Litter Size of Sheep (Ovis aries): Inbreeding Depression and Homozygous Regions




Abstract
:1. Introduction
2. Materials and Methods
2.1. Data
2.2. ROH Calling and Inbreeding
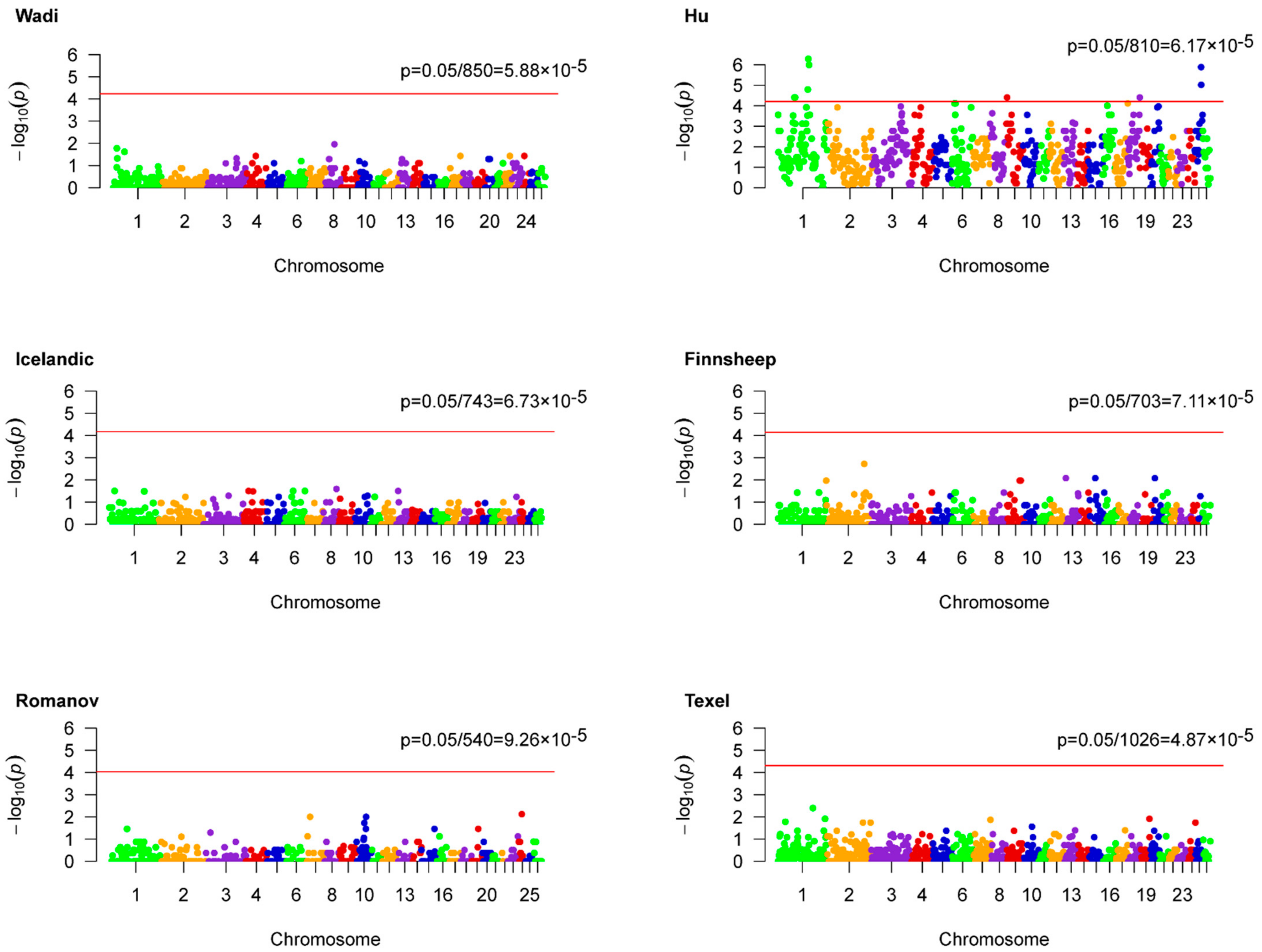
2.3. Genome-Wide ROH Hotspots Association Analysis
3. Results
3.1. Quality Control and Population Structure
3.2. Pattern of ROH and Inbreeding Level
3.3. Genome-Wide ROH Hotspots Association Analysis
4. Discussion
4.1. ROH and Inbreeding
4.2. Candidate Genes Likely Functioning in Prolificacy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, S.S.; Gao, L.; Xie, X.L.; Ren, Y.L.; Shen, Z.Q.; Wang, F.; Shen, M.; Eyϸórsdóttir, E.; Hallsson, J.H.; Kiseleva, T.; et al. Genome-wide association analyses highlight the potential for different genetic mechanisms for litter size among sheep breeds. Front. Genet. 2018, 9, 118. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; He, X.; Wang, F.; Zhong, Y.; Pan, L.; Wang, X.; Gan, S.; Di, R.; Chu, M. Luzhong mutton sheep: Inbreeding and selection signatures. J. Anim. Sci. Technol. 2020, 62, 777–789. [Google Scholar] [CrossRef]
- Howard, J.T.; Pryce, J.E.; Baes, C.; Maltecca, C. Inbreeding in the genomics era: Inbreeding, inbreeding depression, and management of genomic variability. J. Dairy Sci. 2017, 100, 6009–6024. [Google Scholar] [CrossRef] [PubMed]
- McQuillan, R.; Leutenegger, A.L.; Abdel-Rahman, R.; Franklin, C.S.; Pericic, M.; Barac-Lauc, L.; Smolej-Narancic, N.; Janicijevic, B.; Polasek, O.; Tenesa, A.; et al. Runs of homozygosity in European populations. Am. J. Hum. Genet. 2008, 83, 359–372. [Google Scholar] [CrossRef] [Green Version]
- Baes, C.F.; Makanjuola, B.O.; Miglior, F.; Marras, G.; Howard, J.T.; Fleming, A.; Maltecca, C. Symposium review: The genomic architecture of inbreeding: How homozygosity affects health and performance. J. Dairy Sci. 2019, 102, 2807–2817. [Google Scholar] [CrossRef]
- Drobik, W.; Martyniuk, E. Inbreeding and its impact on the prolific Polish Olkuska sheep population. Small Rumin. Res. 2016, 137, 28–33. [Google Scholar] [CrossRef]
- Vostry, L.; Milerski, M.; Schmidova, J.; Vostra-Vydrova, H. Genetic diversity and effect of inbreeding on litter size of the Romanov sheep. Small Rumin. Res. 2018, 168, 25–31. [Google Scholar] [CrossRef]
- Tam, V.; Patel, N.; Turcotte, M.; Bossé, Y.; Paré, G.; Meyre, D. Benefits and limitations of genome-wide association studies. Nat. Rev. Genet. 2019, 20, 467–484. [Google Scholar] [CrossRef]
- Peripolli, E.; Munari, D.; Silva, M.; Lima, A.; Irgang, R.; Baldi, F. Runs of homozygosity: Current knowledge and applications in livestock. Anim. Genet. 2017, 48, 255–271. [Google Scholar] [CrossRef]
- Grilz-Seger, G.; Druml, T.; Neuditschko, M.; Mesaric, M.; Cotman, M.; Brem, G. Analysis of ROH patterns in the Noriker horse breed reveals signatures of selection for coat color and body size. Anim. Genet. 2019, 50, 334–346. [Google Scholar] [CrossRef]
