Sexual Difference in the Optimum Environmental Conditions for Growth and Maturation of the Brown Alga Undaria pinnatifida in the Gametophyte Stage

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation and Culture of Gametophytes
2.2. Measurement of Growth and Maturation of Gametophytes
2.3. Temperature Data
2.4. Experimental Design
2.5. Statistical Analysis
3. Results
3.1. Regional Differences of Undaria pinnatifida
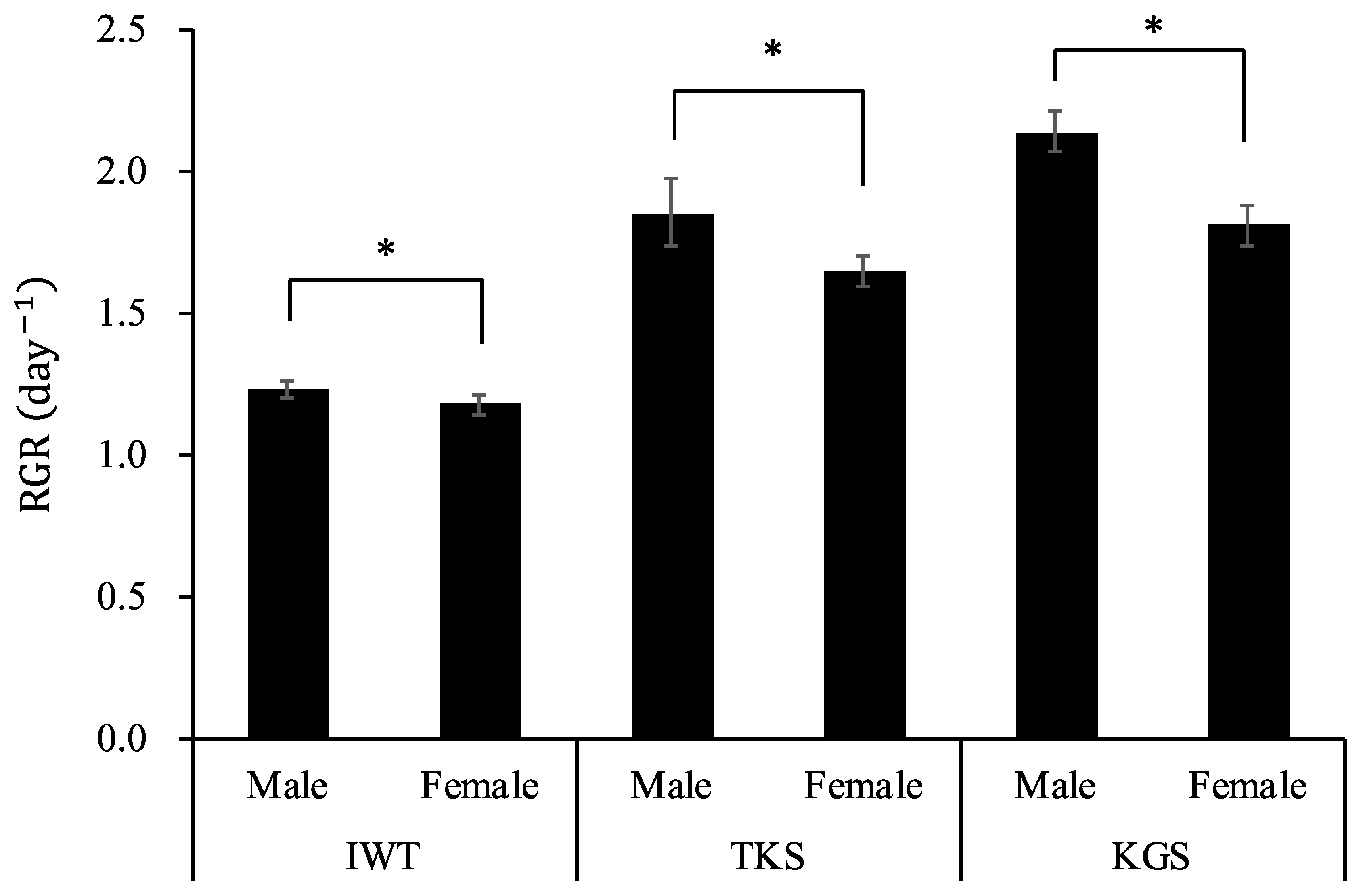
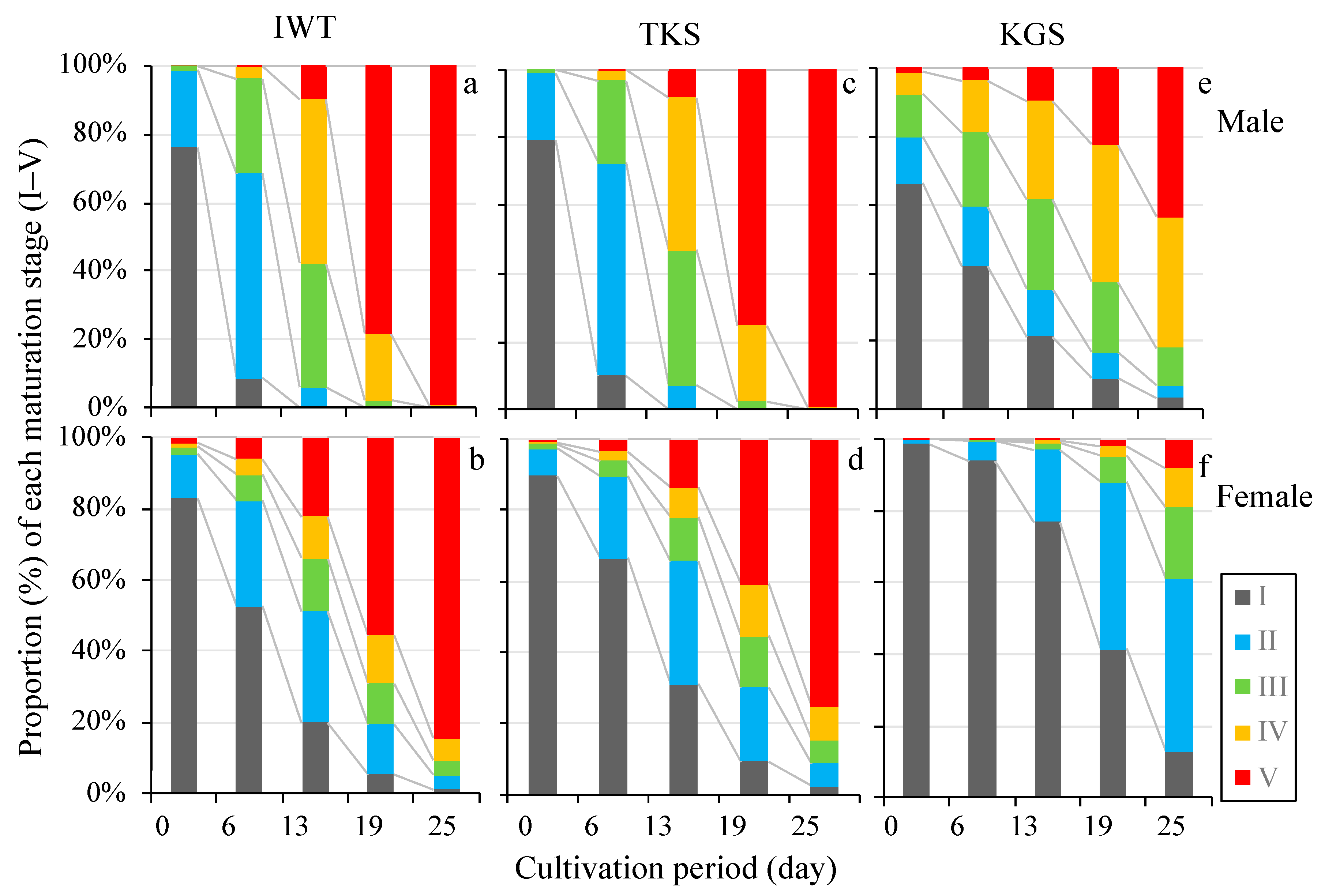
3.2. Growth and Maturation of Gametophytes
3.3. Optimum Conditions for Growth of Gametophytes
3.4. Optimum Conditions for Maturation of Gametophytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rise, W.R. Sex chromosomes and the evolution of sexual dimorphism. Evolution 1984, 38, 735–742. [Google Scholar]
- Umen, J.; Coelho, S. Algal sex determination and the evolution of anisogamy. Annu. Rev. Microbiol. 2019, 73, 267–291. [Google Scholar] [CrossRef] [PubMed]
- Coelho, S.M.; Gueno, J.; Lipinska, A.P.; Cock, J.M.; Umen, J.G. UV chromosomes and haploid sexual systems. Trends Plant Sci. 2018, 23, 794–807. [Google Scholar] [CrossRef] [PubMed]
- Luthringer, R.; Cormier, A.; Ahmed, S.; Peters, A.F.; Cock, J.M.; Coelho, S.M. Sexual dimorphism in the brown algae. Perspect. Phycol. 2014, 1, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, R.; Akiyama, K. Cultivation and utilization of Undaria pinnatifida (wakame) as food. J. Appl. Phycol. 1993, 5, 249–253. [Google Scholar] [CrossRef]
- Kanda, T. On the gametophytes of some Japanese species of Laminariales. Sci. Pap. Inst. Alg. Res. Fac. Sci. Hokkaido Imp. Univ. 1936, 1, 221–260. [Google Scholar]
- Destombe, C.; Oppliger, V. Male gametophyte fragmentation in Laminaria digitata: A life history strategy to enhance reproductive success. Cah. Biol. Mar. 2011, 52, 385. [Google Scholar]
- Dan, A.; Ohno, M.; Matsuoka, M. Changes of the research and development on the resources of Undaria and Laminaria in the culture ground of Tokushima coasts. Bull. Tokushima. Pref. Fish Res. Inst. 2015, 10, 25–48. (In Japanese) [Google Scholar]
- Charrier, B.; Abreu, M.H.; Araujo, R.; Bruhn, A.; Coates, J.C.; De Clerck, O.; Katsaros, C.; Robaina, R.R.; Wichard, T. Furthering knowledge of seaweed growth and development to facilitate sustainable aquaculture. New Phytol. 2017, 216, 967–975. [Google Scholar] [CrossRef] [Green Version]
- Casas, G.N.; Piriz, M.L. Surveys of Undaria pinnatifida (Laminariales Phaeophyta) in Golfo Nuevo, Argentina. Hydrobiologia 1996, 326/327, 213–215. [Google Scholar] [CrossRef]
- Dellatorre, F.G.; Amoroso, R.; Saravia, J.; Orensanz, J.M. Rapid expansion and potential range of the invasive kelp Undaria pinnatifida in the Southwest Atlantic. Aquat. Invasions 2014, 9, 467–478. [Google Scholar] [CrossRef]
