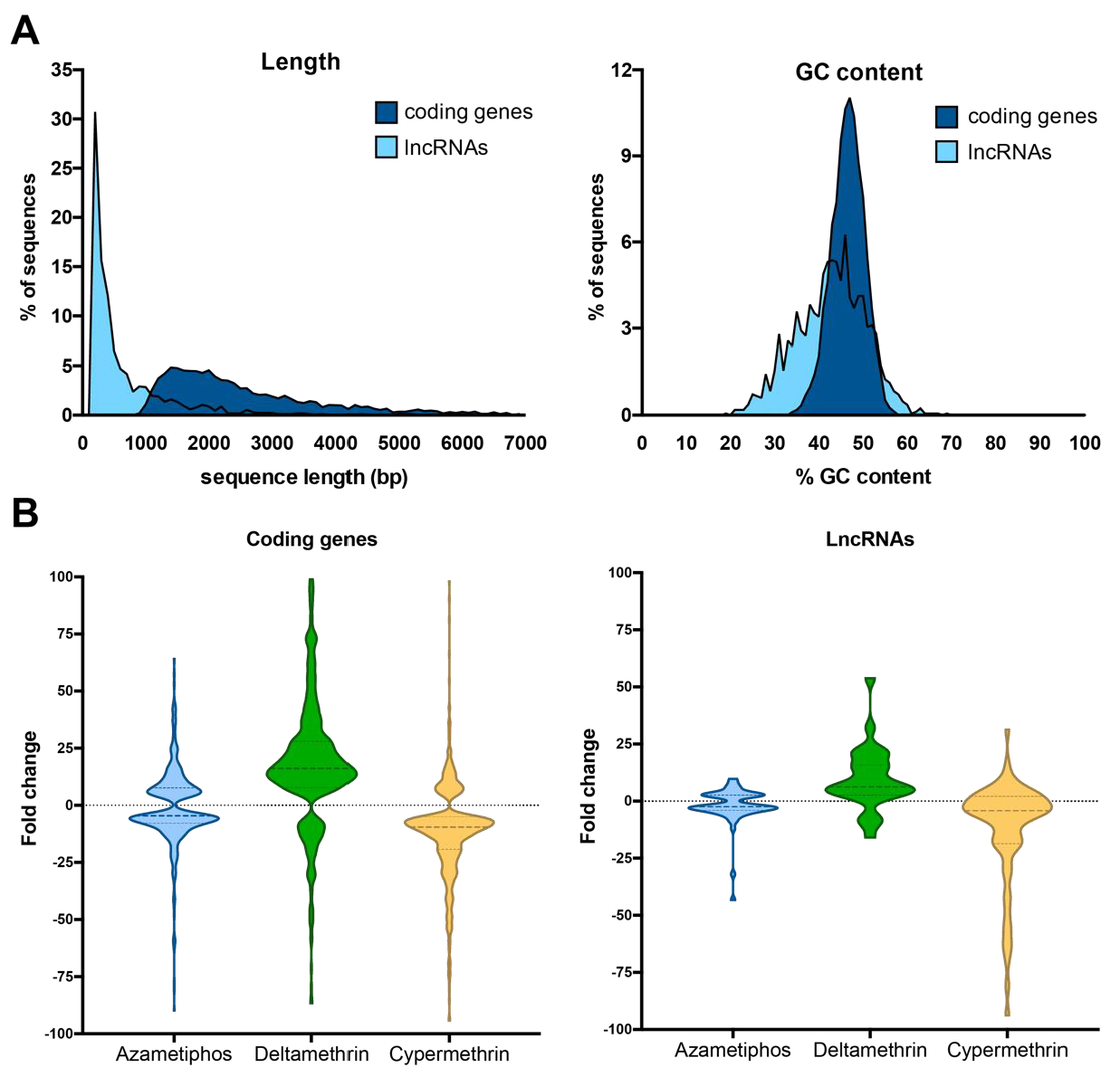
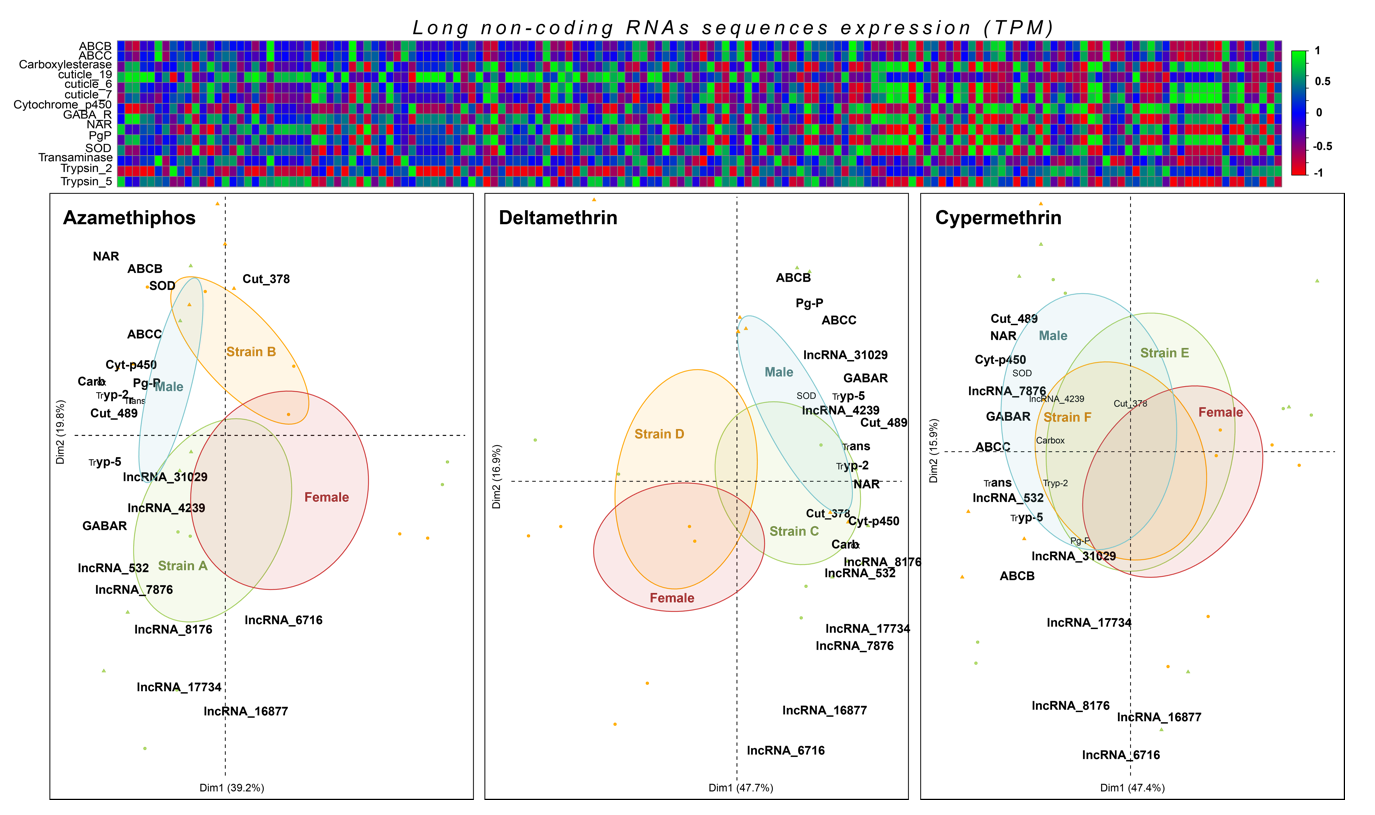
3.3. LncRNA Characterization and Expression Patterns
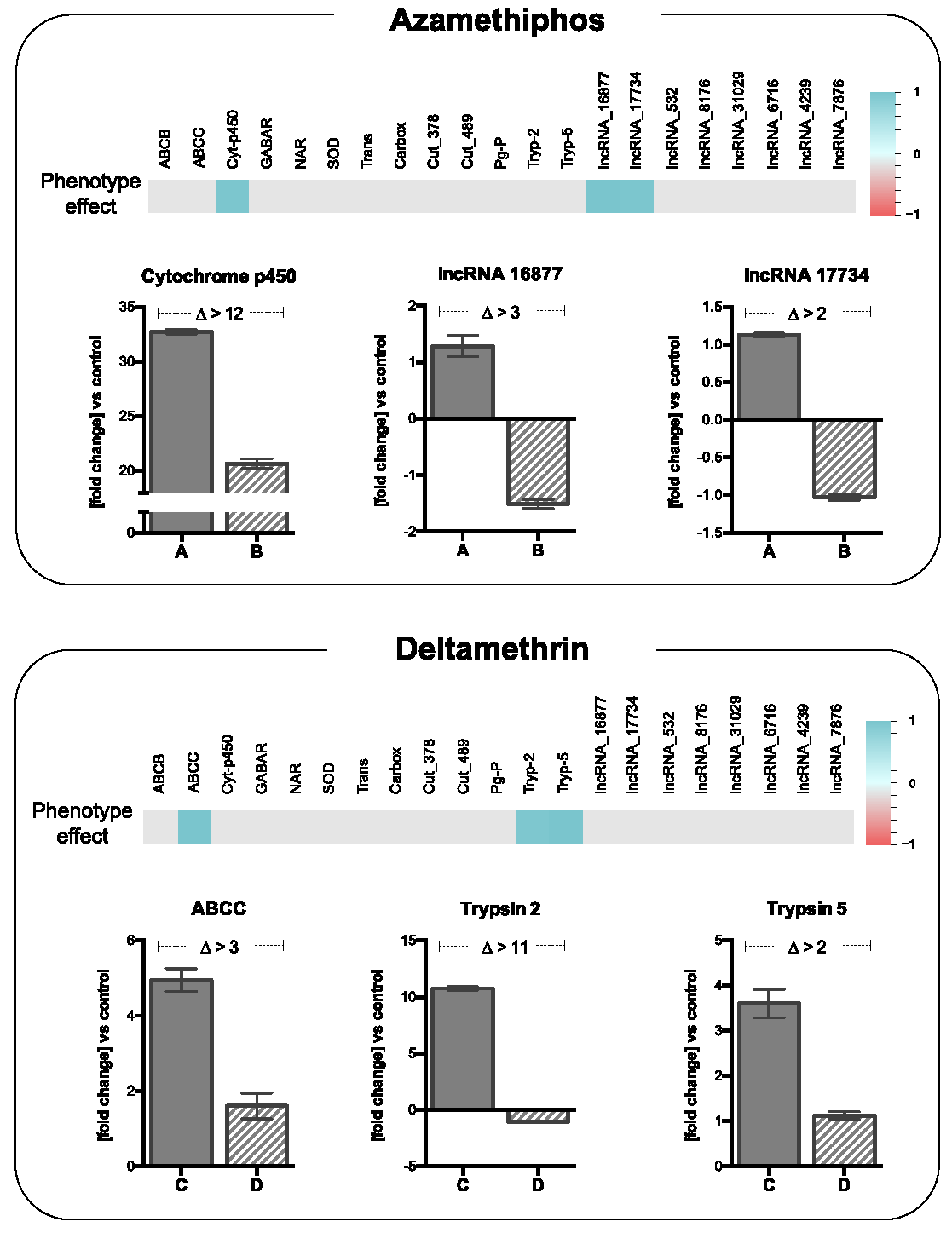
3.4. Validation of Specific Transcripts Associated with Delousing Drug Tolerance/Susceptibility
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Koboldt, D.C.; Steinberg, K.M.; Larson, D.E.; Wilson, R.K.; Mardis, E.R. The next-generation sequencing revolution and its impact on genomics. Cell 2013, 155, 27–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, S.; McPherson, J.D.; McCombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333. [Google Scholar] [CrossRef] [PubMed]
- Necsulea, A.; Kaessmann, H. Evolutionary dynamics of coding and non-coding transcriptomes. Nat. Rev. Genet. 2014, 15, 734–748. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R. Non–coding RNA genes and the modern RNA world. Nat. Rev. Genet. 2001, 2, 919. [Google Scholar] [CrossRef]
- Chen, L.-L.; Carmichael, G.G. Decoding the function of nuclear long non-coding RNAs. Curr. Opin. Cell Biol. 2010, 22, 357–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155. [Google Scholar] [CrossRef]
- Wang, Z. Diverse roles of regulatory non-coding RNAs. J. Mol. Cell Biol. 2018, 10, 91–92. [Google Scholar] [CrossRef] [Green Version]
- Etebari, K.; Furlong, M.J.; Asgari, S. Genome wide discovery of long intergenic non-coding RNAs in Diamondback moth (Plutella xylostella) and their expression in insecticide resistant strains. Sci. Rep. 2015, 5, 14642. [Google Scholar] [CrossRef]
