Metagenome Analysis of a Hydrocarbon-Degrading Bacterial Consortium Reveals the Specific Roles of BTEX Biodegraders


Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling and Study Site Description
2.2. Enrichment Culture and Growth Conditions
2.3. DNA Extraction
2.4. Sequencing of Bacterial 16S rRNA Gene
2.5. Processing of the 16S rRNA Gene Data
2.6. Metagenome Sequencing, Assembly and Analysis
2.7. Identification of CDSs Involved in BTEX Biodegradation
2.8. Greenhouse Biodegradation Experiment
2.9. Geochemical Analysis of Biodegradation
2.9.1. Extraction of Residual Hydrocarbons
2.9.2. Molecular Analysis of Biodegradation
2.10. Statistical Analysis
3. Results
3.1. Bacterial Diversity in the Consortium
3.2. Activation of BTEX
3.2.1. Benzene and Ethylbenzene
3.2.2. Toluene and Xylenes
3.3. Central Metabolism of BTEX
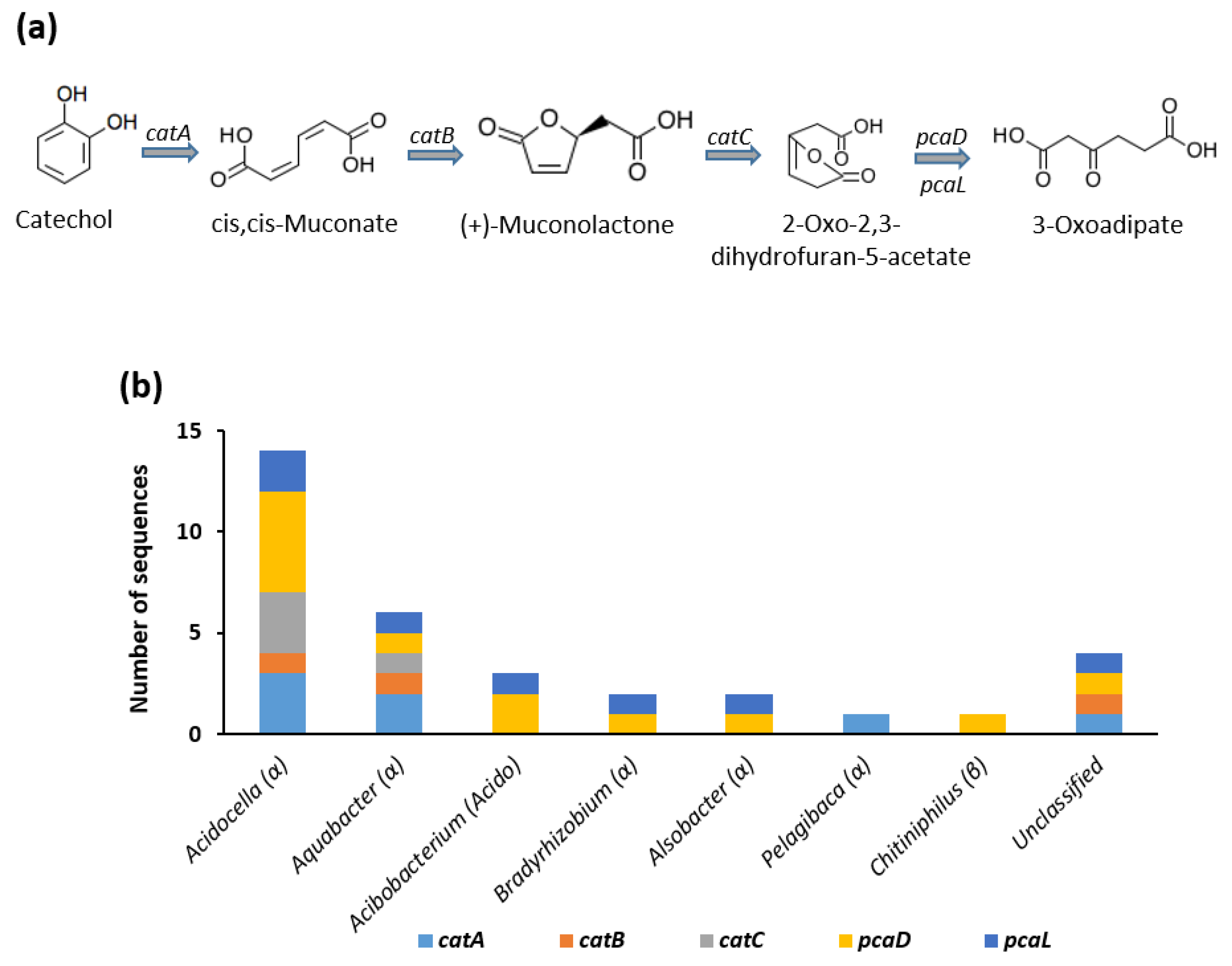
3.3.1. Ortho-Cleavage of Catechol
3.3.2. Meta-Cleavage of Catechol, Methylcatechol, and Ethylcatechol
3.4. Analysis of Residual Total Petroleum Hydrocarbons
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dean, B.J. Recent findings on the genetic toxicology of benzene, toluene, xylenes and phenols. Mutat. Res. Genet. Toxicol. 1985, 154, 153–181. [Google Scholar] [CrossRef]
- Harvey, A.N.; Snape, I.; Siciliano, S.D. Validating potential toxicity assays to assess petroleum hydrocarbon toxicity in polar soil. Environ. Toxicol. Chem. 2011, 31, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Garr, A.L.; Laramore, S.; Krebs, W. Toxic effects of oil and dispersant on marine microalgae. Bull. Environ. Contam. Toxicol. 2014, 93, 654–659. [Google Scholar] [CrossRef] [PubMed]
- McKee, R.H.; White, R. The mammalian toxicological hazards of petroleum-derived substances: An overview of the petroleum industry response to the high production volume challenge program. Int. J. Toxicol. 2013, 33, 4S–16S. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, K.; Matsushita, H.; Sakabe, H.; Takemoto, K. Carcinogenicity of benzyl chloride, benzal chloride, benzotrichloride and benzoyl chloride in mice by skin application. Gan 1981, 72, 655–664. [Google Scholar]
- IARC. Overall Evaluations of Carcinogenicity: An Updating of IARC Monographs Volumes 1 to 42; International Agency for Research on Cancer: Lyon, France, 1987. [Google Scholar]
- IARC. Some Industrial Chemicals and Dyestuffs; International Agency for Research on Cancer: Lyon, France, 1982. [Google Scholar]
- Sakabe, H.; Matsushita, H.; Koshi, S. Cancer among benzoyl chloride manufacturing workers. Ann. N. Y. Acad. Sci. 1976, 271, 67–70. [Google Scholar] [CrossRef]
- Peters, K.E.; Walters, C.C.; Moldowan, J.M. Biomarkers and isotopes in petroleum systems and Earth history. In The Biomarker Guide; Cambridge University Press: Cambridge, UK, 2004; Volume 2. [Google Scholar] [CrossRef]