- Castric-Fey, A.; Beaupoil, C.; Bouchain, J.; Pradier, E.; L’Hardy-Halos, M.T. The introduced alga Undaria pinnatifida (Laminariales, Alariaceae) in the rocky shore ecosystem of the St. Malo area: Growth rates and longevity of the sporophyte. Bot. Mar. 1999, 42, 83–96. [Google Scholar] [CrossRef]
- Silva, P.C.; Woodfield, R.A.; Cohen, A.N.; Harris, L.H.; Goddard, J.H.R. First report of the Asian kelp Undaria pinnatifida in the northeastern Pacific Ocean. Biol. Invasions 2002, 4, 333–338. [Google Scholar] [CrossRef]
- Cecere, E.; Petroccelli, A.; Saracino, O.D. Undaria pinnatifida (Fucophyceae, Laminariales) spread in the central Mediterranean: Its occurrence in the Mar Piccolo of Taranto (Ionian Sea, southern Italy). Cryptogam. Algol. 2000, 21, 305–309. [Google Scholar] [CrossRef]
- Sanderson, J.C. A preliminary survey of the distribution of the introduced macroalga, Undaria pinnatifida (Harvey) Suringar on the east coast of Tasmania. Bot. Mar. 1990, 33, 153–157. [Google Scholar] [CrossRef]
- Hay, C.H.; Luckens, P.A. The Asian kelp Undaria pinnatifida (Phaeophyta: Laminariales) found in a New Zealand harbour. N. Z. J. Bot. 1987, 25, 364–366. [Google Scholar] [CrossRef] [Green Version]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A selection from the Global Invasive Species Database; The Invasive Species Specialist Group (ISSG) a specialist group of the Species Survival Commission (SSC) of the World Conservation Union (IUCN), Hollands Printing Ltd.: Auckland, New Zealand, 2000; p. 12, (reprinted November 2004). [Google Scholar]
- Roleda, M.Y. Stress physiology and reproductive phenology of Arctic endemic kelp Laminaria solidungula J. Agardh. Pol. Biol. 2016, 39, 1967–1977. [Google Scholar] [CrossRef]
- Assis, J.; Berecibar, E.; Claro, B.; Alberto, F.; Reed, D.; Raimondi, P.T.; Serrao, E.A. Major shifts at the range edge of marine forests: The combined effects of climate changes and limited dispersal. Sci. Rep. 2017, 7, 44348. [Google Scholar] [CrossRef] [Green Version]
- Leal, P.P.; Hurd, C.L.; Sander, S.G.; Armstrong, E.; Fernández, P.A.; Suhrhoff, T.J.; Roleda, M.Y. Copper pollution exacerbates the effects of ocean acidification and warming on kelp microscopic early life stages. Sci. Rep. 2018, 8, 14763. [Google Scholar] [CrossRef]
- Matsui, T.; Ohgai, M.; Ohshima, Y.; Kohara, K. The effects of light quality and quantity on gametophyte growth and fertility, and young sporophyte growth, in several species of Laminariales. Nippon Suisan Gakkaishi 1992, 58, 1257–1265. [Google Scholar] [CrossRef] [Green Version]
- Pang, S.J.; Wu, C.Y. Study on gametophyte vegetative growth of Undaria pinnatifida and its applications. Chin. J. Oceanol. Limnol. 1996, 14, 205–211. [Google Scholar]
- Morita, T.; Kurashima, A.; Maegawa, M. Temperature requirements for the growth and maturation of the gametophytes of Undaria pinnatifida and U. undarioides (Laminariales, Phaeophyceae). Phycol. Res. 2003, 51, 154–160. [Google Scholar] [CrossRef]