- Kumar, D.; Hu, X.; Guo, R.; Xue, R.; Cao, G.; Gong, C. Long noncoding RNA: Disclosing new horizon in the molecular world of insects. In Trends in Insect Molecular Biology and Biotechnology; Kumar, D., Gong, C., Eds.; Springer International Publishing: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Valenzuela-Miranda, D.; Etebari, K.; Asgari, S.; Gallardo-Escárate, C. Long noncoding RNAs: Unexplored players in the drug response of the sea louse Caligus rogercresseyi. Agri Gene 2017, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gallardo-Escárate, C.; Arriagada, G.; Carrera, C.; Gonçalves, A.T.; Nuñez-Acuña, G.; Valenzuela-Miranda, D.; Valenzuela-Muñoz, V. The race between host and sea lice in the Chilean salmon farming: A genomic approach. Rev. Aquac. 2019, 11, 325–339. [Google Scholar] [CrossRef]
- Niklitschek, E.J.; Soto, D.; Lafon, A.; Molinet, C.; Toledo, P. Southward expansion of the Chilean salmon industry in the Patagonian Fjords: Main environmental challenges. Rev. Aquac. 2013, 5, 172–195. [Google Scholar] [CrossRef]
- Chávez, C.; Dresdner, J.; Figueroa, Y.; Quiroga, M. Main issues and challenges for sustainable development of salmon farming in Chile: A socio-economic perspective. Rev. Aquac. 2019, 11, 403–421. [Google Scholar] [CrossRef]
- Bravo, S.; Silva, M.T.; Agusti, C.; Sambra, K.; Horsberg, T.E. The effect of chemotherapeutic drugs used to control sea lice on the hatching viability of egg strings from Caligus rogercresseyi. Aquaculture 2015, 443, 77–83. [Google Scholar] [CrossRef]
- Myhre Jensen, E.; Sevatdal, S.; Bakke, M.J.; Kaur, K.; Horsberg, T.E. A selection study on a laboratory-designed population of salmon lice (Lepeophtheirus salmonis) using organophosphate and pyrethroid pesticides. PLoS ONE 2017, 12, e0178068. [Google Scholar] [CrossRef]
- Fast, M.D. Fish Immune Responses to Parasitic Copepod (namely Sea lice) Infection. Dev. Comp. Immunol. 2014, 43, 300–312. [Google Scholar] [CrossRef]