- Neidle, E.L.; Hartnett, C.; Bonitz, S.; Ornston, L.N. DNA sequence of the Acinetobacter calcoaceticus catechol 1,2-dioxygenase I structural gene catA: Evidence for evolutionary divergence of intradiol dioxygenases by acquisition of DNA sequence repetitions. J. Bacteriol. 1988, 170, 4874–4880. [Google Scholar] [CrossRef] [Green Version]
- Hamzah, R.Y.; Al-Baharna, B.S. Catechol ring-cleavage in Pseudomonas cepacia: The simultaneous induction of ortho and meta pathways. Appl. Microbiol. Biotechnol. 1994, 41, 250–256. [Google Scholar] [CrossRef]
- Bhatt, P.; Pathak, V.M.; Joshi, S.; Bisht, T.S.; Singh, K.; Chandra, D. Chapter 12—Major metabolites after degradation of xenobiotics and enzymes involved in these pathways. In Smart Bioremediation Technologies; Bhatt, P., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 205–215. [Google Scholar] [CrossRef]
- Das, N.; Chandran, P. Microbial degradation of petroleum hydrocarbon contaminants: An overview. Biotechnol. Res. Int. 2011, 2011, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gemmell, R.T.; Knowles, C.J. Utilisation of aliphatic compounds by acidophilic heterotrophic bacteria. The potential for bioremediation of acidic wastewaters contaminated with toxic organic compounds and heavy metals. FEMS Microbiol. Lett. 2000, 192, 185–190. [Google Scholar] [CrossRef]
- Bhatt, P.; Gangola, S.; Bhandari, G.; Zhang, W.; Maithani, D.; Mishra, S.; Chen, S. New insights into the degradation of synthetic pollutants in contaminated environments. Chemosphere 2020. [Google Scholar] [CrossRef] [PubMed]
- Robledo-Ortíz, J.R.; Ramírez-Arreola, D.E.; Pérez-Fonseca, A.A.; Gómez, C.; González-Reynoso, O.; Ramos-Quirarte, J.; González-Núñez, R. Benzene, toluene, and o-xylene degradation by free and immobilized P. putida F1 of postconsumer agave-fiber/polymer foamed composites. Int. Biodeteriorat. Biodegradat. 2011, 65, 539–546. [Google Scholar] [CrossRef]
- Di Martino, C.; López, N.I.; Raiger Iustman, L.J. Isolation and characterization of benzene, toluene and xylene degrading Pseudomonas sp. selected as candidates for bioremediation. Int. Biodeteriorat. Biodegradat. 2012, 67, 15–20. [Google Scholar] [CrossRef]
- De Lima-Morales, D.; Chaves-Moreno, D.; Wos-Oxley, M.L.; Jáuregui, R.; Vilchez-Vargas, R.; Pieper, D.H. Degradation of benzene by Pseudomonas veronii 1YdBTEX2 and 1YB2 is catalyzed by enzymes encoded in distinct catabolism gene clusters. Appl. Environ. Microbiol. 2015, 82, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Atashgahi, S.; Hornung, B.; Van Der Waals, M.J.; Da Rocha, U.N.; Hugenholtz, F.; Nijsse, B.; Molenaar, D.; Van Spanning, R.; Stams, A.J.M.; Gerritse, J.; et al. A benzene-degrading nitrate-reducing microbial consortium displays aerobic and anaerobic benzene degradation pathways. Sci. Rep. 2018, 8, 4490. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Lee, Y.; Jeon, C.O. Biodegradation of naphthalene, BTEX, and aliphatic hydrocarbons by Paraburkholderia aromaticivorans BN5 isolated from petroleum-contaminated soil. Sci. Rep. 2019, 9, 860. [Google Scholar] [CrossRef]
