Rimbp, a New Marker for the Nervous System of the Tunicate Ciona robusta

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
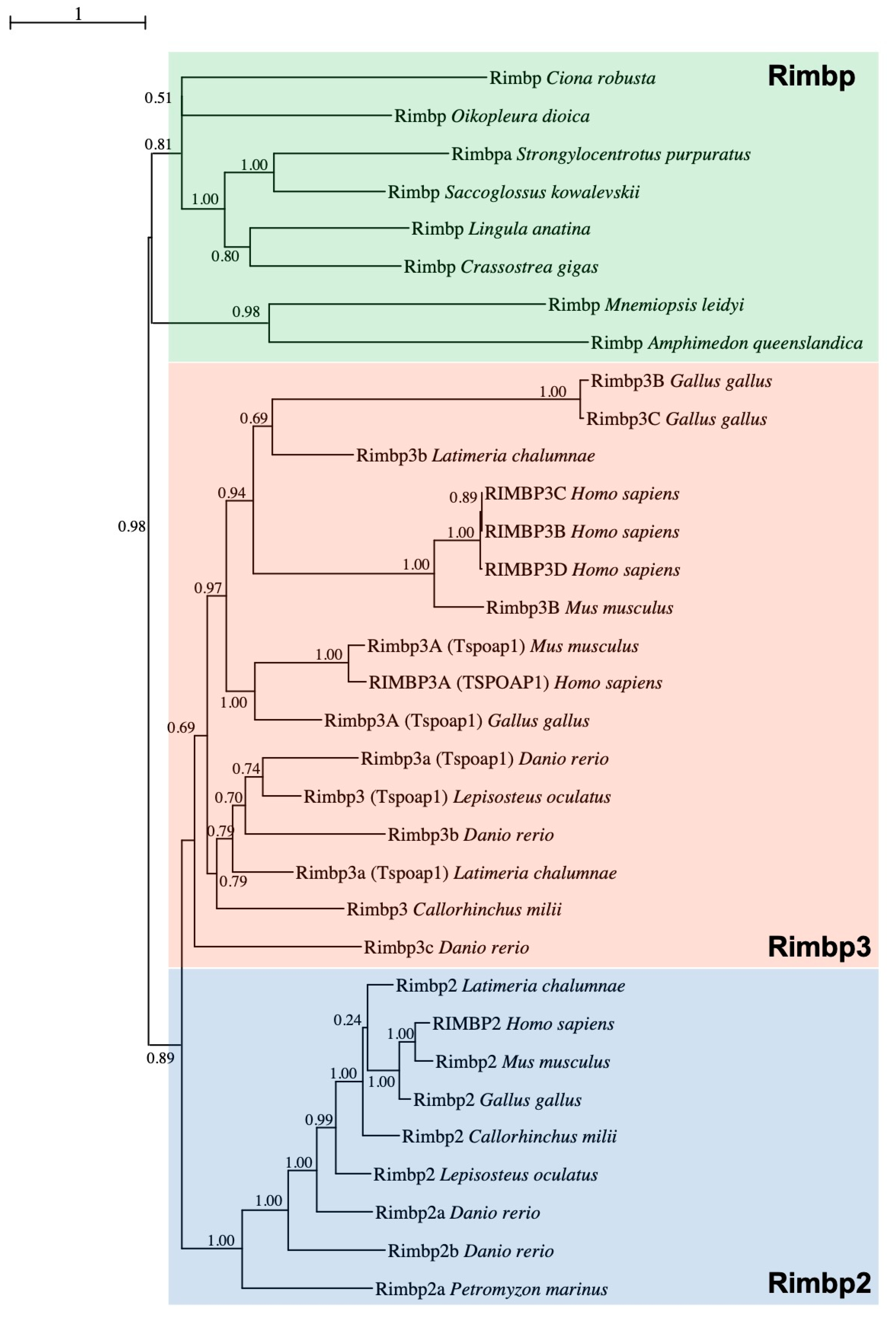
2.1. Phylogenetic and Synteny Analysis
2.2. Animals and Embryo Electroporation
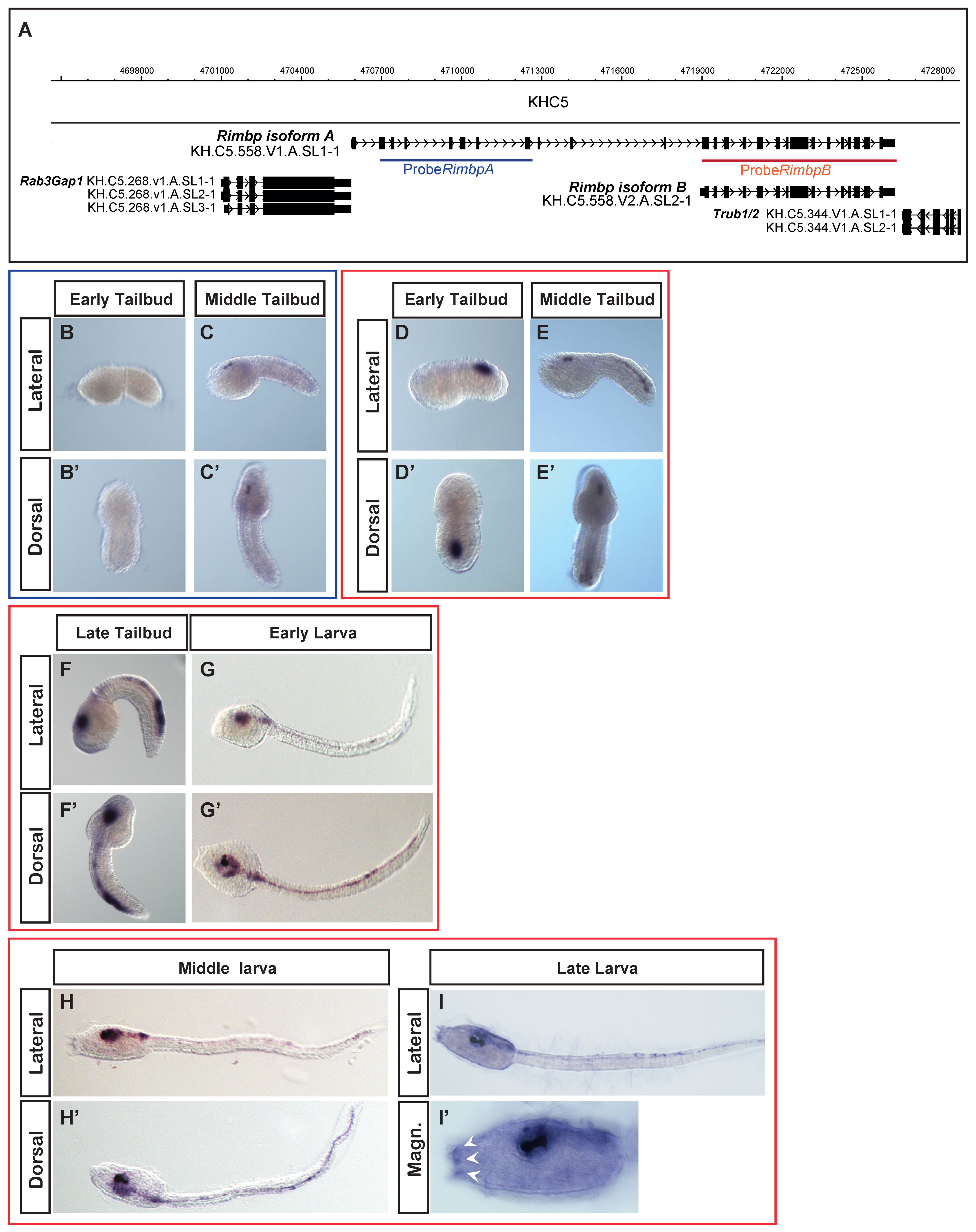
2.3. In Situ Hybridization
2.4. Molecular Cloning
3. Results
3.1. Evolutionary Survey of Rimbp2/3 Genes
3.2. Dynamic Expression Pattern of Rimbp in the Ciona robusta Nervous System
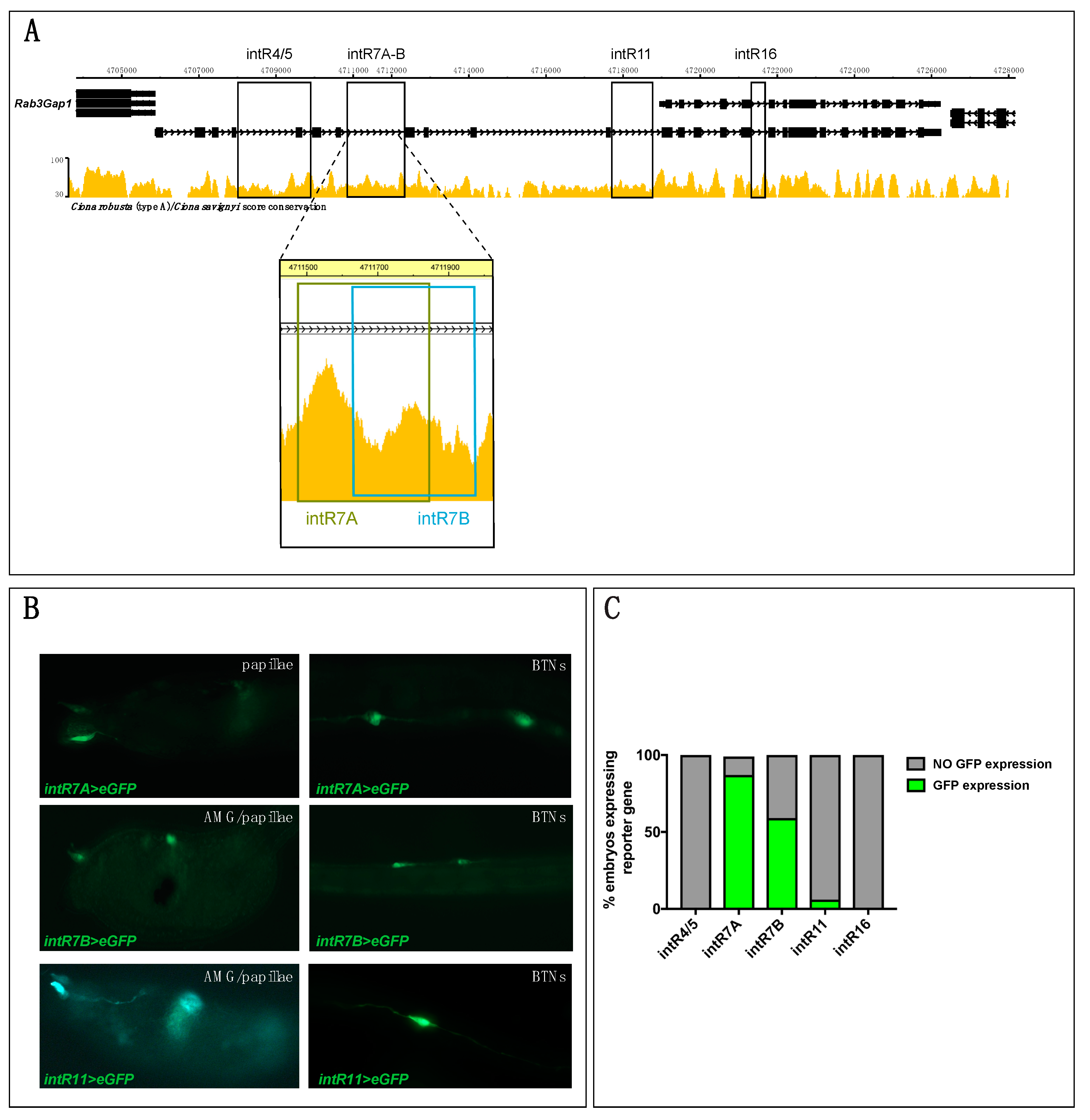
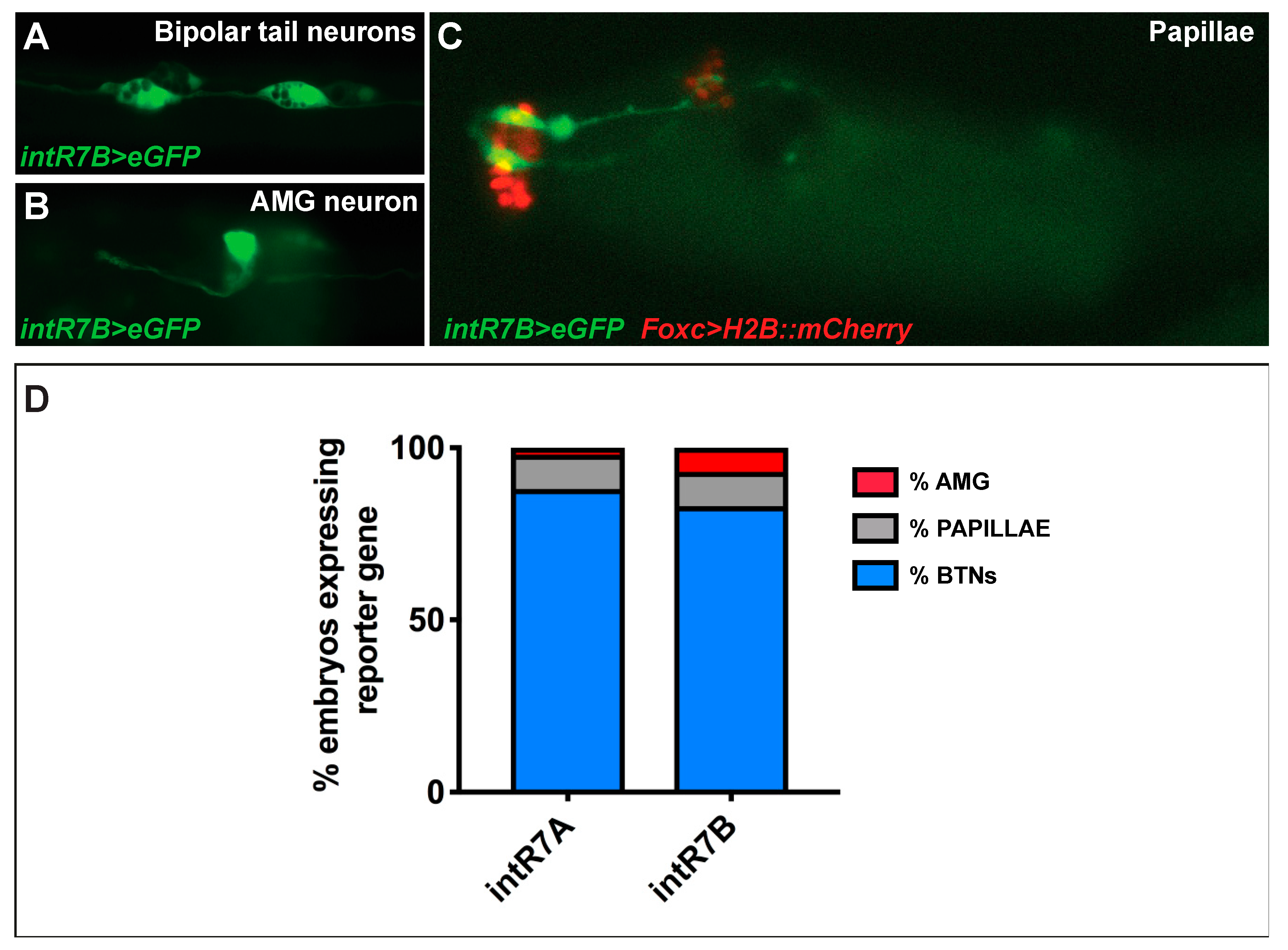
3.3. Intronic Cis-Regulatory Elements for Rimbp Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Davis, G.W. Homeostatic control of neural activity: From phenomenology to molecular design. Annu. Rev. Neurosci. 2006, 29, 307–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marder, E. Variability, compensation, and modulation in neurons and circuits. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 3), 15542–15548. [Google Scholar] [CrossRef] [Green Version]
- Turrigiano, G.G. The self-tuning neuron: Synaptic scaling of excitatory synapses. Cell 2008, 135, 422–435. [Google Scholar] [CrossRef] [Green Version]
- Davis, G.W. Homeostatic signaling and the stabilization of neural function. Neuron 2013, 80, 718–728. [Google Scholar] [CrossRef] [Green Version]
- Plomp, J.J.; van Kempen, G.T.; Molenaar, P.C. Adaptation of quantal content to decreased postsynaptic sensitivity at single endplates in alpha-bungarotoxin-treated rats. J. Physiol. 1992, 458, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Wojcik, S.M.; Brose, N. Regulation of membrane fusion in synaptic excitation-secretion coupling: Speed and accuracy matter. Neuron 2007, 55, 11–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittelstaedt, T.; Alvaréz-Baron, E.; Schoch, S. RIM proteins and their role in synapse function. Biol. Chem. 2010, 391, 599–606. [Google Scholar] [CrossRef]
