
Identification, Molecular Characterization, and Tissue Expression Profiles of Three Smad Genes from Water Buffalo (Bubalus bubalis)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, RNA Extraction, and cDNA Synthesis
2.2. Primer Design and Gene Isolation
2.3. Analysis of Gene Sequence and Structure
2.4. Physiochemical Characterization, Motif and Structure
2.5. RT-qPCR and Tissue Differential Expression
3. Results
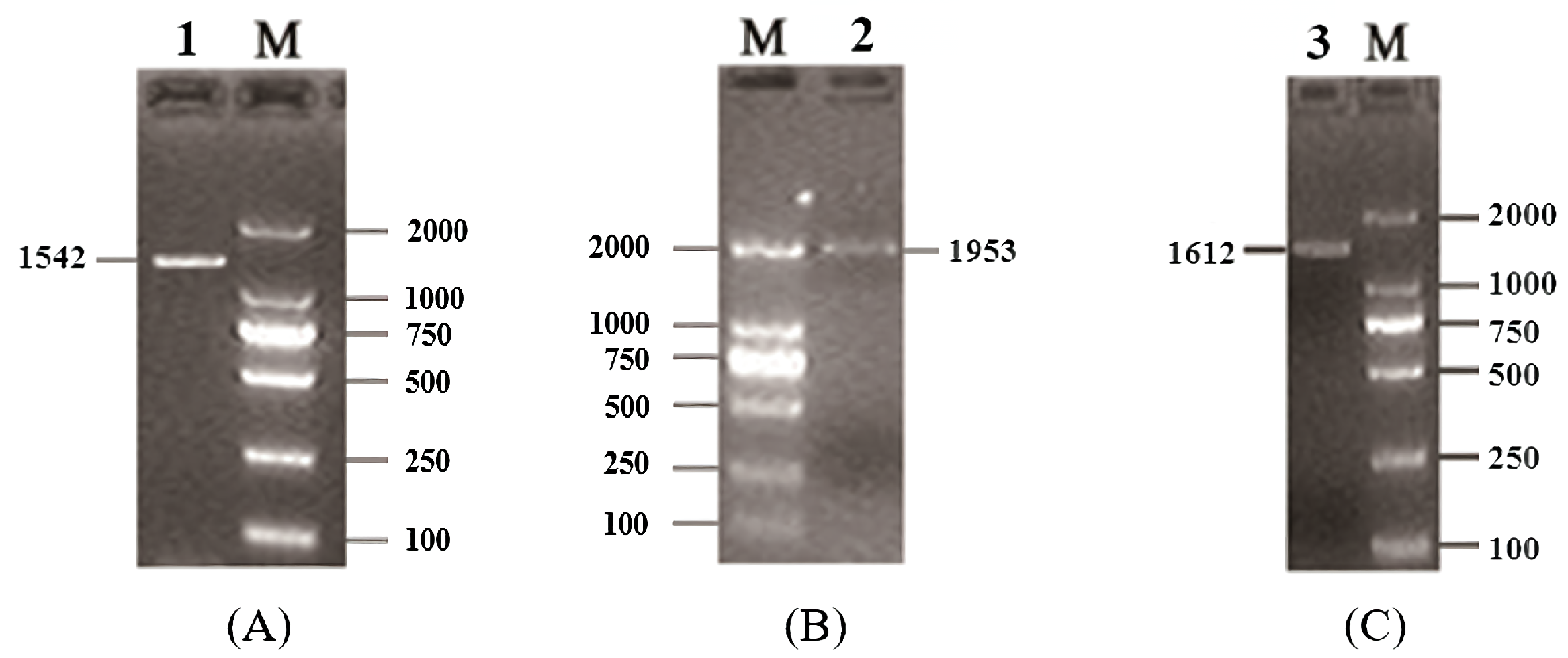
3.1. Isolation and Identification of Buffalo Smad1/4/5
3.2. Gene Sequence and Structure Analysis
3.3. Basic Physicochemical Properties of the Three Proteins
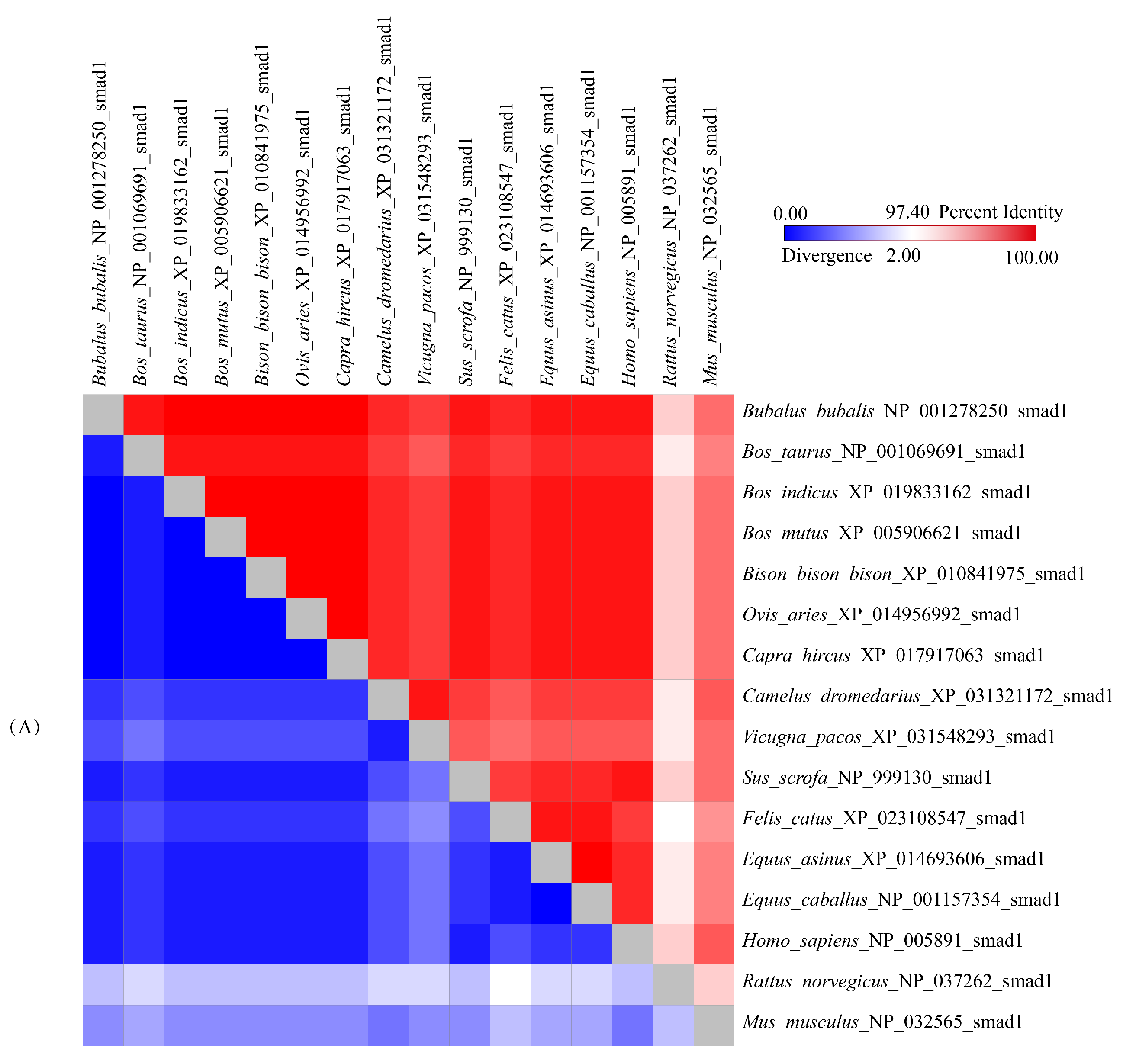
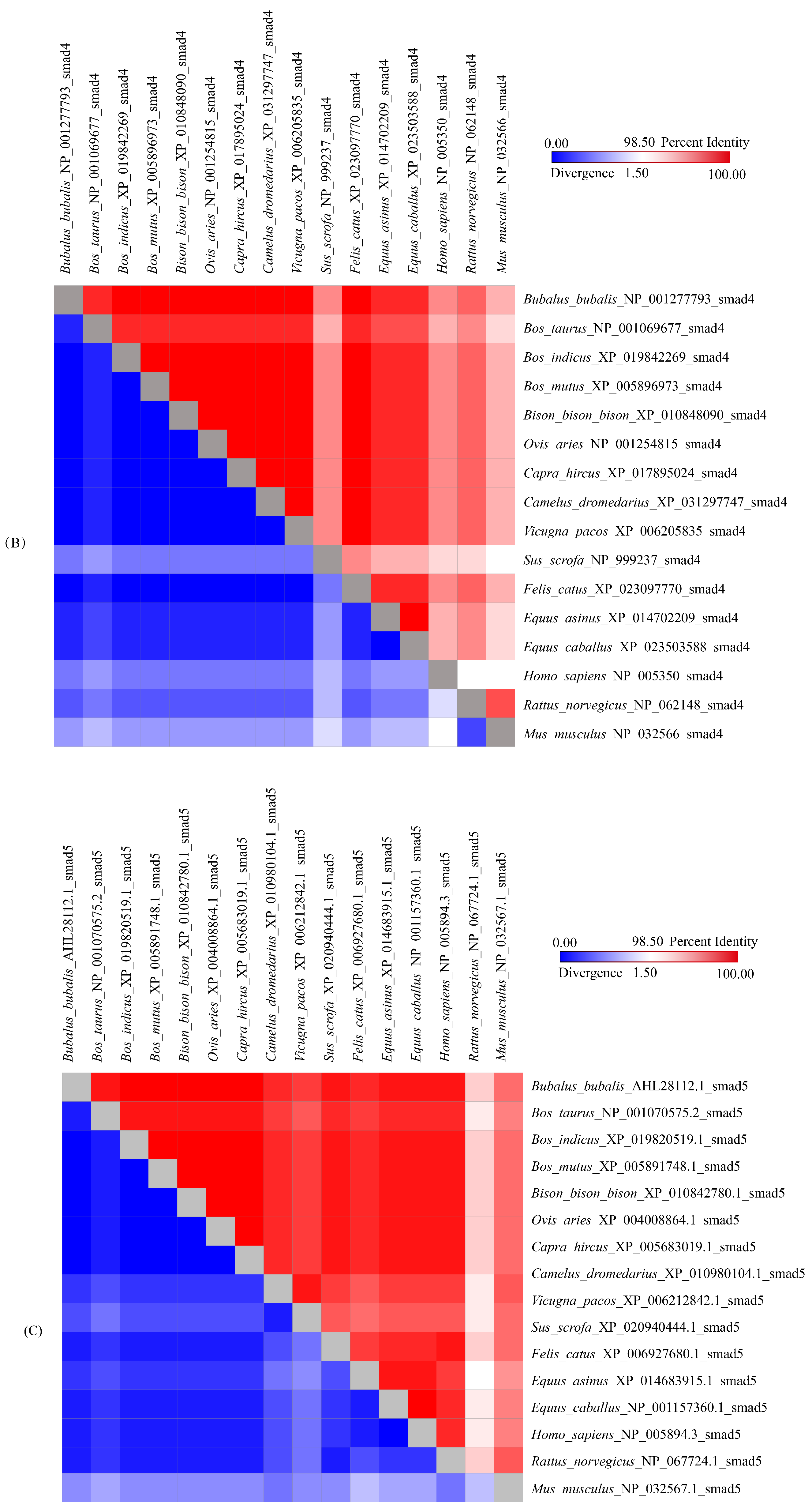