- Moores, G.D.; Devonshire, A.L.; Denholm, I. A microtitre plate assay for characterizing insensitive acetylcholinesterase genotypes of insecticide-resistant insects. Bull. Entomol. Res. 1988, 78, 537–544. [Google Scholar] [CrossRef]
- Shafer, T.J.; Meyer, D.A. Effects of pyrethroids on voltage-sensitive calcium channels: A critical evaluation of strengths, weaknesses, data needs, and relationship to assessment of cumulative neurotoxicity. Toxicol. Appl. Pharmacol. 2004, 196, 303–318. [Google Scholar] [CrossRef]
- Kaur, K.; Besnier, F.; Glover, K.A.; Nilsen, F.; Aspehaug, V.T.; Fjørtoft, H.B.; Horsberg, T.E. The mechanism (Phe362Tyr mutation) behind resistance in Lepeophtheirus salmonis pre-dates organophosphate use in salmon farming. Sci. Rep. 2017, 7, 12349. [Google Scholar] [CrossRef]
- Bakke, M.J.; Agusti, C.; Bruusgaard, J.C.; Sundaram, A.Y.M.; Horsberg, T.E. Deltamethrin resistance in the salmon louse, Lepeophtheirus salmonis (Krøyer): Maternal inheritance and reduced apoptosis. Sci. Rep. 2018, 8, 8450. [Google Scholar] [CrossRef]
- Chávez-Mardones, J.; Gallardo-Escárate, C. Next-Generation Transcriptome Profiling of the Salmon Louse Caligus rogercresseyi Exposed to Deltamethrin (AlphaMax™): Discovery of Relevant Genes and Sex-Related Differences. Mar. Biotechnol. 2015, 1–18. (in press). [Google Scholar] [CrossRef]
- Chavez-Mardones, J.; Gallardo-Escárate, C. Deltamethrin (AlphaMax™) reveals modulation of genes related to oxidative stress in the ectoparasite Caligus rogercresseyi: Implications on delousing drug effectiveness. Aquaculture 2014, 433, 421–429. [Google Scholar] [CrossRef]
