1. Introduction
India is the fourth largest producer of oilseeds just after the USA, China, and Brazil, accounting for about 19% of the global area and 2.7% of global production [
1]. Presently, India needs 25 million tons (MT) of vegetable oils, of which merely 10.5 MT is produced domestically. Owing to this, India is also the world’s largest consumer and importer of vegetable oils and meets up to 60% of its domestic demand through imports, costing it up to USD 10 billion annually [
2,
3]. Moreover, the domestic edible oils demands will increase in the coming years since per capita consumption of vegetable oil is on a steady rise in India due to its ever increasing population. The low productivity of oilseed crops is the major reason behind such a huge imbalance between demand and supply of edible oils in India [
4]. Among the seven edible oilseed crops grown in the country, brassica oilseeds alone contribute more than 30% to total oil production. Indian mustard [
Brassica juncea (L.) Czern & Coss.] an allotetraploid (2n = 4x = 36, AABB), is the most predominantly cultivated crop occupying approximately 90% of the total area under brassica oilseeds cultivation in India. The average yields of brassica oilseeds are 1245 kg/hectare in India versus global productivity of 1994 kg/hectare [
5,
6]. Such a large instability in yield and production of this crop is mainly due to its sensitivity against various abiotic and biotic extremities which is anticipated to rise in the near future due to changing climatic conditions [
7,
8]. Among various stresses, fungal diseases viz., white rust, alternaria blight, downy mildew, and Sclerotinia stem rot are the major factors influencing crop productivity in Indian mustard. Among them, Sclerotinia stem rot has switched from being of minor significance to major significance since the last decade due to changes in climatic conditions and presently, one of the most devastating diseases of mustard at the global level causing up to 5–100% yield losses [
8,
9,
10,
11,
12]. This disease is caused by ubiquitous, cosmopolitan soil-borne hemibiotrophic fungus,
Sclerotinia sclerotiorum (Lib) de Bary, causing annual yield losses worth over several hundred million dollars [