- Xu, Z.; Dapeng, L.; Hanhua, H.; Tianwei, T. Growth promotion of vegetative gametophytes of Undaria pinnatifida by blue light. Biotech. Let. 2005, 27, 1467–1475. [Google Scholar] [CrossRef] [PubMed]
- Baba, M. Effects of temperature, irradiance and salinity on the growth of Undaria pinnatifida from Niigata Prefecture, Central Japan. Rep. Mar. Ecol. Res. Inst. 2008, 11, 7–15. (In Japanese) [Google Scholar]
- Pang, S.J.; Shan, T.F.; Zhang, Z.H. Responses of vegetative gametophytes of Undaria pinnatifida to high irradiance in the process of gametogenesis. Phycol. Res. 2008, 56, 280–287. [Google Scholar] [CrossRef]
- Watanabe, Y.; Nishihara, G.N.; Tokunaga, S.; Terada, R. The effect of irradiance and temperature responses and the phenology of a native alga, Undaria pinnatifida (Laminariales), at the southern limit of its natural distribution in Japan. J. Appl. Phycol. 2014, 26, 2405–2415. [Google Scholar] [CrossRef]
- Gao, X.; Endo, H.; Taniguchi, K.; Agatsuma, Y. Genetic differentiation of high-temperature tolerance in the kelp Undaria pinnatifida sporophytes from geographically separated populations along the Pacific coast of Japan. J. Appl. Phycol. 2013, 25, 567–574. [Google Scholar] [CrossRef]
- Sato, Y.; Hirano, T.; Niwa, K.; Suzuki, T.; Fukunishi, N.; Abe, T.; Kawano, S. Phenotypic differentiation in the morphology and nutrient uptake kinetics among Undaria pinnatifida thalli cultivated at six sites in Japan. J. Appl. Phycol. 2016, 28, 3447–3458. [Google Scholar] [CrossRef]
- Sato, Y.; Hirano, T.; Ichida, H.; Murakami, M.; Fukunishi, N.; Abe, T.; Kawano, S. Morphological and physiological characterization of Undaria pinnatifida originating from five cultivation sites in Japan through common garden experiments using a tank culture system. J. Appl. Phycol. 2017, 29, 2287–2295. [Google Scholar] [CrossRef]
- Baş, D.; Boyacı, İ.H. Modelling and optimization II: Comparison of estimation capabilities of response surface methodology with artificial neural networks in a biochemical reaction. J. Food Eng. 2007, 78, 846–854. [Google Scholar] [CrossRef]
- Myers, R.H.; Montgomery, D.C.; Anderson-Cook, C.M. Response Surface Methodology: Process and Product Optimization Using Designed Experiments, 3rd ed.; Wiley: Hoboken, NJ, USA, 2009; p. 856. [Google Scholar]
- Amini, M.; Younesi, H.; Bahramifar, N.; Lorestani, A.A.Z.; Ghorbani, F.; Daneshi, A.; Sharifzadeh, M. Application of response surface methodology for optimization of lead biosorption in an aqueous solution by Aspergillus niger. J. Hazard. Mater. 2008, 154, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Annadurai, G.; Ling, L.Y.; Lee, J.F. Statistical optimization of medium components and growth conditions by response surface methodology to enhance phenol degradation by Pseudomonas putida. J. Hazard. Mater. 2008, 151, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Kalil, S.J.; Maugeri, F.; Rodrigues, M.I. Response surface analysis and simulation as a tool for bioprocess design and optimization. Proc. Biochem. 2000, 35, 539–550. [Google Scholar] [CrossRef]