- Kaeser, P.S.; Deng, L.; Wang, Y.; Dulubova, I.; Liu, X.; Rizo, J.; Südhof, T.C. RIM Proteins Tether Ca2 Channels to Presynaptic Active Zones via a Direct PDZ-Domain Interaction. Cell 2011, 144, 282–295. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Sugita, S.; Sudhof, T.C. The RIM/NIM family of neuronal C2 domain proteins. Interactions with Rab3 and a new class of Src homology 3 domain proteins. J. Biol. Chem. 2000, 275, 20033–20044. [Google Scholar] [CrossRef] [Green Version]
- Hibino, H.; Pironkova, R.; Onwumere, O.; Vologodskaia, M.; Hudspeth, A.J.; Lesage, F. RIM Binding Proteins (RBPs) Couple Rab3-Interacting Molecules (RIMs) to Voltage-Gated Ca2 Channels. Neuron 2002, 34, 411–423. [Google Scholar] [CrossRef] [Green Version]
- Schoch, S.; Gundelfinger, E.D. Molecular organization of the presynaptic active zone. Cell Tissue Res. 2006, 326, 379–391. [Google Scholar] [CrossRef]
- Schoch, S.; Mittelstaedt, T.; Kaeser, P.S.; Padgett, D.; Feldmann, N.; Chevaleyre, V.; Castillo, P.E.; Hammer, R.E.; Han, W.; Schmitz, F.; et al. Redundant functions of RIM1α and RIM2α in Ca2 -triggered neurotransmitter release. EMBO J. 2006, 25, 5852–5863. [Google Scholar] [CrossRef]
- Graf, E.R.; Daniels, R.W.; Burgess, R.W.; Schwarz, T.L.; DiAntonio, A. Rab3 dynamically controls protein composition at active zones. Neuron 2009, 64, 663–677. [Google Scholar] [CrossRef] [Green Version]
- Davydova, D.; Marini, C.; King, C.; Klueva, J.; Bischof, F.; Romorini, S.; Montenegro-Venegas, C.; Heine, M.; Schneider, R.; Schröder, M.S.; et al. Bassoon Specifically Controls Presynaptic P/Q-type Ca2 Channels via RIM-Binding Protein. Neuron 2014, 82, 181–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittelstaedt, T.; Schoch, S. Structure and evolution of RIM-BP genes: Identification of a novel family member. Gene 2007, 403, 70–79. [Google Scholar] [CrossRef]