13].
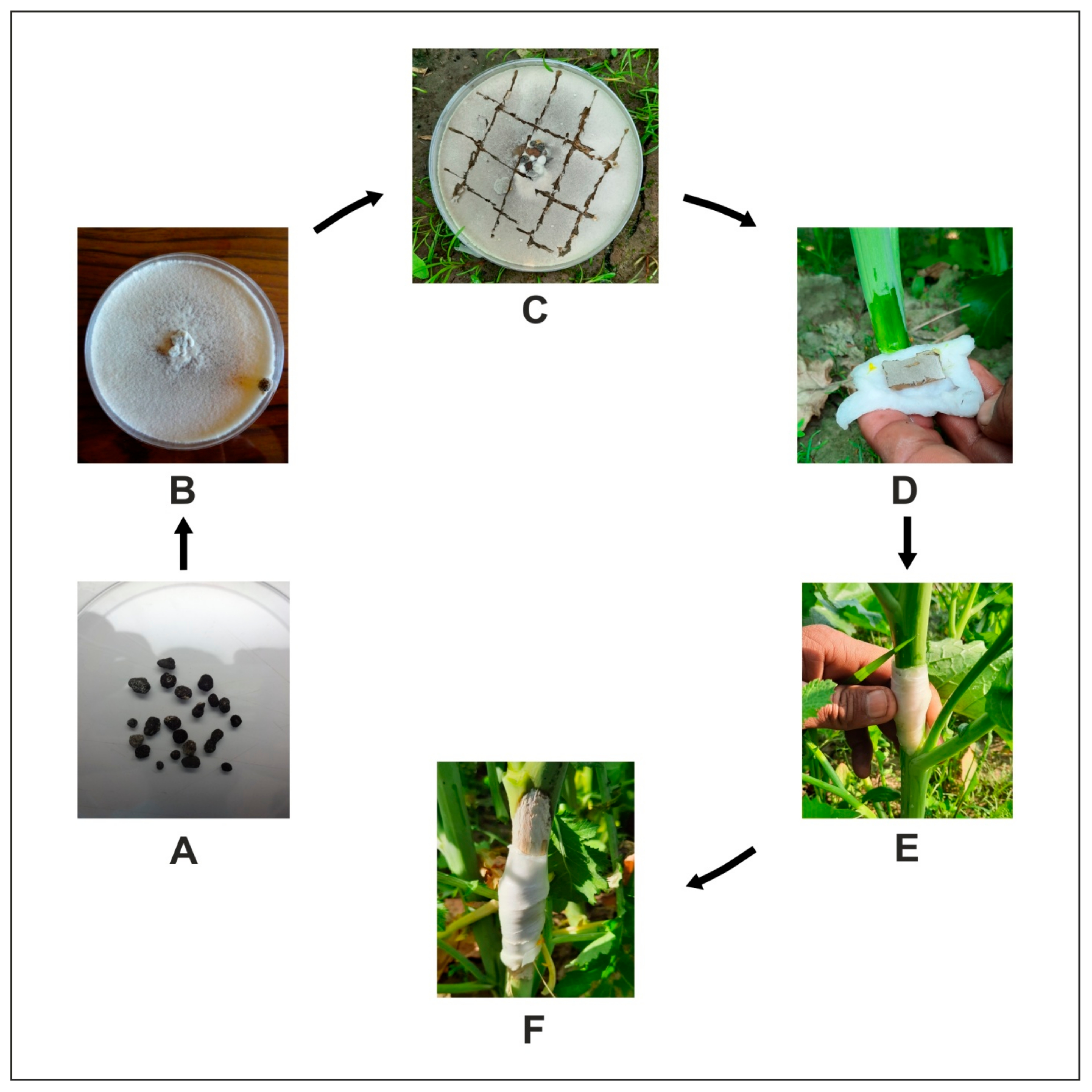
S. sclerotiorum exhibits dual infection mode in its host as its resting bodies (sclerotia) and can germinate either myceliogenically (soil-borne infection) to cause disease in the basal stem or can germinate carpogenically (air-borne infection) to cause disease in leaves and siliquae (
Figure 1).
Besides affecting almost all plant parts, the stem is the most affected host tissue and infection to stem is directly related to its girdling and plant lodging which is one of the most ultimate reasons for major yield losses in mustard at the field level [
12,
14,
15].
Changing climatic conditions and modern agricultural practices increase the risk of Sclerotinia rot epidemic development by allowing the pathogen to accumulate a high inoculum load [
16]. The control of this pathogen through cultural and chemical control is often very tedious and not that effective because of its complex mode of infection and its longer survival ability (up to 10 years in soil without host availability) in the form of a resting structure called sclerotia [
17]. In addition, fungicide application poses a serious threat to climate and adds further cost to crop cultivation. Therefore, host genetic resistance is the most convenient, economic, and eco-friendly approach for the effective control of this devastating pathogen [
18,
19]. Earlier attempts to identify resistant sources against this disease in Indian mustard were hampered as all the
B. juncea genotypes evaluated were found susceptible to Sclerotinia stem rot and any of the resistant sources reported belonged to other cruciferous crops and its wild relatives such as
Brassica napus,
B. fruticulosa,
B. rupestris,
B. incana,
B. insularis,
B. villosa,
Erucastrum cardaminoides,
E. abyssinicum,
Sinapis alba, and
Diplotaxis tenuisiliqua [
20,
21,
22,
23,
24,
25,
26,
27,
28,
29,
30,
31,
32], with no reports available about its resistance in
B. juncea, which is an important oil yielding crop in the Indian context. However, in recent few years, increasing attention has been paid which has ultimately led to the identification of a few Indian mustard genotypes resistant against this pathogen [
3,
5,
33,
34].