- Aktaș, N.; Boyacı, İ.H.; Mutlu, M.; Tanyolaç, A. Optimization of lactose utilization in deproteinated whey by Kluyveromyces marxianus using response surface methodology (RSM). Biores. Tech. 2006, 97, 2252–2259. [Google Scholar]
- Denavi, G.; Tapia-Blácido, D.R.; Añón, M.C.; Sobral, P.J.A.; Mauri, A.N.; Menegalli, F.C. Effects of drying conditions on some physical properties of soy protein films. J. Food Eng. 2009, 90, 341–349. [Google Scholar] [CrossRef]
- Hallenbeck, P.; Grogger, M.; Mraz, M.; Veverka, D. The use of Design of Experiments and Response Surface Methodology to optimize biomass and lipid production by the oleaginous marine green alga, Nannochloropsis gaditana in response to light intensity, inoculum size and CO2. Biores. Tech. 2015, 184, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Nazir, Y.; Shuib, S.; Kalil, M.S.; Song, Y.; Hamid, A.A. Optimization of culture conditions for enhanced growth, lipid and docosahexaenoic acid (DHA) production of Aurantiochytrium SW1 by Response Surface Methodology. Sci. Rep. 2018, 8, 8909. [Google Scholar] [CrossRef]
- Takayama, K.; Morva, A.; Fujikawa, M.; Hattori, Y.; Obata, Y.; Nagai, T. Formula optimization of theophylline controlled-release tablet based on artificial neural networks. J Control. Release 2000, 68, 175–186. [Google Scholar] [CrossRef]
- Derringer, G.; Suich, R. Simultaneous optimization of several response variables. J. Qual. Technol. 1980, 12, 214–219. [Google Scholar] [CrossRef]
- Tatewaki, M. Formation of a crustose sporophyte with unilocular sporangia in Scytosiphon lomentaria. Phycologia 1966, 6, 62–66. [Google Scholar] [CrossRef]
- Antony, J. Design of Experiments for Engineers and Scientists, 2nd ed.; Elsevier: Jamestown Road, UK, 2014; pp. 154–196. [Google Scholar]
- Uwai, S.; Yotsukura, N.; Serisawa, Y.; Muraoka, D.; Hiraoka, M.; Kogame, K. Intraspecific genetic diversity of Undaria pinnatifida in Japan, based on the mitochondrial cox3 gene and the ITS1 of nrDNA. Hydrobiologia 2006, 553, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.A.; Brinkhuis, B.H. Seasonal light and temperature interaction effects on development of Laminaria saccharina (Phaeophyta) gametophytes and juvenile sporophytes. J. Phycol. 1988, 24, 181–191. [Google Scholar] [CrossRef]
- Monteiro, C.; Heinrich, S.; Bartsch, I.; Valentin, K.; Corre, E.; Collén, J.; Harms, L.; Glöckner, G.; Bischof, K. Temperature modulates sex-biased gene expression in the gametophytes of the kelp Saccharina latissima. Front. Mar. Sci. 2019, 6, 769. [Google Scholar] [CrossRef] [Green Version]
- Dieck, I.T. Temperature tolerance and survival in darkness of kelp gametophytes (Laminariales, Phaeophyta): Ecological and biogeographical implications. Mar. Ecol. Prog. Ser. 1993, 100, 253–264. [Google Scholar] [CrossRef]
- Norton, T.A. Experiments on the factors influencing the geographical distributions of Saccorhiza polyschides and Saccorhiza dermatodea. New Phytol. 1977, 78, 625–635. [Google Scholar] [CrossRef]