- Valenzuela-Muñoz, V.; Nuñez-Acuña, G.; Gallardo-Escárate, C. Molecular characterization and transcription analysis of P-glycoprotein gene from the salmon louse Caligus rogercresseyi. J. Aquac. Res. Dev. 2014, 5, 236. [Google Scholar]
- Chávez-Mardones, J.; Valenzuela-Muños, V.; Gallardo-Escárate, C. In silico transcriptome analysis of cuticle-related genes associated with delousing drug responses in the sea louse Caligus rogercresseyi. Aquaculture 2016, 450, 123–135. [Google Scholar] [CrossRef]
- Valenzuela-Muñoz, V.; Chavez-Mardones, J.; Gallardo-Escárate, C. RNA-seq analysis evidences multiple gene responses in Caligus rogercresseyi exposed to the anti-salmon lice drug azamethiphos. Aquaculture 2015, 446, 156–166. [Google Scholar] [CrossRef]
- Valenzuela-Muñoz, V.; Sturm, A.; Gallardo-Escárate, C. Transcriptomic insights on the ABC transporter gene family in the salmon louse Caligus rogercresseyi. Parasites Vectors 2015, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Helgesen, K.; Bravo, S.; Sevatdal, S.; Mendoza, J.; Horsberg, T. Deltamethrin resistance in the sea louse Caligus rogercresseyi (Boxhall and Bravo) in Chile: Bioassay results and usage data for antiparasitic agents with references to Norwegian conditions. J. Fish Dis. 2014, 37, 877–890. [Google Scholar] [CrossRef]
- Agusti, C.; Bravo, S.; Contreras, G.; Bakke, M.J.; Helgesen, K.O.; Winkler, C.; Silva, M.T.; Mendoza, J.; Horsberg, T.E. Sensitivity assessment of Caligus rogercresseyi to anti-louse chemicals in relation to treatment efficacy in Chilean salmonid farms. Aquaculture 2016, 458, 195–205. [Google Scholar] [CrossRef]
- Marín, S.; Ibarra, R.; Medina, M.; Jansen, P. Sensitivity of Caligus rogercresseyi (Boxshall and Bravo 2000) to pyrethroids and azamethiphos measured using bioassay tests—A large scale spatial study. Prev. Vet. Med. 2015, (in press). [Google Scholar]