Successful infection of
S. sclerotiorum on mustard stems leads to the development of typical symptoms in the form of white greyish, water-soaked lesions which often extend as the disease progresses and cause stem girdling which ultimately leads to lodging and wilting of plants [
5]. Measurement of lesion length at a particular time after infection is generally used to assess the damage caused by this pathogen at the individual plant/stem level. This is because lesion length expansion has a direct positive relationship with disease severity, damage in the form of stem breakage, and plant collapse at the field level as well as the amount of secondary inoculum produced [
15]. Therefore, measurement of stem lesion length at a particular time after infection is a very useful component for the assessment of quantitative resistance against this pathogen in Indian mustard.
The knowledge about the pattern of inheritance and the nature of gene action involved in resistance is crucial for the effective utilization of the resistant source in disease resistance breeding programs. Information regarding the genetic basis of resistance allows breeders to frame an efficient breeding strategy for the development of resistant cultivars. The generation mean analysis (GMA) is a simple but effective approach for the estimation of the nature and magnitude of gene actions (additive or dominance) involved in a particular trait. Besides this, GMA also helps breeders in the detection of various types of epistasis, viz., additive × additive, additive × dominance and dominance × dominance operating in the inheritance of a particular trait [
35,
36,
37,
38,
39].
Recent advancement in plant biotechnology offers several techniques to assist crop geneticists and breeders in developing crop cultivars more efficiently. Proteomics is comparatively new tool among the different omic approaches frequently used by plant scientists. The proteome, the translational version of the genome, is a crucial functional player for mediating specific cellular processes, offering several advantages over other omics techniques. Post-translational modification reflects the functional impression of proteins at the cellular level. The information derived from proteomic studies can help plant breeders to modify plant genetic architecture and enable crop cultivars with high yield potential to improve crop qualities and various stress tolerances. The recent advancement in high-throughput analysis of crop proteins using LC-MS/MS helps to identify particular proteins. Moreover, it offers a new alternative for the determination of genes that are responsible for a particular trait. It might assist plant breeders in developing disease-resistant cultivars for sustainable agriculture [
40,
41,
42,
43]. Several recent studies indicate that the proteomic approach helps in understanding the molecular mechanism involved in plant–pathogen interaction, identification of the host’s resistance/susceptible factor(s), and pathogen’s virulence factor(s) at the molecular level [
43,
44,
45,
46].
In this context, the present investigation was designed with the aim to study the inheritance and protein expression pattern involved in Sclerotinia stem rot resistance in Indian mustard.
4. Discussion
Information regarding the pattern of inheritance as well as the nature of gene action involved in resistance against
Sclerotinia sclerotiorum helps crop geneticists and breeders to elect appropriate selection methods for breeding resistant cultivars. Previous studies have revealed that inheritance of Sclerotinia stem rot resistance varies from crop to crop being monogenic in
Vicia faba [
55] while polygenic in
B. napus [
23,
32]. Our study is perhaps the first to report on the genetic investigation and protein expression patterns of Sclerotinia stem rot resistance in
B. juncea. The meteorological data given in
Supplementary Table S1 revealed that environmental conditions at the experimental site were quite favorable for pathogen proliferation and disease development [
56]. The significance of generations for lesion length development observed in this study reveals that Sclerotinia stem rot resistance is a heritable trait. The increase in mean lesion length of F
1, F
2, BC
1P
1, and BC
1P
2 progenies over resistant parents indicates that genes contributing to resistance were recessive. The partial dominance of susceptibility over resistance was again confirmed when the mean lesion length of the F
1 generation was compared with their respective mid-parent values. A similar result was also reported by Baswana et al. [
57] in cauliflower, while overdominance of susceptibility in
Brassica napus was delineated by Khan et al. [
58,
59]. Similar to Khan et al. [
59] and Zhao et al. [
60], we also observed transgressive segregants for resistance in F
2 and BC
1P
1 generations of both populations. Such transgressive segregants were nearly asymptomatic without any lesion development which may be fixed later in selfing generations and can be utilized as a source of resistance in the future.