- Lee, J.A.; Brinkhuis, B.H. Reproductive phenology of Laminaria saccharina (L.) Lamour. (Phaeophyta) at the southern limit of its distribution in the northwestern Atlantic Ocean. J. Phycol. 1986, 22, 276–285. [Google Scholar] [CrossRef]
- Borlongan, I.A.; Matsumoto, K.; Nakazaki, Y.; Shimada, N.; Kozono, J.; Nishihara, G.N.; Shimada, S.; Watanabe, Y.; Terada, R. Photosynthetic activity of two life history stages of Costaria costata (Laminariales, Phaeophyceae) in response to PAR and temperature gradient. Phycologia 2018, 57, 159–168. [Google Scholar] [CrossRef]
- Borlongan, I.A.; Nishihara, G.N.; Shimada, S.; Terada, R. Assessment of photosynthetic performance in the two life history stages of Alaria crassifolia (Laminariales, Phaeophyceae). Phycol. Res. 2019, 67, 28–38. [Google Scholar] [CrossRef]
- Lüning, K.; Neushul, M. Light and temperature demands for growth and reproduction of laminarian gametophytes in sourthern and central California. Mar. Biol. 1978, 45, 297–309. [Google Scholar] [CrossRef]
- Saito, Y. An ecological study of Undaria pinnatifida Sur. I. On the influence of environmental factors upon the development of gametophytes. Bull. Jap. Soc. Sci. Fish 1956, 22, 229–234. (In Japanese) [Google Scholar] [CrossRef] [Green Version]
- Lüning, K.; Dring, M.J. Reproduction induced by blue light in female gametophytes of Laminaria saccharina. Planta 1972, 104, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Lüning, K.; Dring, M.J. Reproduction, growth and photosynthesis of gametophytes of Laminaria saccharina grown in blue and red light. Mar. Biol. 1975, 29, 195–200. [Google Scholar] [CrossRef]
- Wang, W.J.; Wang, F.J.; Sun, X.T.; Liu, F.L.; Liang, Z.R. Comparison of transcriptome under red and blue light culture of Saccharina japonica (Phaeophyceae). Planta 2013, 237, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Yao, J.; Fu, G.; Guo, H.; Duan, D. Isolation, expression, and characterization of blue Light receptor AUREOCHROME gene from Saccharina japonica (Laminariales, Phaeophyceae). Mar. Biotech. 2014, 16, 135–143. [Google Scholar] [CrossRef]
- Pearson, G.A.; Serrão, E.A.; Dring, M.; Schmid, R. Blue- and green-light signals for gamete release in the brown alga, Silvetia compressa. Oecologia 2004, 138, 193–201. [Google Scholar] [CrossRef]
- Kawashima, Y.; Tokuda, H. Regeneration from callus of Undaria pinnatifida (Harvey) Suringar (Laminariales, Phaeophyta). Hydrobiologia 1993, 260, 385–389. [Google Scholar] [CrossRef]
- Lewis, R.J.; Jiang, B.Y.; Neushul, M.; Fei, X.G. Haploid parthenogenic sporophytes of Laminaria japonica (Phaeophyceae). J. Phycol. 1993, 29, 363–369. [Google Scholar] [CrossRef]
- Oppliger, L.V.; Correa, J.A.; Peters, A.F. Parthenogenesis in the brown alga Lessonia nigrescens (Laminariales, Phaeophyceae) from central Chile. J. Phycol. 2007, 43, 1295–1301. [Google Scholar] [CrossRef]