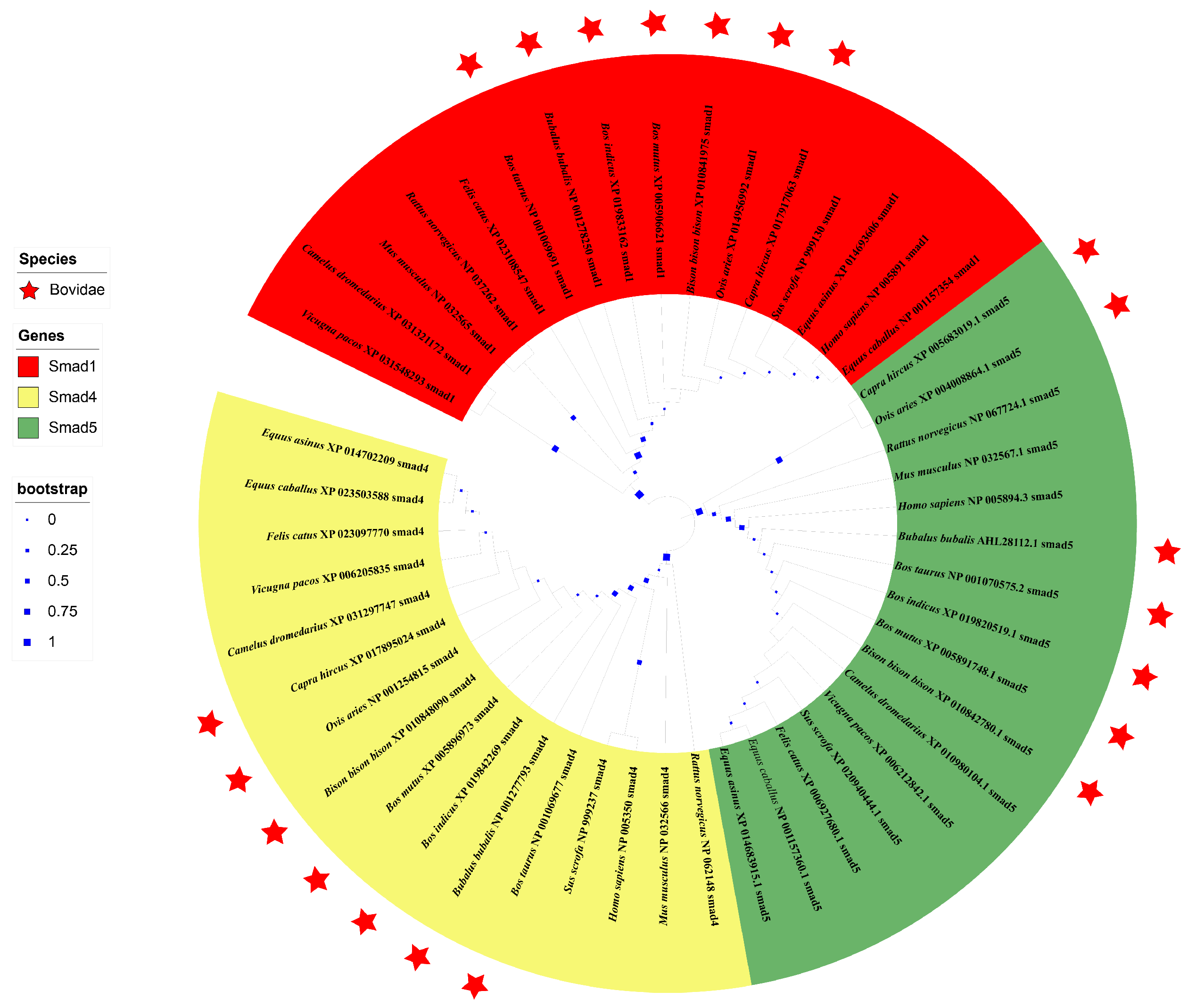
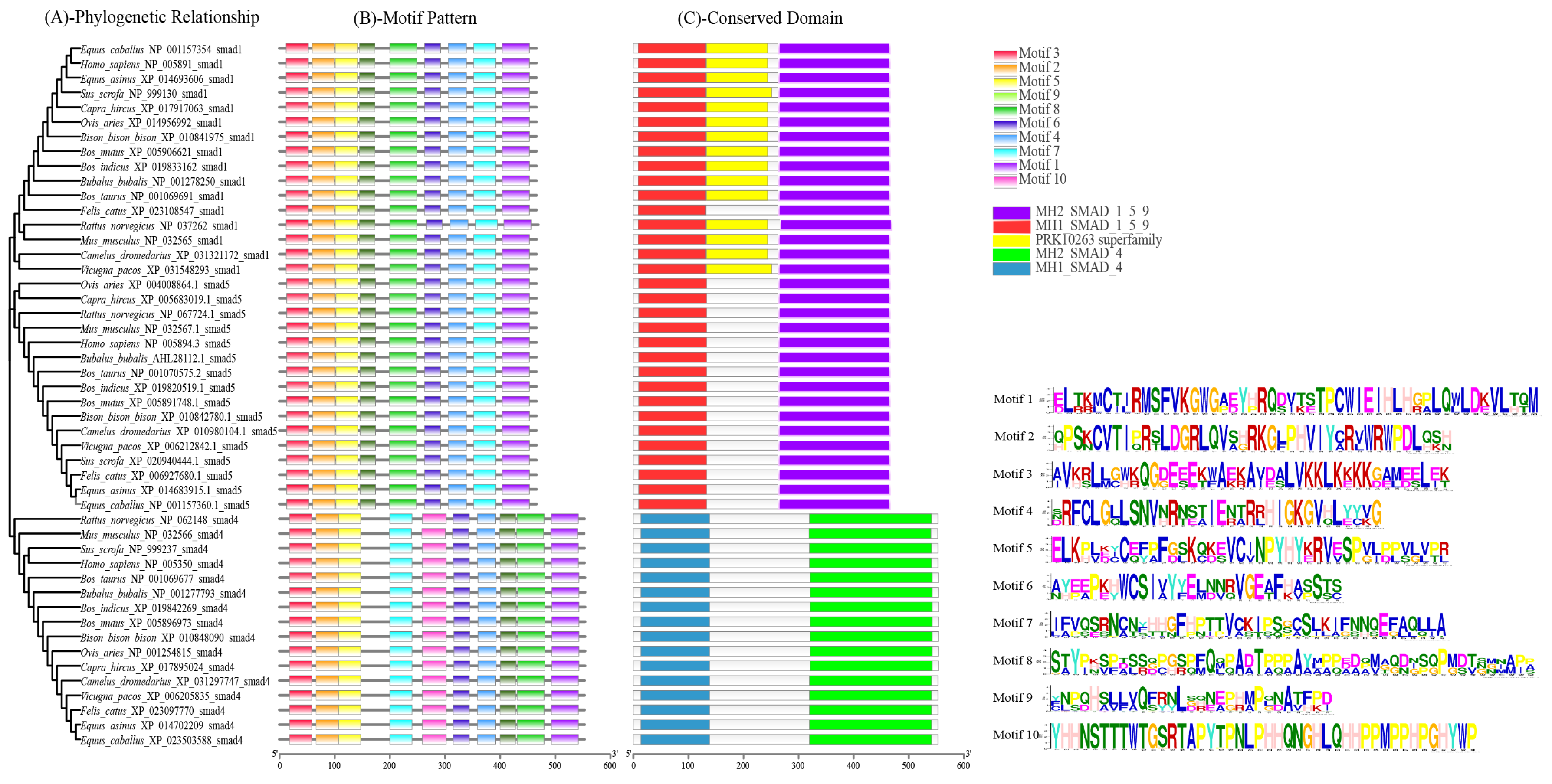
3.4. Sequence Identities and Phylogenetic Analysis
3.5. Structure of Buffalo Smads 1, 4, and 5
3.6. Subcellular Location and Molecular Function
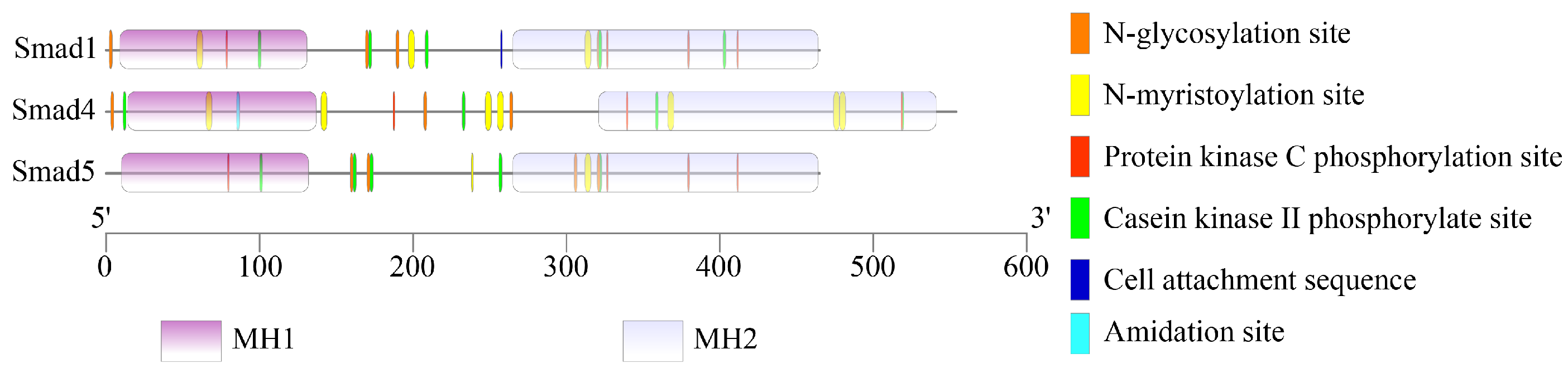
3.7. Amino Acid Modifications
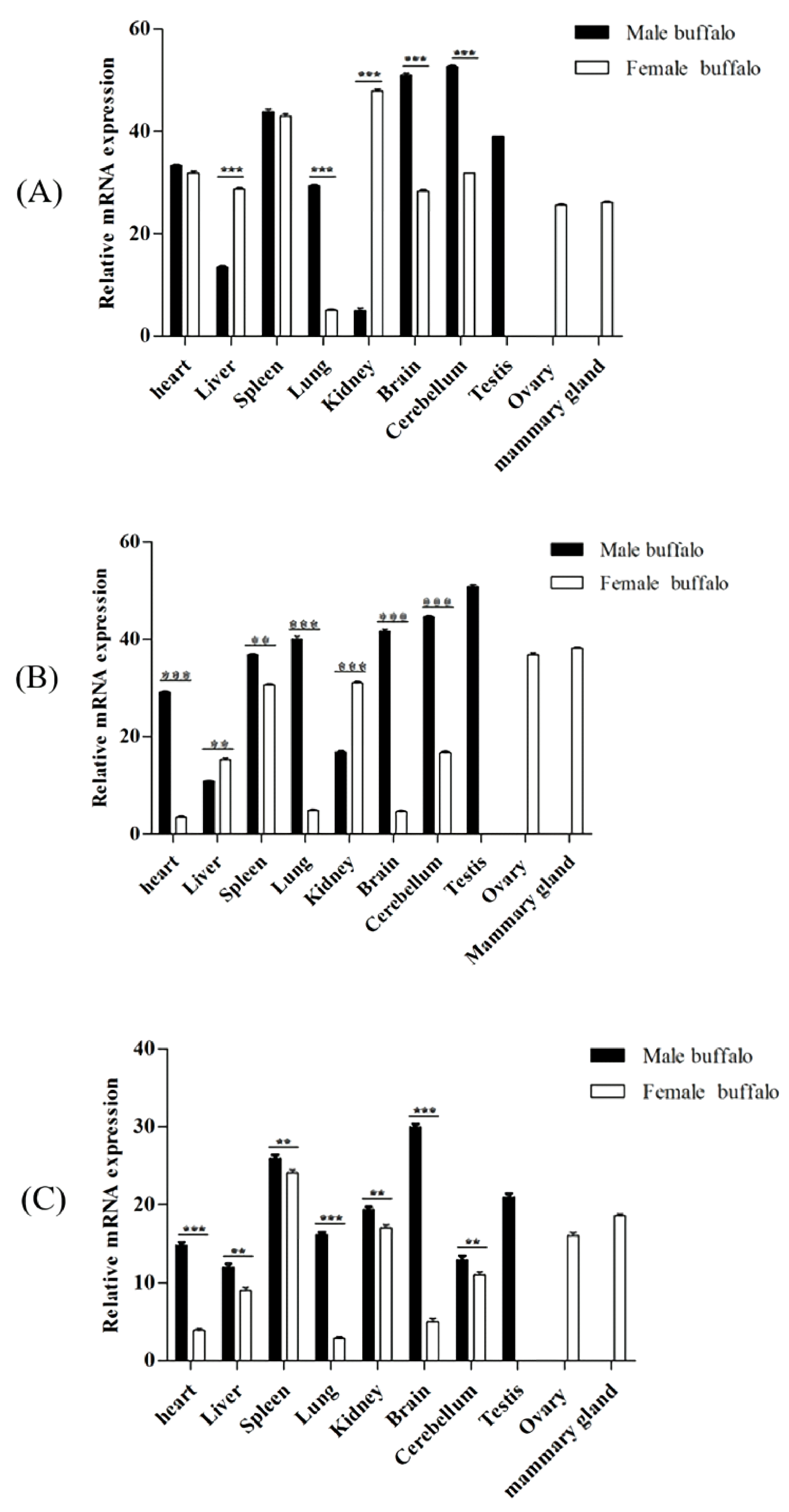
3.8. Tissue Expression Profile
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Massagué, J.; Seoane, J.; Wotton, D. Smad transcription factors. Genes Dev. 2005, 19, 2783–2810. [Google Scholar] [CrossRef] [Green Version]
- Mullen, A.C.; Wrana, J.L. TGF-β Family Signaling in Embryonic and Somatic Stem-Cell Renewal and Differentiation. Cold Spring Harb. Perspect. Biol. 2017, 9, a022186. [Google Scholar] [CrossRef]