- Lohi, A.; Cuenca, M.A.; Anania, G.; Upreti, S.R.; Wan, L. Biodegradation of diesel fuel-contaminated wastewater using a three-phase fluidized bed reactor. J. Hazard. Mater. 2008, 154, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Rahul Mathur, A.K.; Balomajumder, C. Biological treatment and modeling aspect of BTEX abatement process in a biofilter. Bioresour. Technol. 2013, 142, 9–17. [Google Scholar] [CrossRef]
- El-Naas, M.H.; Acio, J.A.; El Telib, A.E. Aerobic biodegradation of BTEX: Progresses and prospects. J. Environ. Chem. Eng. 2014, 2, 1104–1122. [Google Scholar] [CrossRef]
- Eze, M.O.; Lütgert, S.A.; Neubauer, H.; Balouri, A.; Kraft, A.A.; Sieven, A.; Daniel, R.; Wemheuer, B. Metagenome assembly and metagenome-assembled genome sequences from a historical oil field located in Wietze, Germany. Microbiol. Resour. Announc. 2020, 9. [Google Scholar] [CrossRef]
- Liang, C.; Huang, Y.; Wang, H. pahE, a functional marker gene for polycyclic aromatic hydrocarbon-degrading bacteria. Appl. Environ. Microbiol. 2018, 85. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.-L.; Jiangyang, J.-H.; Nie, Y.; Wu, X.-L. Regulation of the alkane hydroxylase CYP153 gene in a gram-positive alkane-degrading bacterium, dietzia sp. strain DQ12-45-1b. Appl. Environ. Microbiol. 2016, 82, 608–619. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Sanz, D.; Redondo-Nieto, M.; Guirado, M.; Jiménez, O.P.; Millán, R.; Martín, M.; Rivilla, R. Metagenomic insights into the bacterial functions of a diesel-degrading consortium for the rhizoremediation of diesel-polluted soil. Genes 2019, 10, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, J.; Gerali, F.; Macaulay, F.; Sorkhabi, R. The history of the European oil and gas industry (1600s–2000s). Geol. Soc. Lond. Spec. Publ. 2018, 465, 1–24. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2012, 41, e1. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Wemheuer, F.; Berkelmann, D.; Wemheuer, B.; Daniel, R.; Vidal, S.; Daghela, H.B.B. Agroforestry management systems drive the composition, diversity, and function of fungal and bacterial endophyte communities in Theobroma cacao leaves. Microorganisms 2020, 8, 405. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Project, G.; et al. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyatt, D.; Chen, G.-L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Eze, M.O.; George, S.C.; Hose, G.C. Dose-response analysis of diesel fuel phytotoxicity on selected plant species. Chemosphere 2021, 263, 128382. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org (accessed on 24 May 2019).