- Lotze, H.K.; Worm, B. Complex interactions of climatic and ecological controls on macroalgal recruitment. Limnol. Oceanogr. 2002, 46, 1734–1741. [Google Scholar] [CrossRef]
- Hurd, C.L.; Harrison, P.J.; Bischof, K.; Lobban, C.S. Seaweed Ecology and Physiology, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014; p. 551. [Google Scholar]
- Tanada, N.; Dan, A.; Kusaka, K.; Oka, N.; Hamano, T. A new practical method for mass seedling production of Undaria pinnatifida (Harvey) Suringar using free-living gametophytes that originated from single zoospores and its application for commercial cultivation. Algal Resour. 2015, 8, 23–36. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Factor 1 (X1) | Factor 2 (X2) | Factor 3 (X3) | Response (Y, RGR) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | Light Intensity (µmol m−2 s−2) | Light Color | Male | Female | ||||||
| IWT | TKS | KGS | IWT | TKS | KGS | |||||
| 1 | 15 | 2 | Green | 0.504 | 0.780 | 1.073 | 0.538 | 0.714 | 1.022 | |
| 2 | 15 | 21 | Red | 0.473 | 0.627 | 1.114 | 0.649 | 1.006 | 1.090 | |
| 3 | 15 | 21 | Blue | 0.960 | 1.250 | 1.905 | 1.018 | 1.522 | 1.587 | |
| 4 | 15 | 40 | Green | 1.09 | 1.459 | 1.724 | 1.042 | 1.573 | 1.515 | |
| 5 | 16 | 32 | White | 1.109 | 1.293 | 1.887 | 1.066 | 1.494 | 1.524 | |
| 6 | 20 | 2 | Blue | 0.771 | 1.017 | 1.343 | 0.689 | 0.783 | 1.043 | |
| 7 | 20 | 2 | White | 0.739 | 0.841 | 1.218 | 0.617 | 0.627 | 0.932 | |
| 8 | 20 | 21 | Green | 1.231 | 1.743 | 1.971 | 1.065 | 1.527 | 1.772 | |
| 9 | 20 | 40 | Blue | 1.277 | 1.672 | 1.994 | 1.134 | 1.517 | 1.536 | |
| 10 | 25 | 2 | Red | 0.224 | 0.246 | 0.454 | 0.128 | 0.193 | 0.413 | |
| 11 | 25 | 2 | Green | 0.733 | 1.004 | 1.143 | 0.460 | 0.709 | 0.992 | |
| 12 | 25 | 21 | Blue | 1.110 | 1.450 | 1.975 | 0.881 | 1.393 | 1.556 | |
| 13 | 25 | 40 | Red | 0.652 | 0.878 | 1.105 | 0.515 | 0.804 | 0.907 | |
| 14 | 25 | 40 | White | 1.107 | 1.325 | 1.770 | 0.812 | 1.165 | 1.292 | |
| 15 | 25 | 40 | Green | 1.161 | 1.635 | 1.794 | 0.847 | 1.32 | 1.485 | |
| Sex | Assay | Factor 1 (X1) | Factor 2 (X2) | Factor 3 (X3) | Maturation Degree at the Final Date of Cultivation | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | Light Intensity(µmol m−2 s−2) | Light Color | IWT | TKS | KGS | ||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | |||||
| 1 | 10 | 10 | Blue | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.03 | 0.20 | 0.77 | 0.91 | 0.04 | 0.03 | 0.01 | 0.00 | |
| 2 | 10 | 30 | White | 0.00 | 0.00 | 0.00 | 0.02 | 0.98 | 0.00 | 0.00 | 0.04 | 0.22 | 0.74 | 0.83 | 0.08 | 0.06 | 0.03 | 0.01 | |
| 3 | 10 | 50 | Blue | 0.00 | 0.00 | 0.01 | 0.33 | 0.66 | 0.00 | 0.00 | 0.02 | 0.15 | 0.82 | 0.54 | 0.16 | 0.17 | 0.10 | 0.02 | |
| 4 | 16 | 26 | White | 0.00 | 0.00 | 0.00 | 0.02 | 0.98 | 0.00 | 0.00 | 0.00 | 0.02 | 0.98 | 0.00 | 0.00 | 0.00 | 0.02 | 0.98 | |