- Consortium, S. Sea Lice Resistance to Chemotherapeutants: A Handbook in Resistance Management; SEARCH: Sacramento, CA, USA, 2006. [Google Scholar]
- Helgesen, K.O.; Horsberg, T.E. Single-dose field bioassay for sensitivity testing in sea lice, Lepeophtheirus salmonis: Development of a rapid diagnostic tool. J. Fish Dis. 2013, 36, 261–272. [Google Scholar] [CrossRef]
- Agusti-ridaura, C.; Dondrup, M.; Horsberg, T.E.; Leong, J.S.; Koop, B.F.; Bravo, S.; Mendoza, J.; Kaur, K. Caligus rogercresseyi acetylcholinesterase types and variants: A potential marker for organophosphate resistance. Parasites Vectors 2018, 11, 570. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Escárate, C.; Valenzuela-Muñoz, V.; Núñez-Acuña, G. RNA-Seq analysis using de novo transcriptome assembly as a reference for the salmon louse Caligus rogercresseyi. PLoS ONE 2014, 9, e92239. [Google Scholar] [CrossRef] [PubMed]
- Bairoch, A.; Apweiler, R. The SWISS-PROT protein sequence database and its supplement TrEMBL in 2000. Nucleic Acids Res. 2000, 28, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Park, H.J.; Dasari, S.; Wang, S.; Kocher, J.-P.; Li, W. CPAT: Coding-Potential Assessment Tool using an alignment-free logistic regression model. Nucleic Acids Res. 2013, 41, e74. [Google Scholar] [CrossRef]
- Kong, L.; Zhang, Y.; Ye, Z.-Q.; Liu, X.-Q.; Zhao, S.-Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, W345–W349. [Google Scholar] [CrossRef]
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: http://www.R-project.org/ (accessed on 1 April 2020).
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix, R Package Version 0.84. 2017. Available online: https://github.com/taiyun/corrplot (accessed on 1 April 2020).