- Van der Vliet, L.; Ritz, C. Statistics for analyzing ecotoxicity test data. In Encyclopedia of Aquatic Ecotoxicology; Férard, J.-F., Blaise, C., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 1081–1096. [Google Scholar] [CrossRef]
- Lee, S.; Lee, D.-K. What is the proper way to apply the multiple comparison test? Korean J. Anesthesiol. 2018, 71, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Rivers, A.R.; Sharma, S.; Tringe, S.G.; Martin, J.; Joye, S.B.; Moran, M.A. Transcriptional response of bathypelagic marine bacterioplankton to the Deepwater Horizon oil spill. ISME J. 2013, 7, 2315–2329. [Google Scholar]
- Xu, X.; Liu, W.; Tian, S.; Wang, W.; Qi, Q.; Jiang, P.; Gao, X.; Li, F.; Li, H.; Yu, H. Petroleum hydrocarbon-degrading bacteria for the remediation of oil pollution under aerobic conditions: A perspective analysis. Front. Microbiol. 2018, 9, 2885. [Google Scholar]
- Labud, V.; Garcia, C.; Hernandez, T. Effect of hydrocarbon pollution on the microbial properties of a sandy and a clay soil. Chemosphere 2007, 66, 1863–1871. [Google Scholar] [CrossRef]
- Van Dorst, J.; Siciliano, S.D.; Winsley, T.; Snape, I.; Ferrari, B.C. Bacterial targets as potential indicators of diesel fuel toxicity in subantarctic soils. Appl. Environ. Microbiol. 2014, 80, 4021. [Google Scholar]
- Bouhajja, E.; McGuire, M.; Liles, M.R.; Bataille, G.; Agathos, S.N.; George, I.F. Identification of novel toluene monooxygenase genes in a hydrocarbon-polluted sediment using sequence- and function-based screening of metagenomic libraries. Appl. Microbiol. Biotechnol. 2016, 101, 797–808. [Google Scholar] [CrossRef]
- Iwaki, H.; Yamamoto, T.; Hasegawa, Y. Isolation of marine xylene-utilizing bacteria and characterization of Halioxenophilus aromaticivorans gen. nov., sp. nov. and its xylene degradation gene cluster. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef] [PubMed]
- Takeda, Y.; Takase, K.; Yamato, I.; Abe, K. Sequencing and characterization of the xyl operon of a gram-positive bacterium, tetragenococcus halophila. Appl. Environ. Microbiol. 1998, 64, 2513–2519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roling, W.; Ortega-Lucach, O.; Larter, S.; Head, I. Acidophilic microbial communities associated with a natural, biodegraded hydrocarbon seepage. J. Appl. Microbiol. 2006, 101, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Obieze, C.C.; Chikere, C.B.; Selvarajan, R.; Adeleke, R.; Ntushelo, K.; Akaranta, O. Functional attributes and response of bacterial communities to nature-based fertilization during hydrocarbon remediation. Int. Biodeterior. Biodegrad. 2020, 154, 105084. [Google Scholar] [CrossRef]