- Girotto, G.; Vuckovic, D.; Buniello, A.; Lorente-Cánovas, B.; Lewis, M.; Gasparini, P.; Steel, K.P. Expression and replication studies to identify new candidate genes involved in normal hearing function. PLoS ONE 2014, 9, e85352. [Google Scholar] [CrossRef]
- Okuda, H.; DeBoer, K.; O’Connor, A.E.; Merriner, D.J.; Jamsai, D.; O’Bryan, M.K. LRGUK1 is part of a multiprotein complex required for manchette function and male fertility. FASEB J. 2017, 31, 1141–1152. [Google Scholar] [CrossRef] [Green Version]
- Kittel, R.J. Bruchpilot Promotes Active Zone Assembly, Ca2 Channel Clustering, and Vesicle Release. Science 2006, 312, 1051–1054. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.S.Y.; Siebert, M.; Mertel, S.; Knoche, E.; Wegener, S.; Wichmann, C.; Matkovic, T.; Muhammad, K.; Depner, H.; Mettke, C.; et al. RIM-Binding Protein, a Central Part of the Active Zone, Is Essential for Neurotransmitter Release. Science 2011, 334, 1565–1569. [Google Scholar] [CrossRef] [Green Version]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef]
- Gertz, E.M.; Yu, Y.-K.; Agarwala, R.; Schäffer, A.A.; Altschul, S.F. Composition-based statistics and translated nucleotide searches: Improving the TBLASTN module of BLAST. BMC Biol. 2006, 4, 41. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, D.H.; Scornavacca, C. Dendroscope 3: An interactive tool for rooted phylogenetic trees and networks. Syst. Biol. 2012, 61, 1061–1067. [Google Scholar] [CrossRef] [Green Version]
- de Castro, E.; Sigrist, C.J.A.; Gattiker, A.; Bulliard, V.; Langendijk-Genevaux, P.S.; Gasteiger, E.; Bairoch, A.; Hulo, N. ScanProsite: Detection of PROSITE signature matches and ProRule-associated functional and structural residues in proteins. Nucleic Acids Res. 2006, 34, W362–W365. [Google Scholar] [CrossRef]
- Christiaen, L.; Wagner, E.; Shi, W.; Levine, M. Isolation of Sea Squirt (Ciona) Gametes, Fertilization, Dechorionation, and Development. Cold Spring Harb. Protoc. 2009, 2009, pdb.prot5344. [Google Scholar] [CrossRef]