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper–Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Wu, H.-W.; Tian, H.-S.; Wu, G.-L.; Langdon, G.; Kurtis, J.; Shen, B.; Ma, L.; Li, X.-L.; Gu, Y.; Hu, X.-B. Culex pipiens pallens: Identification of genes differentially expressed in deltamethrin-resistant and-susceptible strains. Pestic. Biochem. Physiol. 2004, 79, 75–83. [Google Scholar] [CrossRef]
- Imamura, L.; Yasuda, M.; Kuramitsu, K.; Hara, D.; Tabuchi, A.; Tsuda, M. Deltamethrin, a pyrethroid insecticide, is a potent inducer for the activity-dependent gene expression of brain-derived neurotrophic factor in neurons. J. Pharmacol. Exp. Ther. 2006, 316, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Olsvik, P.A.; Ørnsrud, R.; Lunestad, B.T.; Steine, N.; Fredriksen, B.N. Transcriptional responses in Atlantic salmon (Salmo salar) exposed to deltamethrin, alone or in combination with azamethiphos. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014, 162, 23–33. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, Y.; Jiang, Y.; Zhu, F.; Zeng, L.; Wang, Y.; Lei, X.; Yao, Y.; Hou, Y.; Xu, L. Transcriptional responses in the hepatopancreas of Eriocheir sinensis exposed to deltamethrin. PLoS ONE 2017, 12, e0184581. [Google Scholar] [CrossRef] [PubMed]
- Vester, A.; Caudle, W.M. The synapse as a central target for neurodevelopmental susceptibility to pesticides. Toxics 2016, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, T.; Momiyama, A. Different types of calcium channels mediate central synaptic transmission. Nature 1993, 366, 156. [Google Scholar] [CrossRef] [PubMed]
- Collingridge, G.; Bliss, T. NMDA receptors-their role in long-term potentiation. Trends Neurosci. 1987, 10, 288–293. [Google Scholar] [CrossRef]
- Bleakman, D.; Lodge, D. Neuropharmacology of AMPA and kainate receptors. Neuropharmacology 1998. [Google Scholar] [CrossRef]
- Kojika, S.; Griffin, J.D. Notch receptors and hematopoiesis. Exp. Hematol. 2001, 29, 1041–1052. [Google Scholar] [CrossRef]
- Rao, V.R.; Finkbeiner, S. NMDA and AMPA receptors: Old channels, new tricks. Trends Neurosci. 2007, 30, 284–291. [Google Scholar] [CrossRef]
- Chávez-Mardones, J.; Boltaña, S.; Valenzuela-Muñoz, V.; Gallardo-Escárate, C. Evidence for the induction of key components of the Notch signaling pathway via deltamethrin and azamethiphos treatment in the sea louse Caligus rogercresseyi. Comp. Biochem. Phys. D. 2015, (in press). [Google Scholar]
- Dickson, B.J. Molecular mechanisms of axon guidance. Science 2002, 298, 1959–1964. [Google Scholar] [CrossRef] [Green Version]
- Xing, H.; Wang, C.; Wu, H.; Chen, D.; Li, S.; Xu, S. Effects of atrazine and chlorpyrifos on DNA methylation in the brain and gonad of the common carp. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2015, 168, 11–19. [Google Scholar] [CrossRef]
- Lauschke, V.M.; Barragan, I.; Ingelman-Sundberg, M. Pharmacoepigenetics and Toxicoepigenetics: Novel Mechanistic Insights and Therapeutic Opportunities. Ann. Rev. Pharmacol. Toxicol. 2018, 58, 161–185. [Google Scholar] [CrossRef] [PubMed]