| 5 | 16 | 26 | White | 0.00 | 0.00 | 0.03 | 0.51 | 0.46 | 0.00 | 0.00 | 0.00 | 0.02 | 0.98 | 0.00 | 0.00 | 0.00 | 0.02 | 0.98 | |
| Male | 6 | 17 | 50 | White | 0.00 | 0.00 | 0.00 | 0.02 | 0.98 | 0.00 | 0.00 | 0.00 | 0.01 | 0.98 | 0.04 | 0.03 | 0.11 | 0.38 | 0.43 |
| 7 | 18 | 10 | Blue | 0.00 | 0.00 | 0.00 | 0.09 | 0.90 | 0.00 | 0.00 | 0.00 | 0.01 | 0.99 | 0.11 | 0.09 | 0.23 | 0.38 | 0.19 | |
| 8 | 19 | 34 | Blue | 0.00 | 0.00 | 0.02 | 0.33 | 0.65 | 0.00 | 0.00 | 0.00 | 0.01 | 0.99 | 0.11 | 0.09 | 0.22 | 0.39 | 0.20 | |
| 9 | 19 | 34 | Blue | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.01 | 0.99 | 0.06 | 0.05 | 0.16 | 0.41 | 0.32 | |
| 10 | 25 | 10 | White | 0.00 | 0.00 | 0.00 | 0.06 | 0.94 | 0.00 | 0.00 | 0.00 | 0.02 | 0.98 | 0.08 | 0.07 | 0.20 | 0.41 | 0.24 | |
| 11 | 25 | 30 | Blue | 0.00 | 0.00 | 0.00 | 0.02 | 0.98 | 0.00 | 0.00 | 0.00 | 0.01 | 0.99 | 0.00 | 0.00 | 0.00 | 0.01 | 0.99 | |
| 12 | 25 | 50 | White | 0.00 | 0.00 | 0.01 | 0.22 | 0.77 | 0.00 | 0.00 | 0.00 | 0.01 | 0.98 | 0.10 | 0.08 | 0.21 | 0.40 | 0.22 | |
| 1 | 10 | 10 | Blue | 0.84 | 0.12 | 0.02 | 0.01 | 0.01 | 0.94 | 0.05 | 0.01 | 0.00 | 0.01 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 2 | 10 | 30 | White | 0.80 | 0.15 | 0.02 | 0.01 | 0.01 | 0.86 | 0.10 | 0.02 | 0.01 | 0.01 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 3 | 10 | 50 | Blue | 0.30 | 0.37 | 0.13 | 0.09 | 0.12 | 0.82 | 0.13 | 0.02 | 0.01 | 0.02 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 4 | 16 | 26 | White | 0.13 | 0.28 | 0.16 | 0.15 | 0.28 | 0.26 | 0.34 | 0.14 | 0.09 | 0.18 | 0.56 | 0.37 | 0.04 | 0.02 | 0.01 | |
| 5 | 16 | 26 | White | 0.13 | 0.28 | 0.16 | 0.15 | 0.28 | 0.26 | 0.34 | 0.14 | 0.09 | 0.18 | 0.56 | 0.37 | 0.04 | 0.02 | 0.01 | |
| 6 | 17 | 50 | White | 0.04 | 0.12 | 0.11 | 0.15 | 0.58 | 0.12 | 0.24 | 0.15 | 0.13 | 0.36 | 0.50 | 0.41 | 0.05 | 0.02 | 0.01 | |
| Female | 7 | 18 | 10 | Blue | 0.10 | 0.25 | 0.16 | 0.16 | 0.34 | 0.28 | 0.34 | 0.13 | 0.08 | 0.16 | 0.95 | 0.05 | 0.00 | 0.00 | 0.00 |
| 8 | 19 | 34 | Blue | 0.02 | 0.06 | 0.06 | 0.10 | 0.76 | 0.03 | 0.07 | 0.07 | 0.09 | 0.74 | 0.19 | 0.52 | 0.16 | 0.08 | 0.05 | |
| 9 | 19 | 34 | Blue | 0.02 | 0.06 | 0.06 | 0.10 | 0.76 | 0.03 | 0.07 | 0.07 | 0.09 | 0.74 | 0.19 | 0.52 | 0.16 | 0.08 | 0.05 | |
| 10 | 25 | 10 | White | 0.91 | 0.07 | 0.01 | 0.01 | 0.01 | 0.78 | 0.16 | 0.03 | 0.01 | 0.02 | 0.89 | 0.10 | 0.01 | 0.00 | 0.00 | |
| 11 | 25 | 30 | Blue | 0.30 | 0.37 | 0.13 | 0.09 | 0.12 | 0.07 | 0.17 | 0.13 | 0.13 | 0.50 | 0.81 | 0.17 | 0.01 | 0.01 | 0.00 | |
| 12 | 25 | 50 | White | 0.44 | 0.35 | 0.09 | 0.05 | 0.07 | 0.18 | 0.30 | 0.15 | 0.11 | 0.25 | 0.87 | 0.12 | 0.01 | 0.00 | 0.00 | |
| Line | Sex | Source | Sum of Square | df | Mean Square | F | Significance |
|---|---|---|---|---|---|---|---|
| IWT | Male | Regression | 31.576 | 8 | 3.947 | 179.95 | <0.0001 |
| Residual | 6.953 | 317 | 0.022 | ||||
| Total | 38.529 | 325 | |||||
| Female | Regression | 25.278 | 8 | 3.160 | 174.42 | <0.0001 | |