- Sipilä, T.P.; Keskinen, A.-K.; Åkerman, M.-L.; Fortelius, C.; Haahtela, K.; YrjãLã, K. High aromatic ring-cleavage diversity in birch rhizosphere: PAH treatment-specific changes of I.E.3 group extradiol dioxygenases and 16S rRNA bacterial communities in soil. ISME J. 2008, 2, 968–981. [Google Scholar] [CrossRef] [Green Version]
- Azubuike, C.C.; Chikere, C.B.; Okpokwasili, G.C. Bioremediation techniques–classification based on site of application: Principles, advantages, limitations and prospects. World J. Microbiol. Biotechnol. 2016, 32, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-X.; Hu, X.; Cao, Y.; Pang, W.-J.; Huang, J.-Y.; Guo, P.; Huang, L. Biodegradation of phenanthrene and heavy metal removal by acid-tolerant Burkholderia fungorum FM-2. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Okoh, A.; Ajisebutu, S.; Babalola, G.; Trejo-Hernandez, M.R. Potential of Burkholderia cepacia RQ1 in the biodegradation of heavy crude oil. Int. Microbiol. 2001, 4, 83–87. [Google Scholar] [CrossRef]
- Lee, Y.; Jeon, C.O. Paraburkholderia aromaticivorans sp. nov., an aromatic hydrocarbon-degrading bacterium, isolated from gasoline-contaminated soil. Int. J. Syst. Evol. Microbiol. 2018, 68, 1251–1257. [Google Scholar] [CrossRef]
- Dias, G.M.; Pires, A.D.S.; Grilo, V.S.; Castro, M.R.; Vilela, L.D.F.; Neves, B.C. Comparative genomics of Paraburkholderia kururiensis and its potential in bioremediation, biofertilization, and biocontrol of plant pathogens. Microbiology 2019, 8, e00801. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, R.D.; Savage, D.C.; Sayler, G.S.; Stacey, G. Biodegradation of aromatic hydrocarbons in an extremely acidic environment. Appl. Environ. Microbiol. 1998, 64, 4180–4184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovanella, P.; Vieira, G.A.L.; Ramos Otero, I.V.; Pais Pellizzer, E.; de Jesus Fontes, B.; Sette, L.D. Metal and organic pollutants bioremediation by extremophile microorganisms. J. Hazard. Mater. 2020, 382, 121024. [Google Scholar] [CrossRef] [PubMed]
- Arenghi, F.L.G.; Berlanda, D.; Galli, E.; Sello, G.; Barbieri, P. Organization and regulation of metaCleavage pathway genes for toluene and o-xylene derivative degradation in Pseudomonas stutzeri OX1. Appl. Environ. Microbiol. 2001, 67, 3304–3308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartilson, M.; Shingler, V. Nucleotide sequence and expression of the catechol 2,3-dioxygenase-encoding gene of phenolcatabolizing Pseudomonas CF600. Gene 1989, 85, 233–238. [Google Scholar] [CrossRef]
- Suenaga, H.; Koyama, Y.; Miyakoshi, M.; Miyazaki, R.; Yano, H.; Sota, M.; Ohtsubo, Y.; Tsuda, M.; Miyazaki, K. Novel organization of aromatic degradation pathway genes in a microbial community as revealed by metagenomic analysis. ISME J. 2009, 3, 1335–1348. [Google Scholar] [CrossRef]