The significant individual and joint scaling test for mean lesion length in both populations indicate that resistance/susceptibility did not follow the simple Mendelian pattern of inheritance. This indicates the role of epistasis in the genetic control of resistance/susceptibility. Hence, the authors suggest six parameter models most appropriate to explain the inheritance pattern of resistance in the present study. Among six genetic components, our results signify the role of five genetic components in the genetic control of lesion length development, except additive × dominance (j) type of epistasis, which was non-significant in both populations. Among these genetic components, additive (d) gene effect and dominance × dominance (l) type of epistasis were of higher magnitude in the negative direction while dominance (h) gene effect and additive × additive type of epistasis was having higher magnitudes in the positive direction. According to Mather and Jinks [
61], the direction of gene effects controlling particular traits is determined by their associated signs. Therefore, in the present study, the additive (h) gene effect and the dominance × dominance (l) type of epistasis mainly governed resistance while the dominance (h) gene effect and the additive × additive (i) type of epistasis imparted susceptibility to
S. sclerotiorum in Indian mustard. In both populations, estimates of dominance-by-dominance effects (l) were significant and opposite in sign to those of dominance effects alone (h), indicating the role of duplicate epistasis in resistance expression. Similarly, Khan et al. [
59] also detected the role of dominance × dominance type of digenic interaction for cotyledon resistance in
Brassica napus against
S. sclerotiorum.
Both populations showed high broad and narrow-sense heritability for lesion length development indicating minimal environmental influence. Therefore, selection for resistance may be effective in Indian mustard because all genetic effects were above 80% according to the high broad-sense heritability for lesion length detected in the present study. Another explanation for high heritability during the present study is that the resistance/susceptibility might be controlled by a few major genes. The observed variation between narrow (h
2 ns) and broad sense (h
2 bs) heritability exhibits the involvement of dominance effect in the heredity of resistance. However, the value of h
2 ns was higher than h
2bs in C-I, which might be due to counteracting effects of additive and dominance genetic variance. Khan et al. [
58] observed moderate broad-sense heritability for leaf resistance against
S. sclerotiorum in oilseed rape. However, the estimates of the genetic parameters, viz., heritability and genetic advance together, are highly desirable for more accurately predicting the genetic gain under selection. In the present study, we found high heritability along with a moderate genetic advance for lesion length development. This indicates that effective progress towards resistance can be made through the selection of lower lesion length as selection efficiency depends upon the magnitudes of heritability and genetic advancement. As duplicate epistasis was prevalent, the selection of transgressive segregants for resistance is also possible in these populations. Similar to this, moderate to high narrow-sense heritability for resistance against
S. sclerotiorum has been reported by Baswana et al. [
57] in cauliflower and Castano et al. [
62] in sunflower.
The Wright estimates of the effective factors/minimum number of genes responsible for resistance/susceptibility ranged from 2.46 to 3.50 with an overall average of 2.98. This result strongly reveals the oligogenic nature of inheritance stating that Sclerotinia stem rot resistance might be controlled by at least three major effect genes. Although, inheritance patterns suggested that there may be few to many minor effect genes involved as well, along with major effect genes. Similar to the present study, Vleugels and Bockstaele [
63] also detected three major genes responsible for resistance against
S. trifoliorum in red clover while Moellers et al. [
64] reported both the main gene’s effect and epistatic loci responsible for resistance against
S. sclerotiorum in soybean. The estimates of the average degree of dominance and potency ratio lay between zero to one, which again revealed the partial dominance nature of genes responsible for lesion length development. The estimate of F value is an indicator of association between additive (D) and dominance (H) genetic variance over all loci controlling the trait under study. The negative F value in the present study indicated the presence of partial dominant genes in susceptible parents; therefore, susceptibility is partially dominant over resistance. The ratio of F/
was close to zero during the present investigation, which reveals that the magnitude and sign of the genes controlling the character are not equal and hence
is not a good estimator of dominance and only explains average dominance for resistance/susceptibility. The results of the present study suggested the role of both additive and non-additive genetic effects—although the non-additive part is slightly higher. Thus, a breeding approach that could exploit both additive and non-additive gene actions would be appropriate in the present situation. Hence, initial single seed descent till high homozygosity is achieved followed by reciprocal recurrent selection in succeeding generations seems to be the most appropriate breeding procedure for improvement and/or introgression of resistance into a desirable agronomic background. However, Barbetti et al. [
65] suggested that stem resistance in oilseed brassica against
S. sclerotiorum could be race/isolate specific(vertical resistance) or race non-specific (durable resistance), the latter being the most optimal and most effective resistance source for breeding novel cultivars with robust resistance across multiple pathotypes of
S. sclerotiorum. Although, we screened these populations only against a single isolate (
Hisar isolate) which is a prevalent virulent isolate/race of
S. sclerotiorum in CCS HAU, Hisar. However, the resistant parent (RH 1222-28) involved in the present study was found resistant/highly tolerant against multiple isolates of different locations as screened in the previous studies [
3,
5,
33,
34].