| Residual | 5.743 | 317 | 0.018 | ||||
| Total | 31.021 | 325 | |||||
| TKS | Male | Regression | 54.291 | 8 | 6.786 | 61.52 | <0.0001 |
| Residual | 34.305 | 311 | 0.110 | ||||
| Total | 88.596 | 319 | |||||
| Female | Regression | 57.178 | 8 | 7.147 | 153.79 | <0.0001 | |
| Residual | 14.500 | 312 | 0.046 | ||||
| Total | 71.678 | 320 | |||||
| KGS | Male | Regression | 56.814 | 7 | 8.116 | 149.60 | <0.0001 |
| Residual | 14.431 | 266 | 0.054 | ||||
| Total | 71.245 | 273 | |||||
| Female | Regression | 44.490 | 7 | 6.356 | 163.47 | <0.0001 | |
| Residual | 13.530 | 348 | 0.039 | ||||
| Total | 58.019 | 355 |
| Environmental Factor | IWT | TKS | KGS | |||
|---|---|---|---|---|---|---|
| Male | Female | Male | Female | Male | Female | |
| Temperature (°C) | 20.7 | 18.6 | 20.9 | 16.5 | 20.7 | 19.8 |
| Light intensity (µmol m−2 s−1) | 33.7 | 32.5 | 32.7 | 31.3 | 28.6 | 26.9 |
| Light color | Blue | Blue | Green | Blue | Blue | Blue |
| Line | Sex | Log Likelihood × (−1) | df | 𝝌2 | Significance |
|---|---|---|---|---|---|
| IWT | Male | 1475.97 | 7 | 3827.07 | <0.001 |
| Female | 1496.28 | 7 | 1461.8 | <0.001 | |
| TKS | Male | 1549.45 | 7 | 3625.8 | <0.001 |
| Female | 1148.54 | 7 | 1240.27 | <0.001 | |
| KGS | Male | 226.21 | 7 | 133.92 | <0.001 |
| Female | 461.88 | 7 | 552.48 | <0.001 |
| Environmental Factor | IWT | TKS | KGS | |||
|---|---|---|---|---|---|---|
| Male | Female | Male | Female | Male | Female | |
| Temperature (°C) | 19.5 | 17.9 | 19.5 | 18.7 | 20.6 | 20.6 |
| Light intensity (µmol m−2 s−1) | 39.8 | 50 | 39.3 | 44.6 | 50 | 32.5 |
| Light color | Blue | Blue | Blue | Blue | White | White |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, Y.; Endo, H.; Oikawa, H.; Kanematsu, K.; Naka, H.; Mogamiya, M.; Kawano, S.; Kazama, Y. Sexual Difference in the Optimum Environmental Conditions for Growth and Maturation of the Brown Alga Undaria pinnatifida in the Gametophyte Stage. Genes 2020, 11, 944. https://doi.org/10.3390/genes11080944
Sato Y, Endo H, Oikawa H, Kanematsu K, Naka H, Mogamiya M, Kawano S, Kazama Y. Sexual Difference in the Optimum Environmental Conditions for Growth and Maturation of the Brown Alga Undaria pinnatifida in the Gametophyte Stage. Genes. 2020; 11(8):944. https://doi.org/10.3390/genes11080944
Chicago/Turabian StyleSato, Yoichi, Hikaru Endo, Hiroki Oikawa, Koichi Kanematsu, Hiroyuki Naka, Miho Mogamiya, Shigeyuki Kawano, and Yusuke Kazama. 2020. "Sexual Difference in the Optimum Environmental Conditions for Growth and Maturation of the Brown Alga Undaria pinnatifida in the Gametophyte Stage" Genes 11, no. 8: 944. https://doi.org/10.3390/genes11080944
APA StyleSato, Y., Endo, H., Oikawa, H., Kanematsu, K., Naka, H., Mogamiya, M., Kawano, S., & Kazama, Y. (2020). Sexual Difference in the Optimum Environmental Conditions for Growth and Maturation of the Brown Alga Undaria pinnatifida in the Gametophyte Stage. Genes, 11(8), 944. https://doi.org/10.3390/genes11080944