- Christiaen, L.; Wagner, E.; Shi, W.; Levine, M. Whole-Mount In Situ Hybridization on Sea Squirt (Ciona intestinalis) Embryos. Cold Spring Harb. Protoc. 2009, 2009, pdb.prot5348. [Google Scholar] [CrossRef]
- Racioppi, C.; Kamal, A.K.; Razy-Krajka, F.; Gambardella, G.; Zanetti, L.; di Bernardo, D.; Sanges, R.; Christiaen, L.A.; Ristoratore, F. Fibroblast growth factor signalling controls nervous system patterning and pigment cell formation in Ciona intestinalis. Nat. Commun. 2014, 5, 4830. [Google Scholar] [CrossRef] [Green Version]
- D’Aniello, E.; Pezzotti, M.R.; Locascio, A.; Branno, M. Onecut is a direct neural-specific transcriptional activator of Rx in Ciona intestinalis. Dev. Biol. 2011, 355, 358–371. [Google Scholar] [CrossRef] [Green Version]
- Hotta, K.; Mitsuhara, K.; Takahashi, H.; Inaba, K.; Oka, K.; Gojobori, T.; Ikeo, K. A web-based interactive developmental table for the ascidianCiona intestinalis, including 3D real-image embryo reconstructions: I. From fertilized egg to hatching larva. Dev. Dyn. 2007, 236, 1790–1805. [Google Scholar] [CrossRef]
- Brozovic, M.; Dantec, C.; Dardaillon, J.; Dauga, D.; Faure, E.; Gineste, M.; Louis, A.; Naville, M.; Nitta, K.R.; Piette, J.; et al. ANISEED 2017: Extending the integrated ascidian database to the exploration and evolutionary comparison of genome-scale datasets. Nucleic Acids Res. 2018, 46, D718–D725. [Google Scholar] [CrossRef]
- Zeller, R.W.; Weldon, D.S.; Pellatiro, M.A.; Cone, A.C. Optimized green fluorescent protein variants provide improved single cell resolution of transgene expression in ascidian embryos. Dev. Dyn. 2006, 235, 456–467. [Google Scholar] [CrossRef]
- Satou, Y.; Mineta, K.; Ogasawara, M.; Sasakura, Y.; Shoguchi, E.; Ueno, K.; Yamada, L.; Matsumoto, J.; Wasserscheid, J.; Dewar, K.; et al. Improved genome assembly and evidence-based global gene model set for the chordate Ciona intestinalis: New insight into intron and operon populations. Genome Biol. 2008, 9, R152. [Google Scholar] [CrossRef] [Green Version]
