Impact of Genetic Variability in ACE2 Expression on the Evolutionary Dynamics of SARS-CoV-2 Spike D614G Mutation



Abstract
:1. Introduction
2. Materials and Methods
2.1. Frequency Analysis of D614 and G614 Variants across Geographic Regions
2.2. ACE2 Expresion Levels in Different Genetic Human Populations
2.3. Polymorphism Analysis of 3′UTR, 5′UTR, and Promoter Regions in ACE2
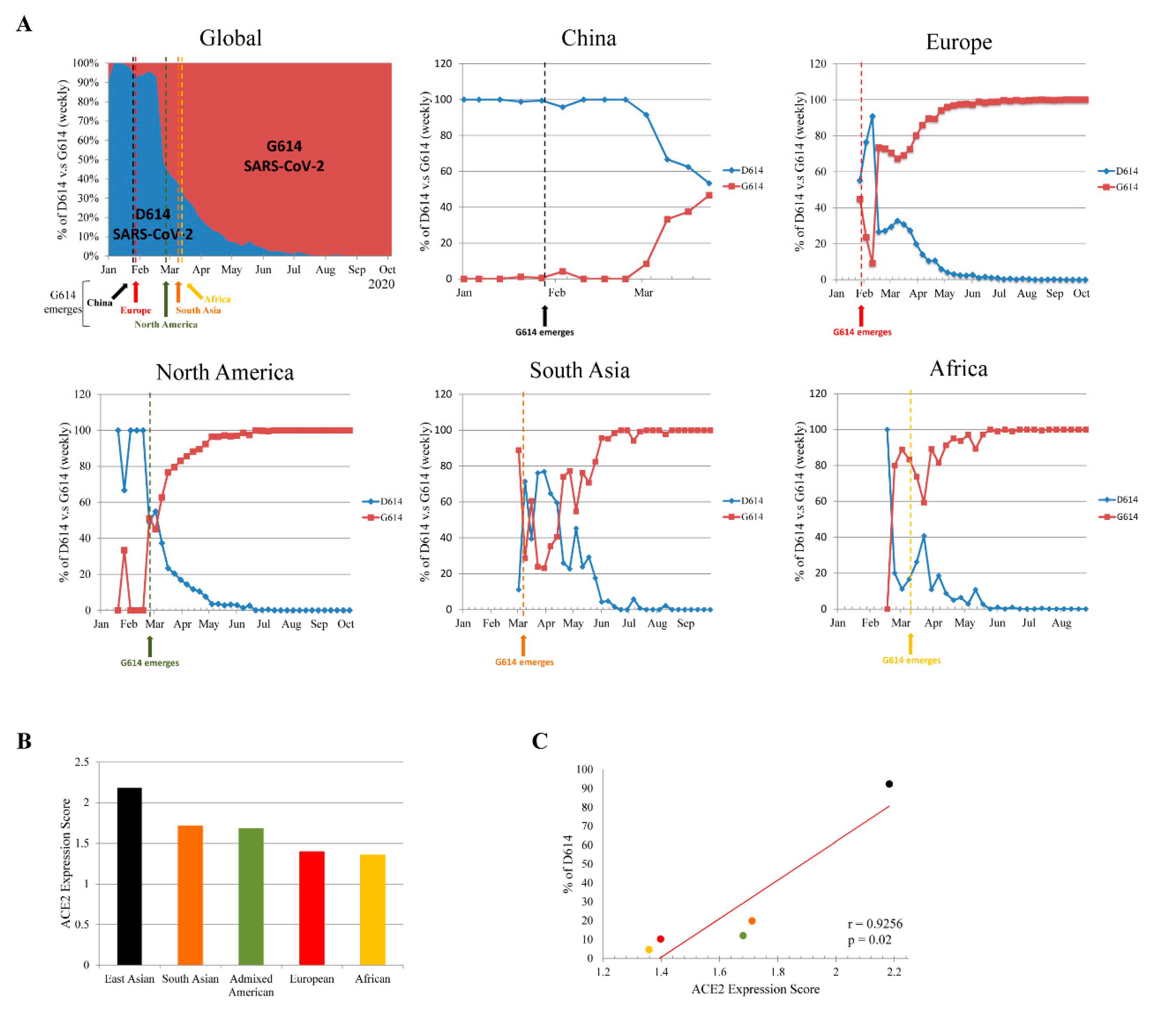
3. Results
3.1. Dynamic Tracking of SARS-CoV-2 S Glycoprotein D614G Mutation
3.2. Genetic Variability in ACE2 Expression
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundstrom, K. Coronavirus Pandemic-Therapy and Vaccines. Biomedicines 2020, 8, 109. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Wang, W.H.; Urbina, A.N.; Tseng, S.P.; Lu, P.L.; Chen, Y.H.; Yu, M.L.; Wang, S.F. Importation of SARS-CoV-2 infection leads to major COVID-19 epidemic in Taiwan. Int. J. Infect. Dis. 2020, 97, 240–244. [Google Scholar] [CrossRef]
- Li, X.; Wang, W.; Zhao, X.; Zai, J.; Zhao, Q.; Li, Y.; Chaillon, A. Transmission dynamics and evolutionary history of 2019-nCoV. J. Med. Virol. 2020, 92, 501–511. [Google Scholar] [CrossRef]
- Kim, J.S.; Jang, J.H.; Kim, J.M.; Chung, Y.S.; Yoo, C.K.; Han, M.G. Genome-Wide Identification and Characterization of Point Mutations in the SARS-CoV-2 Genome. Osong Public Health Res. Perspect. 2020, 11, 101–111. [Google Scholar] [CrossRef]
- Forster, P.; Forster, L.; Renfrew, C.; Forster, M. Phylogenetic network analysis of SARS-CoV-2 genomes. Proc. Natl. Acad. Sci. USA 2020, 117, 9241–9243. [Google Scholar] [CrossRef] [Green Version]
- Laamarti, M.; Alouane, T.; Kartti, S.; Elfihri, M.W.; Hakmi, M.; Essabbar, A.; Laamart, M.; Hlali, H.; Allam, L.; Hafidi, N.E.; et al. Large scale genomic analysis of 3067 SARS CoV-2 genomes reveals a clonal geo-distribution and a rich genetic variations of hotspots mutations. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zhang, L.; Jackson, C.B.; Mou, H.; Ojha, A.; Peng, H.; Quinlan, B.D.; Rangarajan, E.S.; Pan, A.; Vanderheiden, A.; Suthar, M.S.; et al. SARS-CoV-2 spike-protein D614G mutation increases virion spike density and infectivity. Nat. Commun. 2020, 11, 6013. [Google Scholar] [CrossRef]
- Yurkovetskiy, L.; Wang, X.; Pascal, K.E.; Tomkins-Tinch, C.; Nyalile, T.P.; Wang, Y.; Baum, A.; Diehl, W.E.; Dauphin, A.; Carbone, C.; et al. Structural and Functional Analysis of the D614G SARS-CoV-2 Spike Protein Variant. Cell 2020, 183, 739–751. [Google Scholar] [CrossRef]
- Plante, J.A.; Liu, Y.; Liu, J.; Xia, H.; Johnson, B.A.; Lokugamage, K.G.; Zhang, X.; Muruato, A.E.; Zou, J.; Fontes-Garfias, C.R.; et al. Spike mutation D614G alters SARS-CoV-2 fitness. Nature 2020. [Google Scholar] [CrossRef] [PubMed]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B.; et al. Tracking Changes in SARS-CoV-2 Spike: Evidence that D614G Increases Infectivity of the COVID-19 Virus. Cell 2020, 182, 812–827. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.J.; Chiba, S.; Halfmann, P.; Ehre, C.; Kuroda, M.; Dinnon, K.H., 3rd; Leist, S.R.; Schafer, A.; Nakajima, N.; Takahashi, K.; et al. SARS-CoV-2 D614G variant exhibits efficient replication ex vivo and transmission in vivo. Science 2020. [Google Scholar] [CrossRef] [PubMed]