- Cesarani, A.; Gaspa, G.; Pauciullo, A.; Degano, L.; Vicario, D.; Macciotta, N.P.P. Genome-wide analysis of homozygosity regions in european simmental bulls. J. Anim. Breed Genet. 2021, 138, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Abdoli, R.; Zamani, P.; Mirhoseini, S.Z.; Ghavi Hossein-Zadeh, N.; Nadri, S. A review on prolificacy genes in sheep. Reprod. Domest. Anim. 2016, 51, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Li, M.Y.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Feng, Z.; Teresky, A.K.; Levine, A.J. p53 regulates maternal reproduction through LIF. Nature 2007, 450, 721–724. [Google Scholar] [CrossRef]
- Yuan, Z.; Zhang, J.; Li, W.; Wang, W.; Li, F.; Yue, X. Association of polymorphisms in candidate genes with the litter size in two sheep breeds. Animals 2019, 9, 958. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Peng, W.F.; Hu, X.J.; Zhao, Y.X.; Lv, F.H.; Yang, J. Global genomic diversity and conservation priorities for domestic animals are associated with the economies of their regions of origin. Sci. Rep. 2018, 8, 11677. [Google Scholar] [CrossRef] [Green Version]
- Norberg, E.; Sørensen, A.C. Inbreeding trend and inbreeding depression in the Danish populations of Texel, Shropshire, and Oxford Down. J. Anim. Sci. 2007, 85, 299–304. [Google Scholar] [CrossRef]
- Liu, B. Animal Breeding; China Agriculture Press: Beijing, China, 2007; p. 146. [Google Scholar]
- Haresign, W. The physiological basis for variation in ovulation rate and litter size in sheep: A review. Livest Prod. Sci. 1985, 13, 3–20. [Google Scholar] [CrossRef]
- Hanrahan, J.; Quirke, J. Contribution of variation in ovulation rate and embryo survival to within breed variation in litter size. In Genetics of Reproduction in Sheep; Land, R.B., Robinson, D.W., Eds.; Garden City Press Ltd.: Letchworth Garden City, Hertfordshire, UK, 1984. [Google Scholar]
- Hu, W.; Feng, Z.; Atwal, G.S.; Levine, A.J. p53: A new player in reproduction. Cell Cycle 2008, 7, 848–852. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Mei, S.; Zhang, X.; Peng, X.; Yang, J.; Zhu, L.; Zhou, J.; Wu, H.; Wang, L.; Hua, L.; et al. Transcription factor C/EBPβ and 17β-estradiol promote transcription of the porcine p53 gene. Int. J. Biochem. Cell Biol. 2014, 47, 76–82. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Bai, L.; Hou, J.; Zhao, H.; Peng, J.; Song, Y.; Wang, J.; Cao, B. Molecular cloning, tissue expression and SNP analysis in the goat nerve growth factor gene. Mol. Biol. Rep. 2013, 40, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Naicy, T.; Venkatachalapathy, R.T.; Aravindakshan, T.V.; Radhika, G.; Raghavan, K.C.; Mini, M.; Shyama, K. Nerve Growth Factor gene ovarian expression, polymorphism identification, and association with litter size in goats. Theriogenology 2016, 86, 2172–2178.e2173. [Google Scholar] [CrossRef]