- Galiègue, S.; Jbilo, O.; Combes, T.; Bribes, E.; Carayon, P.; Le Fur, G.; Casellas, P. Cloning and Characterization of PRAX-1. J. Biol. Chem. 1999, 274, 2938–2952. [Google Scholar] [CrossRef] [Green Version]
- Abi-Rached, L.; Gilles, A.; Shiina, T.; Pontarotti, P.; Inoko, H. Evidence of en bloc duplication in vertebrate genomes. Nat. Genet. 2002, 31, 100–105. [Google Scholar] [CrossRef]
- Dehal, P.; Boore, J.L. Two rounds of whole genome duplication in the ancestral vertebrate. PLoS Biol. 2005, 3, e314. [Google Scholar] [CrossRef] [Green Version]
- Coppola, U.; Caccavale, F.; Scelzo, M.; Holland, N.D.; Ristoratore, F.; D’Aniello, S. Ran GTPase, an eukaryotic gene novelty, is involved in amphioxus mitosis. PLoS ONE 2018, 13, e0196930. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Qi, W.; Chen, G.; Feng, D.; Liu, J.; Ma, B.; Zhou, C.; Mu, C.; Zhang, W.; Chen, Q.; et al. Mitochondrial outer-membrane E3 ligase MUL1 ubiquitinates ULK1 and regulates selenite-induced mitophagy. Autophagy 2015, 11, 1216–1229. [Google Scholar] [CrossRef] [Green Version]
- Aligianis, I.A.; Johnson, C.A.; Gissen, P.; Chen, D.; Hampshire, D.; Hoffmann, K.; Maina, E.N.; Morgan, N.V.; Tee, L.; Morton, J.; et al. Mutations of the catalytic subunit of RAB3GAP cause Warburg Micro syndrome. Nat. Genet. 2005, 37, 221–224. [Google Scholar] [CrossRef]
- Aligianis, I.A.; Morgan, N.V.; Mione, M.; Johnson, C.A.; Rosser, E.; Hennekam, R.C.; Adams, G.; Trembath, R.C.; Pilz, D.T.; Stoodley, N.; et al. Mutation in Rab3 GTPase-activating protein (RAB3GAP) noncatalytic subunit in a kindred with Martsolf syndrome. Am. J. Hum. Genet. 2006, 78, 702–707. [Google Scholar] [CrossRef] [Green Version]