- Hegarty, S.V.; O’Keeffe, G.W.; Sullivan, A.M. BMP-Smad 1/5/8 signalling in the development of the nervous system. Prog. Neurobiol. 2013, 109, 28–41. [Google Scholar] [CrossRef]
- Rahman, M.S.; Akhtar, N.; Jamil, H.M.; Banik, R.S.; Asaduzzaman, S.M. TGF-β/BMP signaling and other molecular events: Regulation of osteoblastogenesis and bone formation. Bone Res. 2015, 3, 15005. [Google Scholar] [CrossRef] [Green Version]
- Lochab, A.K.; Extavour, C.G. Bone Morphogenetic Protein (BMP) signaling in animal reproductive system development and function. Dev. Biol. 2017, 427, 258–269. [Google Scholar] [CrossRef]
- McNatty, K.P.; Fidler, A.E.; Juengel, J.L.; Quirke, L.D.; Smith, P.R.; Heath, D.A.; Lundy, T.; O’Connell, A.; Tisdall, D.J. Growth and paracrine factors regulating follicular formation and cellular function. Mol. Cell. Endocrinol. 2000, 163, 11–20. [Google Scholar] [CrossRef]
- Kumar, S.; Lakshmi Devi, H.; Singh Jalmeria, N.; Punetha, M.; Pandey, Y.; Samad, H.A.; Singh, G.; Sarkar, M.; Chouhan, V.S. Expression and functional role of bone morphogenetic proteins (BMPs) in placenta during different stages of pregnancy in water buffalo (Bubalus bubalis). Gen. Comp. Endocrinol. 2020, 285, 113249. [Google Scholar] [CrossRef]
- Lee, K.B.; Folger, J.K.; Rajput, S.K.; Smith, G.W. Temporal regulation of mRNAs for select bone morphogenetic proteins (BMP), BMP receptors and their associated SMAD proteins during bovine early embryonic development: Effects of exogenous BMP2 on embryo developmental progression. Reprod. Biol. Endocrinol. 2014, 12, 67. [Google Scholar] [CrossRef] [Green Version]
- García, E.V.; Hamdi, M.; Barrera, A.D.; Sánchez-Calabuig, M.J.; Gutiérrez-Adán, A.; Rizos, D. Bovine embryo-oviduct interaction in vitro reveals an early cross talk mediated by BMP signaling. Reproduction 2017, 153, 631–643. [Google Scholar] [CrossRef] [Green Version]
- Neira, J.A.; Tainturier, D.; Peña, M.A.; Martal, J. Effect of the association of IGF-I, IGF-II, bFGF, TGF-β1, GM-CSF, and LIF on the development of bovine embryos produced in vitro. Theriogenology 2010, 73, 595–604. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, S.; Gao, X.; Ren, H.; Chen, J. Cloning and bioinformatics analysis of cDNA encoding cattle Smad4 gene. Front. Agric. China 2008, 2, 493–497. [Google Scholar] [CrossRef]
- Zimin, A.V.; Delcher, A.L.; Florea, L.; Kelley, D.R.; Schatz, M.C.; Puiu, D.; Hanrahan, F.; Pertea, G.; Van Tassell, C.P.; Sonstegard, T.S.; et al. A whole-genome assembly of the domestic cow, Bos taurus. Genome Biol. 2009, 10, R42. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT. Food and Agriculture Organization of the United Nations Statistics Database. 2018. Available online: http://www.fao.org/faostat/en/ (accessed on 22 September 2021).