- Englert, N.A.; Luo, G.; Goldstein, J.A.; Surapureddi, S. Epigenetic Modification of Histone 3 Lysine 27 mediator subunit MED25 is required for the dissociation of polycomb repressive complex 2 from the promoter of cytochrome p450 2C9. J. Biol. Chem. 2015, 290, 2264–2278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aaen, S.M.; Helgesen, K.O.; Bakke, M.J.; Kaur, K.; Horsberg, T.E. Drug resistance in sea lice: A threat to salmonid aquaculture. Trends Parasitol. 2015, 31, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Kozhemyakin, M.; Rajasekaran, K.; Kapur, J. Central Cholinesterase Inhibition Enhances Glutamatergic Synaptic Transmission. J. Neurophysiol. 2010, 103, 1748–1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greget, R.; Dadak, S.; Barbier, L.; Lauga, F.; Linossier-Pierre, S.; Pernot, F.; Legendre, A.; Ambert, N.; Bouteiller, J.-M.; Dorandeu, F.; et al. Modeling and simulation of organophosphate-induced neurotoxicity: Prediction and validation by experimental studies. Neurotoxicology 2016, 54, 140–152. [Google Scholar] [CrossRef]
- Aroniadou-Anderjaska, V.; Figueiredo, T.H.; Apland, J.P.; Braga, M.F. Targeting the glutamatergic system to counteract organophosphate poisoning: A novel therapeutic strategy. Neurobiol. Dis. 2019. [Google Scholar] [CrossRef]
- Klaassen, C.D.; Amdur, M.O. Casarett and Doull’s Toxicology: The Basic Science of Poisons; McGraw-Hill New York: New York, NY, USA, 2013; Volume 1236. [Google Scholar]
- Núñez-Acuña, G.; Boltaña, S.; Gallardo-Escárate, C. Pesticides drive stochastic changes in the chemoreception and neurotransmission system of marine ectoparasites. Int. J. Mol. Sci. 2016, 17, 700. [Google Scholar] [CrossRef] [Green Version]
- Ffrench-Constant, R.H.; Daborn, P.J.; Goff, G.L. The genetics and genomics of insecticide resistance. Trends Genet. 2004, 20, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Heckel, D.G. Insecticide Resistance After Silent Spring. Science 2012, 337, 1612–1614. [Google Scholar] [CrossRef] [Green Version]
- Riveron, J.M.; Ibrahim, S.S.; Chanda, E.; Mzilahowa, T.; Cuamba, N.; Irving, H.; Barnes, K.G.; Ndula, M.; Wondji, C.S. The highly polymorphic CYP6M7 cytochrome P450 gene partners with the directionally selected CYP6P9a and CYP6P9b genes to expand the pyrethroid resistance front in the malaria vector Anopheles funestus in Africa. BMC Genom. 2014, 15, 817. [Google Scholar] [CrossRef] [Green Version]
- Gong, M.; Shen, B.; Gu, Y.; Tian, H.; Ma, L.; Li, X.; Yang, M.; Hu, Y.; Sun, Y.; Hu, X.; et al. Serine proteinase over-expression in relation to deltamethrin resistance in Culex pipiens pallens. Arch. Biochem. Biophys. 2005, 438, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Anders, F.; Ian, D.; Tor Einar, H.; Martin, S.W. Novel point mutation in the sodium channel gene of pyrethroid-resistant sea lice Lepeophtheirus salmonis (Crustacea: Copepoda). Dis. Aquat. Org. 2005, 65, 129–136. [Google Scholar]
- Sevatdal, S.; Fallang, A.; Ingebrigtsen, K.; Horsberg, T.E. Monooxygenase mediated pyrethroid detoxification in sea lice (Lepeophtheirus salmonis). Pest. Manag. Sci. 2005, 61, 772–778. [Google Scholar] [CrossRef]
- Valenzuela-Miranda, D.; Gallardo-Escárate, C. Caligus rogercresseyi serine proteases: Transcriptomic analysis in response to delousing drugs treatments. Aquaculture 2016, 465, 65–77. [Google Scholar] [CrossRef]
- Carmona-Antoñanzas, G.; Bekaert, M.; Humble, J.L.; Boyd, S.; Roy, W.; Bassett, D.I.; Houston, R.D.; Gharbi, K.; Bron, J.E.; Sturm, A. Maternal inheritance of deltamethrin resistance in the salmon louse Lepeophtheirus salmonis (Krøyer) is associated with unique mtDNA haplotypes. PLoS ONE 2017, 12, e0180625. [Google Scholar] [CrossRef] [PubMed]
- Spencer, A.G.; Jespersen, J.B.; Knorr, M.; Kristensen, M. Selection and Reversion of Azamethiphos-Resistance in a Field Population of the Housefly Musca domestica (Diptera: Muscidae), and the Underlying Biochemical Mechanisms. J. Econ. Entomol. 2000, 93, 1788–1795. [Google Scholar] [CrossRef]