- Coutard, B.; Valle, C.; de Lamballerie, X.; Canard, B.; Seidah, N.G.; Decroly, E. The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antivir. Res. 2020, 176, 104742. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Wu, L.; Niu, S.; Song, C.; Zhang, Z.; Lu, G.; Qiao, C.; Hu, Y.; Yuen, K.Y.; et al. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2. Cell 2020, 181, 894–904. [Google Scholar] [CrossRef]
- Becerra-Flores, M.; Cardozo, T. SARS-CoV-2 viral spike G614 mutation exhibits higher case fatality rate. Int. J. Clin. Pract. 2020, 74, e13525. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Fernandez, L.; Sawalha, A.H. Genetic variability in the expression of the SARS-CoV-2 host cell entry factors across populations. Genes Immun. 2020, 21, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Laboratory, Data Analysis and Coordinating Center (LDACC)—Analysis Working Group; Statistical Methods Groups—Analysis Working Group; Enhancing GTEx (eGTEx) Groups; NIH Common Fund; NIH/NCI; NIH/NHGRI; NIH/NIMH; NIH/NIDA; Biospecimen Collection Source Site—NDRI; Biospecimen Collection Source Site—RPCI; et al. Genetic effects on gene expression across human tissues. Nature 2017, 550, 204–213. [Google Scholar] [CrossRef]
- Genomes Project, C.; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar]
- Chen, J.; Jiang, Q.; Xia, X.; Liu, K.; Yu, Z.; Tao, W.; Gong, W.; Han, J.J. Individual variation of the SARS-CoV-2 receptor ACE2 gene expression and regulation. Aging Cell 2020, 19, e13168. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, L.; Feng, Z.; Wan, S.; Huang, P.; Sun, X.; Wen, F.; Huang, X.; Ning, G.; Wang, W. Comparative genetic analysis of the novel coronavirus (2019-nCoV/SARS-CoV-2) receptor ACE2 in different populations. Cell Discov. 2020, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, J.; Ye, G.; Shi, K.; Wan, Y.; Luo, C.; Aihara, H.; Geng, Q.; Auerbach, A.; Li, F. Structural basis of receptor recognition by SARS-CoV-2. Nature 2020, 581, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Rs ID | Ref. Allele | Variant | Function Class (a.a. Change) | Global | African | Asian | European | American | Other | Potential Functional Effect | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| rs112171234 | T | G | Intron | 0.058 | 0.2047 | 0 | 0.00076 | 0.0245 | 0.025 | Higher expression | [22] |
| rs4646127 | A | G | Intron | 0.75103 | 0.8009 | 0.9965 | 0.63994 | 0.767 | 0.701 | Higher expression | [22] |
| rs4830974 | G | A | 5′ distal regions (CLTRN intron) † | 0.65138 | 0.73245 | 0.9396 | 0.50745 | 0.713 | 0.564 | Higher expression | [22] |
| rs6632704 | C | A | 5′ distal regions (CLTRN intron) † | 0.58884 | 0.5088 | 0.9415 | 0.50677 | 0.699 | 0.535 | Higher expression | [22] |