- Warriach, H.M.; McGill, D.M.; Bush, R.D.; Wynn, P.C.; Chohan, K.R. A Review of Recent Developments in Buffalo Reproduction. Asian-Australas. J. Anim. Sci. 2015, 28, 451–455. [Google Scholar] [CrossRef]
- Lalitha, S. Primer Premier 5. Biotech Softw. Internet Rep. 2000, 1, 270–272. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Petersen, T.N.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.M. The Proteomics Protocols Handbook; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Hasebe, M.; Entani, T.; Takayama, S.; Tomita, M.; Yanagawa, H. Design of peptide inhibitors for the importin alpha/beta nuclear import pathway by activity-based profiling. Chem. Biol. 2008, 15, 940–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosugi, S.; Hasebe, M.; Matsumura, N.; Takashima, H.; Miyamoto-Sato, E.; Tomita, M.; Yanagawa, H. Six classes of nuclear localization signals specific to different binding grooves of importin alpha. J. Biol. Chem. 2009, 284, 478–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosugi, S.; Hasebe, M.; Tomita, M.; Yanagawa, H. Systematic identification of cell cycle-dependent yeast nucleocytoplasmic shuttling proteins by prediction of composite motifs. Proc. Natl. Acad. Sci. USA 2009, 106, 10171–10176. [Google Scholar] [CrossRef] [Green Version]
- Chou, K.C.; Shen, H.B. A new method for predicting the subcellular localization of eukaryotic proteins with both single and multiple sites: Euk-mPLoc 2.0. PLoS ONE 2010, 5, e9931. [Google Scholar] [CrossRef]
- Hata, A.; Chen, Y.G. TGF-β Signaling from Receptors to Smads. Cold Spring Harb. Perspect. Biol. 2016, 8, a022061. [Google Scholar] [CrossRef]
- Hill, C.S. Transcriptional Control by the SMADs. Cold Spring Harb. Perspect. Biol. 2016, 8, a022079. [Google Scholar] [CrossRef] [Green Version]
- Carreira, A.C.; Alves, G.G.; Zambuzzi, W.F.; Sogayar, M.C.; Granjeiro, J.M. Bone morphogenetic proteins: Structure, biological function and therapeutic applications. Arch. Biochem. Biophys. 2014, 561, 64–73. [Google Scholar]
- Rajesh, G.; Mishra, S.R.; Paul, A.; Punetha, M.; Vidyalakshmi, G.M.; Narayanan, K.; Bag, S.; Bhure, S.K.; Singh Chouhan, V.; Maurya, V.P.; et al. Transcriptional and translational abundance of Bone morphogenetic protein (BMP) 2, 4, 6, 7 and their receptors BMPR1A, 1B and BMPR2 in buffalo ovarian follicle and the role of BMP4 and BMP7 on estrogen production and survival of cultured granulosa cells. Res. Vet. Sci. 2018, 118, 371–388. [Google Scholar] [CrossRef]
- Budi, E.H.; Duan, D.; Derynck, R. Transforming Growth Factor-β Receptors and Smads: Regulatory Complexity and Functional Versatility. Trends Cell Biol. 2017, 27, 658–672. [Google Scholar] [CrossRef]
- Xu, P.; Lin, X.; Feng, X.H. Posttranslational Regulation of Smads. Cold Spring Harb. Perspect. Biol. 2016, 8, a022087. [Google Scholar] [CrossRef] [Green Version]
- Koefoed, K.; Skat-Rørdam, J.; Andersen, P.; Warzecha, C.B.; Pye, M.; Andersen, T.A.; Ajbro, K.D.; Bendsen, E.; Narimatsu, M.; Vilhardt, F.; et al. The E3 ubiquitin ligase SMURF1 regulates cell-fate specification and outflow tract septation during mammalian heart development. Sci. Rep. 2018, 8, 9542. [Google Scholar] [CrossRef]
- Tong, K.K.; Ma, T.C.; Kwan, K.M. BMP/Smad signaling and embryonic cerebellum development: Stem cell specification and heterogeneity of anterior rhombic lip. Dev. Growth Differ. 2015, 57, 121–134. [Google Scholar] [CrossRef]
- Budna, J.; Rybska, M.; Ciesiółka, S.; Bryja, A.; Borys, S.; Kranc, W.; Wojtanowicz-Markiewicz, K.; Jeseta, M.; Sumelka, E.; Bukowska, D.; et al. Expression of genes associated with BMP signaling pathway in porcine oocytes before and after IVM—A microarray approach. Reprod. Biol. Endocrinol. RB&E 2017, 15. [Google Scholar] [CrossRef]
- Shimizu, T.; Magata, F.; Abe, Y.; Miyamoto, A. Bone morphogenetic protein 4 (BMP-4) and BMP-7 induce vascular endothelial growth factor expression in bovine granulosa cells. Anim. Sci. J. 2012, 83, 663–667. [Google Scholar] [CrossRef]
- Rajesh, G.; Paul, A.; Mishra, S.R.; Bharati, J.; Thakur, N.; Mondal, T.; Soren, S.; Harikumar, S.; Narayanan, K.; Chouhan, V.S.; et al. Expression and functional role of Bone Morphogenetic Proteins (BMPs) in cyclical corpus luteum in buffalo (Bubalus bubalis). Gen. Comp. Endocrinol. 2017, 240, 198–213. [Google Scholar] [CrossRef]