- Naicy, T.; Venkatachalapathy, R.T.; Aravindakshan, T.V.; Raghavan, K.C.; Mini, M.; Shyama, K. Relative abundance of tissue mRNA and association of the single nucleotide polymorphism of the goat NGF gene with prolificacy. Anim. Reprod. Sci. 2016, 173, 42–48. [Google Scholar] [CrossRef]
- Fabi, F.; Asselin, E. Expression, activation, and role of AKT isoforms in the uterus. Reproduction 2014, 148, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Artini, P.G.; Tatone, C.; Sperduti, S.; D’Aurora, M.; Franchi, S.; Di Emidio, G.; Ciriminna, R.; Vento, M.; Di Pietro, C.; Stuppia, L.; et al. Cumulus cells surrounding oocytes with high developmental competence exhibit down-regulation of phosphoinositol 1,3 kinase/protein kinase B (PI3K/AKT) signalling genes involved in proliferation and survival. Hum. Reprod. 2017, 32, 2474–2484. [Google Scholar] [CrossRef]
- Hall, S.E.; Upton, R.M.O.; McLaughlin, E.A.; Sutherland, J.M. Phosphoinositide 3-kinase/protein kinase B (PI3K/AKT) and Janus kinase/signal transducer and activator of transcription (JAK/STAT) follicular signalling is conserved in the mare ovary. Reprod. Fertil. Dev. 2018, 30, 624–633. [Google Scholar] [CrossRef]
- Zeleznik, A.J.; Saxena, D.; Little-Ihrig, L. Protein kinase B is obligatory for follicle-stimulating hormone-induced granulosa cell differentiation. Endocrinology 2003, 144, 3985–3994. [Google Scholar] [CrossRef] [Green Version]
- Kalous, J.; Solc, P.; Baran, V.; Kubelka, M.; Schultz, R.M.; Motlik, J. PKB/AKT is involved in resumption of meiosis in mouse oocytes. Biol. Cell 2006, 98, 111–123. [Google Scholar] [CrossRef]
- Hsueh, A.J.W.; Kawamura, K.; Cheng, Y.; Fauser, B.C.J.M. Intraovarian Control of Early Folliculogenesis. Endocr. Rev. 2015, 36, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier, A.; Goupil, A.S.; Le Gac, F.; Lareyre, J.J. A promoter fragment of the sycp1 gene is sufficient to drive transgene expression in male and female meiotic germ cells in zebrafish. Biol. Reprod. 2013, 89, 89. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.L.; Zhang, C.; Hao, J.; Wang, X.L.; Ming, J.; Mi, L.; Na, J.; Hu, X.; Wang, Y. DPPA2/4 and SUMO E3 ligase PIAS4 opposingly regulate zygotic transcriptional program. PLoS Biol. 2019, 17, e3000324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Iaco, A.; Coudray, A.; Duc, J.; Trono, D. DPPA2 and DPPA4 are necessary to establish a 2C-like state in mouse embryonic stem cells. EMBO Rep. 2019, 20, e47382. [Google Scholar] [CrossRef]
- Eckersley-Maslin, M.; Alda-Catalinas, C.; Blotenburg, M.; Kreibich, E.; Krueger, C.; Reik, W. Dppa2 and Dppa4 directly regulate the Dux-driven zygotic transcriptional program. Genes Dev. 2019, 33, 194–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mochizuki, K.; Tando, Y.; Sekinaka, T.; Otsuka, K.; Hayashi, Y.; Kobayashi, H.; Kamio, A.; Ito-Matsuoka, Y.; Takehara, A.; Kono, T.; et al. SETDB1 is essential for mouse primordial germ cell fate determination by ensuring BMP signaling. Development 2018, 145, dev164160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madan, B.; Madan, V.; Weber, O.; Tropel, P.; Blum, C.; Kieffer, E.; Viville, S.; Fehling, H.J. The pluripotency-associated gene Dppa4 is dispensable for embryonic stem cell identity and germ cell development but essential for embryogenesis. Mol. Cell Biol. 2009, 29, 3186–3203. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Cui, J.; Zhao, C.; Zhou, L.; Guo, X.; Shen, R.; Zhang, J.; Ling, X. Differential expression of microRNAs in 2-cell and 4-cell mouse embryos. Zygote 2014, 22, 455–461. [Google Scholar] [CrossRef]
- Lamm, K.Y.B.; Johnson, M.L.; Baker Phillips, J.; Muntifering, M.B.; James, J.M.; Jones, H.N.; Redline, R.W.; Rokas, A.; Muglia, L.J. Inverted formin 2 regulates intracellular trafficking, placentation, and pregnancy outcome. eLife 2018, 7, e31150. [Google Scholar] [CrossRef] [Green Version]
- Landeira, D.; Bagci, H.; Malinowski, A.R.; Brown, K.E.; Soza-Ried, J.; Feytout, A.; Webster, Z.; Ndjetehe, E.; Cantone, I.; Asenjo, H.G.; et al. Jarid2 coordinates nanog expression and PCP/Wnt signaling required for efficient ESC differentiation and early embryo development. Cell Rep. 2015, 12, 573–586. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Chen, X.; Zhao, Y.; Gao, R.; Geng, Y.; Ding, Y.; Liu, X.; Wang, Y.; He, J. The homologous genes Vangl1 and Vangl2 are required for embryo implantation in the uterus of mice during early pregnancy. Gene 2015, 555, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Ju Lee, H.; Bartsch, D.; Xiao, C.; Guerrero, S.; Ahuja, G.; Schindler, C.; Moresco, J.J.; Yates, J.R., 3rd; Gebauer, F.; Bazzi, H.; et al. A post-transcriptional program coordinated by CSDE1 prevents intrinsic neural differentiation of human embryonic stem cells. Nat. Commun. 2017, 8, 1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pergande, M.; Motameny, S.; Özdemir, Ö.; Kreutzer, M.; Wang, H.; Daimagüler, H.S.; Becker, K.; Karakaya, M.; Ehrhardt, H.; Elcioglu, N.; et al. The genomic and clinical landscape of fetal akinesia. Genet. Med. 2020, 22, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Cheung, S.; Parrella, A.; Rosenwaks, Z.; Palermo, G.D. Genetic and epigenetic profiling of the infertile male. PLoS ONE 2019, 14, e0214275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arafat, M.; Har-Vardi, I.; Harlev, A.; Levitas, E.; Zeadna, A.; Abofoul-Azab, M.; Dyomin, V.; Sheffield, V.C.; Lunenfeld, E.; Huleihel, M.; et al. Mutation in TDRD9 causes non-obstructive azoospermia in infertile men. J. Med. Genet. 2017, 54, 633–639. [Google Scholar] [CrossRef]
- Zhao, W.; Quansah, E.; Yuan, M.; Gou, Q.; Mengal, K.; Li, P.; Wu, S.; Xu, C.; Yi, C.; Cai, X. Region-specific gene expression in the epididymis of Yak. Theriogenology 2019, 139, 132–146. [Google Scholar] [CrossRef]