- Borck, G.; Wunram, H.; Steiert, A.; Volk, A.E.; Körber, F.; Roters, S.; Herkenrath, P.; Wollnik, B.; Morris-Rosendahl, D.J.; Kubisch, C. A homozygous RAB3GAP2 mutation causes Warburg Micro syndrome. Hum. Genet. 2011, 129, 45–50. [Google Scholar] [CrossRef]
- Takahashi, Y.; He, H.; Tang, Z.; Hattori, T.; Liu, Y.; Young, M.M.; Serfass, J.M.; Chen, L.; Gebru, M.; Chen, C.; et al. An autophagy assay reveals the ESCRT-III component CHMP2A as a regulator of phagophore closure. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, K.S.; Stolfi, A.; Levine, M.; Satou, Y. Gene regulatory networks underlying the compartmentalization of the Ciona central nervous system. Development 2009, 136, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Imai, J.H.; Meinertzhagen, I.A. Neurons of the ascidian larval nervous system inCiona intestinalis: II. Peripheral nervous system. J. Comp. Neurol. 2007, 501, 335–352. [Google Scholar] [CrossRef]
- Stolfi, A.; Ryan, K.; Meinertzhagen, I.A.; Christiaen, L. Migratory neuronal progenitors arise from the neural plate borders in tunicates. Nature 2015, 527, 371–374. [Google Scholar] [CrossRef] [Green Version]
- Cutter, A.D.; Agrawal, A.F. The evolutionary dynamics of operon distributions in eukaryote genomes. Genetics 2010, 185, 685–693. [Google Scholar] [CrossRef]
- Ryan, K.; Lu, Z.; Meinertzhagen, I.A. The peripheral nervous system of the ascidian tadpole larva: Types of neurons and their synaptic networks. J. Comp. Neurol. 2018, 526, 583–608. [Google Scholar] [CrossRef]
- Wagner, E.; Stolfi, A.; Gi Choi, Y.; Levine, M. Islet is a key determinant of ascidian palp morphogenesis. Development 2014, 141, 3084–3092. [Google Scholar] [CrossRef] [Green Version]
- Conaco, C.; Bassett, D.S.; Zhou, H.; Arcila, M.L.; Degnan, S.M.; Degnan, B.M.; Kosik, K.S. Functionalization of a protosynaptic gene expression network. Proc. Natl. Acad. Sci. USA 2012, 109 (Suppl. 1), 10612–10618. [Google Scholar] [CrossRef] [Green Version]