- Xu, Y.; Li, E.; Han, Y.; Chen, L.; Xie, Z. Differential expression of mRNAs encoding BMP/Smad pathway molecules in antral follicles of high- and low-fecundity Hu sheep. Anim. Reprod. Sci. 2010, 120, 47–55. [Google Scholar] [CrossRef]
- Steuernagel, L.; Meckbach, C.; Heinrich, F.; Zeidler, S.; Schmitt, A.O.; Gültas, M. Computational identification of tissue-specific transcription factor cooperation in ten cattle tissues. PLoS ONE 2019, 14, e0216475. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Pang, Y.; Moses, H.L. TGF-β and immune cells: An important regulatory axis in the tumor microenvironment and progression. Trends Immunol. 2010, 31, 220–227. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers(5′–3′) | Amplicon Length (bp) | Annealing Temperature (°C) | Purpose |
|---|---|---|---|---|
| Smad1 | F: AAGCCTCTTTCACTGTCCTTTCAC | 1542 | 60 | CDS isolation |
| R: GTCTGACTCGTCCATCCTTCAAGT | ||||
| Smad4 | F: GCTGCCTCCGAAAGATCAAAAT | 1953 | 55 | CDS isolation |
| R: GCTCTGAGCCATGCCTGAAAAGT | ||||
| Smad5 | F: TGGCACTTATGAAGATAAAGGTCA | 1612 | 55 | CDS isolation |
| R: CTGTACTGGAAGTTTCCCTAAAAT | ||||
| Smad1 | F: CCCTCACTCTCCGACCAG | 238 | 57.1 | Differential expression |
| R: ACCATCCACCAACACGCT | ||||
| Smad4 | F: TCATTATCCACTGAAGGGCATT | 173 | 54 | Differential expression |
| R: ATACTGGCGGGCTGACTTGT | ||||
| Smad5 | F: TTTCCCTTATCTCCAAAT | 286 | 51.9 | Differential expression |
| R: CGACAGGCTGAACATCTC | ||||
| ACTB | F: TGGGCATGGAATCCTG | 196 | 57.9 | internal reference |
| R: GGCGCGATGATCTTGAT | ||||
| RPS23 | R: CAGGAAGTGTGTCAGGGT | 154 | 50.8 | internal reference |
| F: TCCAGGAATGTCACCAAC | ||||
| RPS15 | R: GATGGTGGGCAGCATGGTT | 159 | 58.6 | internal reference |
| F: AAGCGGGAGGAATGGGTGG |
| Items | Smad1 | Smad4 | Smad5 | |||
|---|---|---|---|---|---|---|
| Buffalo | Cattle | Buffalo | Cattle | Buffalo | Cattle | |
| Isoelectric point (PI) | 6.9 | 6.9 | 6.5 | 6.5 | 7.63 | 7.63 |
| Molecular weight (MW) | 52.25 kDa | 52.22 kDa | 60.57 kDa | 60.54 kDa | 52.33 kDa | 52.25 kDa |
| Formula | ||||||
| Strongly acidic amino acids (E, D) | 44 | 44 | 51 | 51 | 42 | 42 |
| Strongly basic amino acids (R, K) | 42 | 42 | 44 | 44 | 43 | 43 |
| Polar amino acids (S, T, Y, N, C, Q) | 148 | 147 | 160 | 159 | 145 | 145 |
| Hydrophobic amino acids (F, W, V, A, I, L) | 126 | 126 | 173 | 173 | 132 | 132 |
| Instability index (II) | 60.84 | 60.24 | 50.95 | 50.83 | 64.01 | 64.42 |
| Grand average of hydropathicity (GRAVY) | −0.568 | −0.567 | −0.389 | −0.388 | −0.517 | −0.516 |
| Aliphatic index (AI) | 65.35 | 65.35 | 76.06 | 76.06 | 68.09 | 68.09 |
| Motif1 | Motif2 | Motif3 | Motif4 | Motif5 | Motif6 | Motif7 | Motif8 | Motif9 | Motif10 | |
|---|---|---|---|---|---|---|---|---|---|---|
| E-value | 3.0 × 10−2454 | 3.6 × 10−1957 | 9.4 ×10−1793 | 1.2 × 10−1544 | 2.1 × 10−1704 | 1.3 × 10−1271 | 1.5 × 10−1374 | 1.5 × 10−1722 | 1.1 × 10−996 | 9.1 × 10−638 |
| Sites | 48 | 48 | 48 | 48 | 48 | 48 | 45 | 48 | 48 | 16 |
| Width | 50 | 41 | 41 | 34 | 41 | 30 | 41 | 50 | 29 | 44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Zhang, G.; Miao, Y. Identification, Molecular Characterization, and Tissue Expression Profiles of Three Smad Genes from Water Buffalo (Bubalus bubalis). Genes 2021, 12, 1536. https://doi.org/10.3390/genes12101536
Zhang J, Zhang G, Miao Y. Identification, Molecular Characterization, and Tissue Expression Profiles of Three Smad Genes from Water Buffalo (Bubalus bubalis). Genes. 2021; 12(10):1536. https://doi.org/10.3390/genes12101536
Chicago/Turabian StyleZhang, Jie, Guangle Zhang, and Yongwang Miao. 2021. "Identification, Molecular Characterization, and Tissue Expression Profiles of Three Smad Genes from Water Buffalo (Bubalus bubalis)" Genes 12, no. 10: 1536. https://doi.org/10.3390/genes12101536
APA StyleZhang, J., Zhang, G., & Miao, Y. (2021). Identification, Molecular Characterization, and Tissue Expression Profiles of Three Smad Genes from Water Buffalo (Bubalus bubalis). Genes, 12(10), 1536. https://doi.org/10.3390/genes12101536