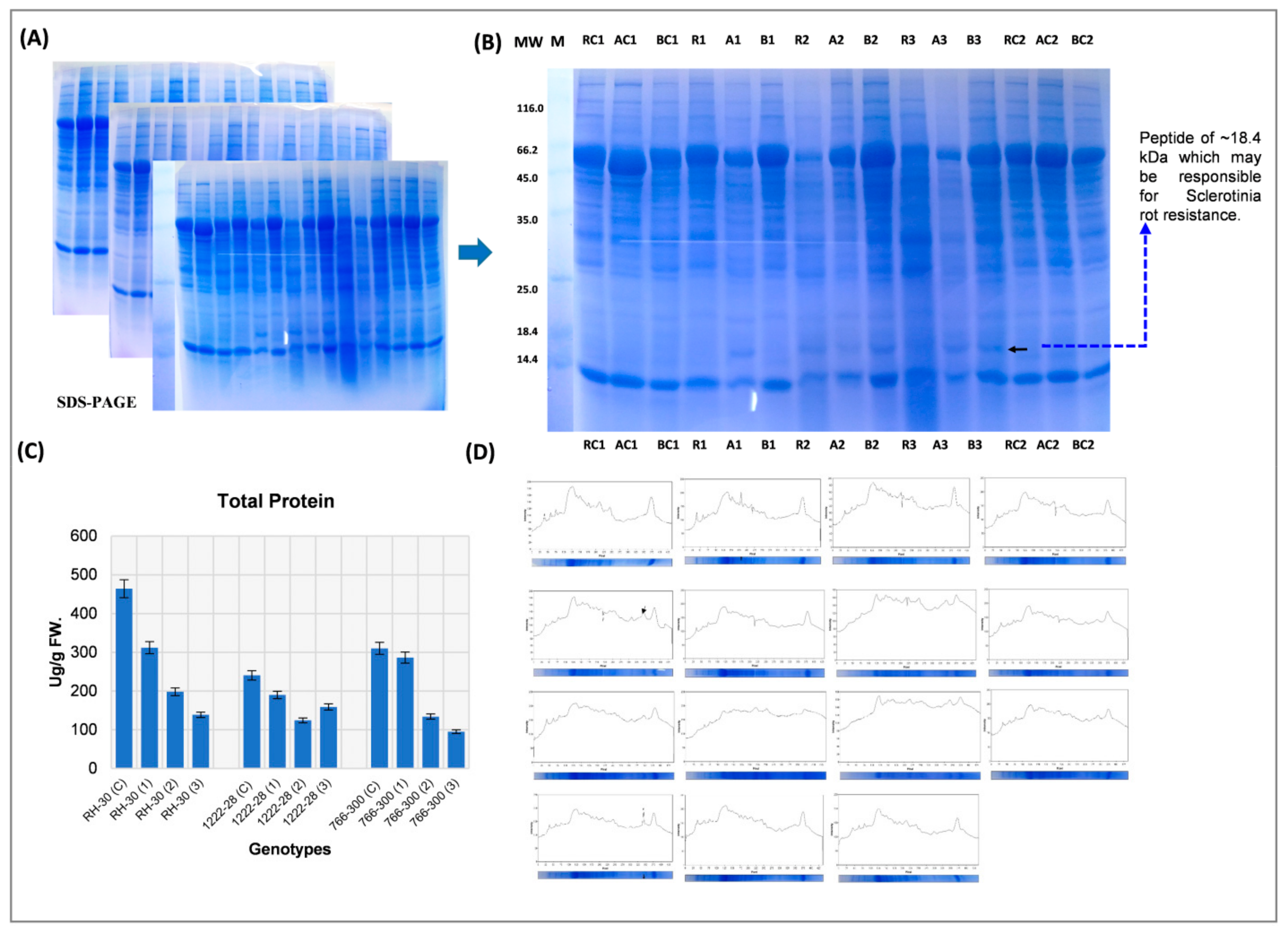
Moreover, the protein expression study revealed the involvement of peptidyl-prolyl-cis/trans-isomerases (PPIases or immunophilins) in resistance against
S. sclerotiorum. We observed a rapid accumulation of PPIase with a molecular weight of 18.4 kDa in the resistance genotype during early infection stage (6th day after infection). In contrast, in the susceptible genotypes, it appeared late (12th and 18th day after infection). The present study suggested that this PPIase has a definite role in resistance and implicates the corresponding protein as a biomarker for separating susceptible and resistant genotypes during the early stages of plant development. PPIases are the class of stress-responsive proteins of the immunophilin family. It catalyzes various biological functions such as transcription regulation, protein folding and degradation, signal transduction, and reactive oxygen species (ROS) regulation, cell wall strengthening to cope with stress conditions [
66,
67,
68,
69,
70]. A plethora of recent reports indicate that PPIases are critical resistance/tolerance factors against various biotic and abiotic stresses in crops [
69,
71,
72,
73,
74,
75]. For example, PPIases trigger both salt tolerance and
P. syringae pv. tabaci resistance in tobacco. Studies reported that PPIase deletion leads to susceptibility toward
P. syringae in
Arabidopsis thaliana and provides resistance against
Plasmodiophora brassicae in
Brassica oleracea [
76]; drought and salt tolerance in sorghum [
66,
67]; drought tolerance in rice [