| rs2158082 | G | A | 5′ distal regions (CLTRN intron) † | 0.72834 | 0.95155 | 0.9955 | 0.50595 | 0.746 | 0.586 | Higher expression | [22] |
| rs1996225 | C | T | 5′ distal regions (CA5B intron) † | 0.51673 | 0.57974 | 0.74372 | 0.41528 | 0.60129 | 0.4926 | Higher expression | [22] |
| rs5936029 | C | T | 5′ distal regions (CA5B intron) † | 0.70741 | 0.88315 | 0.9875 | 0.50442 | 0.726 | 0.587 | Higher expression | [22] |
| rs4830983 | A | C | 5′ distal regions (CA5B intron) † | 0.72881 | 0.95195 | 0.9885 | 0.51156 | 0.746 | 0.585 | Higher expression | [22] |
| rs5936011 | C | T | 5′ distal regions (CA5B intron) † | 0.70679 | 0.88175 | 0.9875 | 0.50893 | 0.734 | 0.574 | Higher expression | [22] |
| rs4060 | C | A | 5′ distal regions (CA5B intron) † | 0.63257 | 0.6688 | 0.9395 | 0.50713 | 0.708 | 0.548 | Higher expression | [22] |
| rs6629110 | T | C | 5′ distal regions (Intergene region) † | 0.57021 | 0.43585 | 0.9575 | 0.50413 | 0.6875 | 0.526 | Higher expression | [22] |
| rs143695310 | T | A | 5′ distal regions (Intergene region) † | 0.0174 | 0.0028 | 0 | 0.03528 | 0.022 | 0.034 | Higher expression | [22] |
| rs4646112 | C | T | Promoter | 0.01921 | 0.06575 | 0 | 0.00012 | 0.013 | 0.009 | Not characterized | - |
| rs4646113 | G | A | Promoter | 0.02078 | 0.07485 | 0 | 0.00058 | 0.0055 | 0.013 | Not characterized | - |
| rs4646114 | C | T | Promoter | 0.01959 | 0.0667 | 0 | 0.00012 | 0.013 | 0.009 | Not characterized | - |
| rs111572878 | T | C | Promoter | 0.05531 | 0.20065 | 0 | 0.00073 | 0.0155 | 0.023 | Not characterized | - |
| rs113208650 | C | T | Promoter | 0.02839 | 0.1031 | 0 | 0.00065 | 0.0055 | 0.014 | Not characterized | - |
| rs185721534 | A | G | Promoter | 0.02512 | 0.0905 | 0 | 0.00012 | 0.009 | 0.009 | Not characterized | - |
| rs113208650 | C | T | Promoter | 0.02839 | 0.1031 | 0 | 0.00065 | 0.0055 | 0.014 | Not characterized | - |
| rs113009615 | A | delA | Promoter | 0.05618 | 0.20125 | 0 | 0.00077 | 0.0165 | 0.023 | Not characterized | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.-W.; Miller, S.O.; Yen, C.-H.; Wang, S.-F. Impact of Genetic Variability in ACE2 Expression on the Evolutionary Dynamics of SARS-CoV-2 Spike D614G Mutation. Genes 2021, 12, 16. https://doi.org/10.3390/genes12010016
Huang S-W, Miller SO, Yen C-H, Wang S-F. Impact of Genetic Variability in ACE2 Expression on the Evolutionary Dynamics of SARS-CoV-2 Spike D614G Mutation. Genes. 2021; 12(1):16. https://doi.org/10.3390/genes12010016
Chicago/Turabian StyleHuang, Szu-Wei, Sorin O. Miller, Chia-Hung Yen, and Sheng-Fan Wang. 2021. "Impact of Genetic Variability in ACE2 Expression on the Evolutionary Dynamics of SARS-CoV-2 Spike D614G Mutation" Genes 12, no. 1: 16. https://doi.org/10.3390/genes12010016
APA StyleHuang, S. -W., Miller, S. O., Yen, C. -H., & Wang, S. -F. (2021). Impact of Genetic Variability in ACE2 Expression on the Evolutionary Dynamics of SARS-CoV-2 Spike D614G Mutation. Genes, 12(1), 16. https://doi.org/10.3390/genes12010016