- Kristensen, M. Glutathione S-transferase and insecticide resistance in laboratory strains and field populations of Musca domestica. J. Econ. Entomol. 2005, 98, 1341–1348. [Google Scholar] [CrossRef]
- Kaur, K.; Jansen, P.A.; Aspehaug, V.T.; Horsberg, T.E. Phe362Tyr in AChE: A major factor responsible for azamethiphos resistance in Lepeophtheirus salmonis in Norway. PLoS ONE 2016, 11, e0149264. [Google Scholar] [CrossRef] [Green Version]
- Fjørtoft, H.B.; Besnier, F.; Stene, A.; Nilsen, F.; Bjørn, P.A.; Tveten, A.-K.; Finstad, B.; Aspehaug, V.; Glover, K.A. The Phe362Tyr mutation conveying resistance to organophosphates occurs in high frequencies in salmon lice collected from wild salmon and trout. Sci. Rep. 2017, 7, 14258. [Google Scholar] [CrossRef] [Green Version]
- Helgesen, K.O.; Røyset, K.; Aspehaug, V.; Jansen, P.A. The protective effect of the Phe362Tyr mutation in salmon lice’(Lepeophtheirus salmonis) AChE when exposed to full-scale azamethiphos bath treatments. Aquaculture 2019, 505, 517–522. [Google Scholar] [CrossRef]
- Valenzuela-Miranda, D.; Nuñez-Acuña, G.; Valenzuela-Muñoz, V.; Asgari, S.; Gallardo-Escárate, C. MicroRNA biogenesis pathway from the salmon louse (Caligus rogercresseyi): Emerging role in delousing drug response. Gene 2015, 555, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Escárate, C.; Valenzuela-Muñoz, V.; Núñez-Acuña, G.; Carrera, C.; Gonçalves, A.T.; Valenzuela-Miranda, D.; Benavente, B.P.; Roberts, S. Catching the complexity of salmon-louse interactions. Fish Shellfish Immunol. 2019, 90, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Arriagada, G.; Stryhn, H.; Campistó, J.; Rees, E.; Sanchez, J.; Ibarra, R.; Medina, M.; St-Hilaire, S. Evaluation of the performance of pyrethroids on different life stages of Caligus rogercresseyi in southern Chile. Aquaculture 2014, 426, 231–237. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Drug | Location | Treatment Efficacy | EC50 | 95% CI |
|---|---|---|---|---|---|
| A | Azamethiphos | Aysén | <60% | 13.72 | 6.392–32.6 |
| B | Azamethiphos | Aysén | >92% | 0.4778 | 0.0765–1.275 |
| C | Deltamethrin | Los Lagos | <50% | 3.042 | 1.826–5.285 |
| D | Deltamethrin | Aysén | >80% | 1.424 | 0.634–3.746 |
| E | Cypermethrin | Los Lagos | <40% | 9.709 | 3.155–57.84 |
| F | Cypermethrin | Los Lagos | >80% | 2.178 | 0.972–5.259 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Núñez-Acuña, G.; Sáez-Vera, C.; Valenzuela-Muñoz, V.; Valenzuela-Miranda, D.; Arriagada, G.; Gallardo-Escárate, C. Tackling the Molecular Drug Sensitivity in the Sea Louse Caligus rogercresseyi Based on mRNA and lncRNA Interactions. Genes 2020, 11, 857. https://doi.org/10.3390/genes11080857
Núñez-Acuña G, Sáez-Vera C, Valenzuela-Muñoz V, Valenzuela-Miranda D, Arriagada G, Gallardo-Escárate C. Tackling the Molecular Drug Sensitivity in the Sea Louse Caligus rogercresseyi Based on mRNA and lncRNA Interactions. Genes. 2020; 11(8):857. https://doi.org/10.3390/genes11080857
Chicago/Turabian StyleNúñez-Acuña, Gustavo, Constanza Sáez-Vera, Valentina Valenzuela-Muñoz, Diego Valenzuela-Miranda, Gabriel Arriagada, and Cristian Gallardo-Escárate. 2020. "Tackling the Molecular Drug Sensitivity in the Sea Louse Caligus rogercresseyi Based on mRNA and lncRNA Interactions" Genes 11, no. 8: 857. https://doi.org/10.3390/genes11080857
APA StyleNúñez-Acuña, G., Sáez-Vera, C., Valenzuela-Muñoz, V., Valenzuela-Miranda, D., Arriagada, G., & Gallardo-Escárate, C. (2020). Tackling the Molecular Drug Sensitivity in the Sea Louse Caligus rogercresseyi Based on mRNA and lncRNA Interactions. Genes, 11(8), 857. https://doi.org/10.3390/genes11080857