- Garrido-Sanz, D.; Manzano, J.; Martín, M.; Redondo-Nieto, M.; Rivilla, R. Metagenomic analysis of a biphenyl-degrading soil bacterial consortium reveals the metabolic roles of specific populations. Front. Microbiol. 2018, 9, 232. [Google Scholar] [CrossRef]
- Sala-Trepat, J.M.; Evans, W.C. The meta Cleavage of Catechol by Azotobacter Species. 4-Oxalocrotonate Pathway. JBIC J. Biol. Inorg. Chem. 1971, 20, 400–413. [Google Scholar] [CrossRef]
- Li, J.; Luo, C.; Zhang, D.; Cai, X.; Jiang, L.; Zhang, G. Stable-Isotope Probing-Enabled Cultivation of the Indigenous Bacterium Ralstonia sp. Strain M1, Capable of Degrading Phenanthrene and Biphenyl in Industrial Wastewater. Appl. Environ. Microbiol. 2019, 85, e00511-19. [Google Scholar] [CrossRef] [Green Version]
- Cao, B.; Nagarajan, K.; Loh, K.-C. Biodegradation of aromatic compounds: Current status and opportunities for biomolecular approaches. Appl. Microbiol. Biotechnol. 2009, 85, 207–228. [Google Scholar] [CrossRef]
- Barriault, D.; Durand, J.; Maaroufi, H.; Eltis, L.D.; Sylvestre, M. Degradation of polychlorinated biphenyl metabolites by naphthalene-catabolizing enzymes. Appl. Environ. Microbiol. 1998, 64, 4637–4642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohrbacher, F.; St-Arnaud, M. Root exudation: The ecological driver of hydrocarbon rhizoremediation. Agronomy 2016, 6, 19. [Google Scholar] [CrossRef]
- He, C.; Li, Y.; Huang, C.; Chen, F.; Ma, Y. Genome sequence and metabolic analysis of a fluoranthene-degrading strain Pseudomonas aeruginosa DN1. Front. Microbiol. 2018, 9, 2595. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Linear Hypothesis (Tukey Contrasts) | Estimate | Std. Error | t Value | Pr(>|t|) |
|---|---|---|---|---|
| Control at T60–Control at T0 == 0 | −1.4133 | 0.0721 | −19.61 | 3.42e-06 *** |
| Inoculated at T60–Control at T0 == 0 | −3.2133 | 0.0721 | −44.59 | 2.56e-08 *** |
| Inoculated at T60–Control at T60 == 0 | −1.8000 | 0.0721 | −24.98 | 8.13e-07 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eze, M.O. Metagenome Analysis of a Hydrocarbon-Degrading Bacterial Consortium Reveals the Specific Roles of BTEX Biodegraders. Genes 2021, 12, 98. https://doi.org/10.3390/genes12010098
Eze MO. Metagenome Analysis of a Hydrocarbon-Degrading Bacterial Consortium Reveals the Specific Roles of BTEX Biodegraders. Genes. 2021; 12(1):98. https://doi.org/10.3390/genes12010098
Chicago/Turabian StyleEze, Michael O. 2021. "Metagenome Analysis of a Hydrocarbon-Degrading Bacterial Consortium Reveals the Specific Roles of BTEX Biodegraders" Genes 12, no. 1: 98. https://doi.org/10.3390/genes12010098
APA StyleEze, M. O. (2021). Metagenome Analysis of a Hydrocarbon-Degrading Bacterial Consortium Reveals the Specific Roles of BTEX Biodegraders. Genes, 12(1), 98. https://doi.org/10.3390/genes12010098