- Burkhardt, P.; Sprecher, S.G. Evolutionary origin of synapses and neurons—Bridging the gap. BioEssays 2017, 39, 1700024. [Google Scholar] [CrossRef]
- Martí-Solans, J.; Belyaeva, O.V.; Torres-Aguila, N.P.; Kedishvili, N.Y.; Albalat, R.; Cañestro, C. Coelimination and Survival in Gene Network Evolution: Dismantling the RA-Signaling in a Chordate. Mol. Biol. Evol. 2016, 33, 2401–2416. [Google Scholar] [CrossRef] [PubMed]
- Coppola, U.; Ristoratore, F.; Albalat, R.; D’Aniello, S. The evolutionary landscape of the Rab family in chordates. Cell. Mol. Life Sci. 2019, 76, 4117–4130. [Google Scholar] [CrossRef] [PubMed]
- Hoegg, S.; Brinkmann, H.; Taylor, J.S.; Meyer, A. Phylogenetic timing of the fish-specific genome duplication correlates with the diversification of teleost fish. J. Mol. Evol. 2004, 59, 190–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuraku, S.; Meyer, A. The evolution and maintenance of HOX gene clusters in vertebrates and the teleost-specific genome duplication. Int. J. Dev. Biol. 2009, 53, 765–773. [Google Scholar] [CrossRef] [Green Version]
- Grauel, M.K.; Maglione, M.; Reddy-Alla, S.; Willmes, C.G.; Brockmann, M.M.; Trimbuch, T.; Rosenmund, T.; Pangalos, M.; Vardar, G.; Stumpf, A.; et al. RIM-binding protein 2 regulates release probability by fine-tuning calcium channel localization at murine hippocampal synapses. Proc. Natl. Acad. Sci. USA 2016, 113, 11615–11620. [Google Scholar] [CrossRef] [Green Version]
- Brockmann, M.M.; Maglione, M.; Willmes, C.G.; Stumpf, A.; Bouazza, B.A.; Velasquez, L.M.; Grauel, M.K.; Beed, P.; Lehmann, M.; Gimber, N.; et al. RIM-BP2 primes synaptic vesicles via recruitment of Munc13-1 at hippocampal mossy fiber synapses. Elife 2019, 8. [Google Scholar] [CrossRef]