- Rival, C.M.; Xu, W.; Shankman, L.S.; Morioka, S.; Arandjelovic, S.; Lee, C.S.; Wheeler, K.M.; Smith, R.P.; Haney, L.B.; Isakson, B.E.; et al. Phosphatidylserine on viable sperm and phagocytic machinery in oocytes regulate mammalian fertilization. Nat. Commun. 2019, 10, 4456. [Google Scholar] [CrossRef] [Green Version]
- Kazarian, E.; Son, H.; Sapao, P.; Li, W.; Zhang, Z.; Strauss, J.F.; Teves, M.E. SPAG17 is required for male germ cell differentiation and fertility. Int. J. Mol. Sci. 2018, 19, 1252. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, S.; Tang, Q.; Jiang, E.; Wang, K.; Lan, X.; Pan, C. Goat sperm associated antigen 17 protein gene (SPAG17): Small and large fragment genetic variation detection, association analysis, and mRNA expression in gonads. Genomics 2020, 112, 5115–5121. [Google Scholar] [CrossRef]
- Han, Y.; Peñagaricano, F. Unravelling the genomic architecture of bull fertility in Holstein cattle. BMC Genet. 2016, 17, 143. [Google Scholar] [CrossRef] [Green Version]
- Woods, L.C., III; Li, Y.; Ding, Y.; Liu, J.; Reading, B.J.; Fuller, S.A.; Song, J. DNA methylation profiles correlated to striped bass sperm fertility. BMC Genom. 2018, 19, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, R.; Assis, J.; Nogueira, A.; Pereira, C.; Coelho, S.; Brandão, M.; Dias, J.; Alves, S.; Pereira, D.; Medeiros, R. Pharmacogenomics in epithelial ovarian cancer first-line treatment outcome: Validation of GWAS-associated NRG3 rs1649942 and BRE rs7572644 variants in an independent cohort. Pharm. J. 2019, 19, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.S.; Vilas Boas, D.B.; Gigliotti, C.B.; Salvadori, D.M. Association among XRCC1, XRCC3, and BLHX gene polymorphisms and chromosome instability in lymphocytes from patients with endometriosis and ovarian cancer. Genet. Mol. Res. 2014, 13, 636–648. [Google Scholar] [CrossRef] [PubMed]
- Mirza, Z.; Abdel-Dayem, U. Uncovering potential roles of differentially expressed genes, upstream regulators, and canonical pathways in endometriosis using an in silico genomics approach. Diagnostics 2020, 10, 416. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhang, A.; Yu, F.; Gao, J.; Liu, Y.; Yu, C.; Zhou, H.; Xu, C. Label-free proteomics uncovers energy metabolism and focal adhesion regulations responsive for endometrium receptivity. J. Proteome Res. 2015, 14, 1831–1842. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | SNP for ROH Calling | Average Litter Size (Mean ± SD) † | CV (%) | ||
|---|---|---|---|---|---|
| Case | Control | Total | |||
| Wadi | 512,178 | 2.28 ± 0.50 A (n = 77) | 1.87 ± 0.10 B (n = 28) | 1.99 ± 0.65 (n = 100) | 32.81 |
| Hu | 503,422 | 2.10 ± 0.24 A (n = 58) | 1.00 ± 0.00 B (n = 13) | 1.90 ± 0.47 (n = 71) | 24.87 |
| Icelandic | 443,125 | 2.29 ± 0.19 A (n = 8) | 1.66 ± 0.10 B (n = 15) | 1.88 ± 0.33 (n = 23) | 17.83 |
| Finnsheep | 492,165 | 3.11 ± 0.45 A (n = 28) | 1.66 ± 0.33 B (n = 9) | 2.76 ± 0.76 (n = 37) | 27.55 |
| Romanov | 465,794 | 2.97 ± 0.29 A (n = 29) | 1.53 ± 0.25 B (n = 9) | 2.63 ± 0.68 (n = 38) | 25.90 |
| Texel | 475,999 | 1.87 ± 0.35 A (n = 28) | 1.15 ± 0.10 B (n = 11) | 1.66 ± 0.45 (n = 39) | 26.86 |
| ROH Feature | Breed † | |||||
|---|---|---|---|---|---|---|
| Hu | Wadi | Icelandic | Finnsheep | Romanov | Texel | |
| Number | ||||||
| Average | 29.86 C | 21.26 C | 81.96 A | 45.38 B | 43.71 B | 85.03 A |
| SD | 20.90 | 11.05 | 29.07 | 15.76 | 9.01 | 14.42 |
| Maximum | 82 | 67 | 125 | 91 | 60 | 125 |
| Total length (Mb) | ||||||
| Average | 247.9 C | 84.97 C | 306.98 A | 173.51 B | 128.15 B | 258.17 A |
| SD | 388.94 | 140.37 | 175.46 | 131.91 | 41.40 | 47.75 |
| Maximum | 1580.38 | 733.08 | 916.21 | 541.73 | 223.37 | 384.18 |
| Average length (Mb) | ||||||
| Average | 4.89 B | 3.01 B | 3.58 A | 3.48 A | 2.91 A | 3.06 A |
| SD | 5.42 | 2.64 | 1.17 | 1.36 | 0.59 | 0.41 |
| Maximum | 22.26 | 14.37 | 7.33 | 7.06 | 4.32 | 3.97 |
| FROH | Estimate ± SE | |||||
|---|---|---|---|---|---|---|
| Wadi | Hu | Icelandic | Finnsheep | Romanov | Texel | |
| FROH1–4 | 0.12 ± 0.16 | −0.22 ± 0.13 | −0.03 ± 0.04 | −0.20 ± 0.20 | 0.35 ± 0.23 | −0.02 ± 0.08 |
| FROH4–8 | −0.15 ± 0.14 | −0.16 ** ± 0.05 | −0.01 ± 0.05 | −0.08 ± 0.12 | −0.06 ± 0.19 | −0.11 ± 0.10 |
| FROH > 8 | −0.01 ± 0.01 | −0.02 ** ± 0.00 | 0.01 ± 0.01 | −0.01 ± 0.03 | −0.11 ± 0.11 | 0.04 ± 0.06 |
| All | −0.01 ± 0.01 | −0.02 ** ± 0.00 | 0.00 ± 0.01 | −0.02 ± 0.03 | −0.02 ± 0.07 | 0.00 ± 0.04 |
| ROH Hotspot | Region † | Length (kb) | N_SNP | N_Gene | p-Value |
|---|---|---|---|---|---|
| S32 | 1:167605062-167736007 | 130.946 | 24 | 0 | 5.14 × 10−7 |
| S325 | 1:171893678-173273898 | 1380.221 | 269 | 5 | 1.01 × 10−6 |
| S736 | 25:34831274-34920313 | 89.04 | 20 | 1 | 1.29 × 10−6 |
| S599 | 25:35553072-36870587 | 1317.516 | 275 | 2 | 9.48 × 10−6 |
| S125 | 1:164718392-165322668 | 604.277 | 107 | 1 | 1.60 × 10−5 |
| S414 | 18:66738545-68488609 | 1750.065 | 299 | 30 | 3.91 × 10−5 |
| S378 | 9:5081783-5791789 | 710.007 | 161 | 1 | 3.91 × 10−5 |
| S312 | 1:94177534-94744969 | 567.436 | 129 | 3 | 3.91 × 10−5 |
| S311 | 1:90921006-92192152 | 1271.147 | 227 | 13 | 3.91 × 10−5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, L.; He, X.; Wang, X.; Di, R.; Chu, M. Litter Size of Sheep (Ovis aries): Inbreeding Depression and Homozygous Regions. Genes 2021, 12, 109. https://doi.org/10.3390/genes12010109
Tao L, He X, Wang X, Di R, Chu M. Litter Size of Sheep (Ovis aries): Inbreeding Depression and Homozygous Regions. Genes. 2021; 12(1):109. https://doi.org/10.3390/genes12010109
Chicago/Turabian StyleTao, Lin, Xiaoyun He, Xiangyu Wang, Ran Di, and Mingxing Chu. 2021. "Litter Size of Sheep (Ovis aries): Inbreeding Depression and Homozygous Regions" Genes 12, no. 1: 109. https://doi.org/10.3390/genes12010109
APA StyleTao, L., He, X., Wang, X., Di, R., & Chu, M. (2021). Litter Size of Sheep (Ovis aries): Inbreeding Depression and Homozygous Regions. Genes, 12(1), 109. https://doi.org/10.3390/genes12010109