- Zhou, J.; Du, Y.-R.; Qin, W.-H.; Hu, Y.-G.; Huang, Y.-N.; Bao, L.; Han, D.; Mansouri, A.; Xu, G.-L. RIM-BP3 is a manchette-associated protein essential for spermiogenesis. Development 2009, 136, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Zeller, R.W. Computational analysis of Ciona intestinalis operons. Integr. Comp. Biol. 2010, 50, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.; Pym, E.C.G.; Tong, A.; Davis, G.W. Rab3-GAP controls the progression of synaptic homeostasis at a late stage of vesicle release. Neuron 2011, 69, 749–762. [Google Scholar] [CrossRef] [Green Version]
- Coppola, U.; Annona, G.; D’Aniello, S.; Ristoratore, F. Rab32 and Rab38 genes in chordate pigmentation: An evolutionary perspective. BMC Evol. Biol. 2016, 16. [Google Scholar] [CrossRef] [Green Version]
- Irimia, M.; Tena, J.J.; Alexis, M.S.; Fernandez-Minan, A.; Maeso, I.; Bogdanovic, O.; de la Calle-Mustienes, E.; Roy, S.W.; Gomez-Skarmeta, J.L.; Fraser, H.B. Extensive conservation of ancient microsynteny across metazoans due to cis-regulatory constraints. Genome Res. 2012, 22, 2356–2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coppola, U.; Olivo, P.; D’Aniello, E.; Johnson, C.J.; Stolfi, A.; Ristoratore, F. Rimbp, a New Marker for the Nervous System of the Tunicate Ciona robusta. Genes 2020, 11, 1006. https://doi.org/10.3390/genes11091006
Coppola U, Olivo P, D’Aniello E, Johnson CJ, Stolfi A, Ristoratore F. Rimbp, a New Marker for the Nervous System of the Tunicate Ciona robusta. Genes. 2020; 11(9):1006. https://doi.org/10.3390/genes11091006
Chicago/Turabian StyleCoppola, Ugo, Paola Olivo, Enrico D’Aniello, Christopher J. Johnson, Alberto Stolfi, and Filomena Ristoratore. 2020. "Rimbp, a New Marker for the Nervous System of the Tunicate Ciona robusta" Genes 11, no. 9: 1006. https://doi.org/10.3390/genes11091006
APA StyleCoppola, U., Olivo, P., D’Aniello, E., Johnson, C. J., Stolfi, A., & Ristoratore, F. (2020). Rimbp, a New Marker for the Nervous System of the Tunicate Ciona robusta. Genes, 11(9), 1006. https://doi.org/10.3390/genes11091006