77]; against fungal pathogen
Leptosphaeria maculans in
Brassica carinata-derived
Brassica napus introgression lines [
78]; against
Xanthomonas campestris in
A. thaliana [
73]. PPIases are involved in a wide range of molecular pathways and play an essential role in protein folding, reactivation of denatured proteins, and restoration of polypeptide active structures. Besides their unique functions, PPIases are part of large chaperone complexes, transmembrane channels responsible for Ca
2+, and other ion transport events. PPIases have a role in various cellular processes such as signal transduction, RNA processing, protein secretion, cell cycle control, development regulation, photosynthesis, and host–pathogen interactions [
79].
It is reported that PPIase has a role in host–pathogen interactions by several mechanisms: (1) Modification of transmembrane and secreted proteins by PPIases of the pathogen to escape or overcome the immune response in intra- or intercellular space of host (most bacteria); (2) stabilization and modification of proteins essential for pathogenesis by PPIases of the host (viruses, protozoa, and some bacteria); (3) suppression of pathogen growth by host PPIases. A study reported that cyclophilin C-CyP (PPIase) with a molecular weight of about 20 kDa isolated from Chinese cabbage (
B. campestris ssp.
pekinensis L.) possesses fungistatic activity. In vitro growth of several fungi, including
Candida albicans,
Rhizoctonia solani,
Botrytis cinerea,
Trichoderma harzianum, and
T. viride was suppressed by C-CyP [
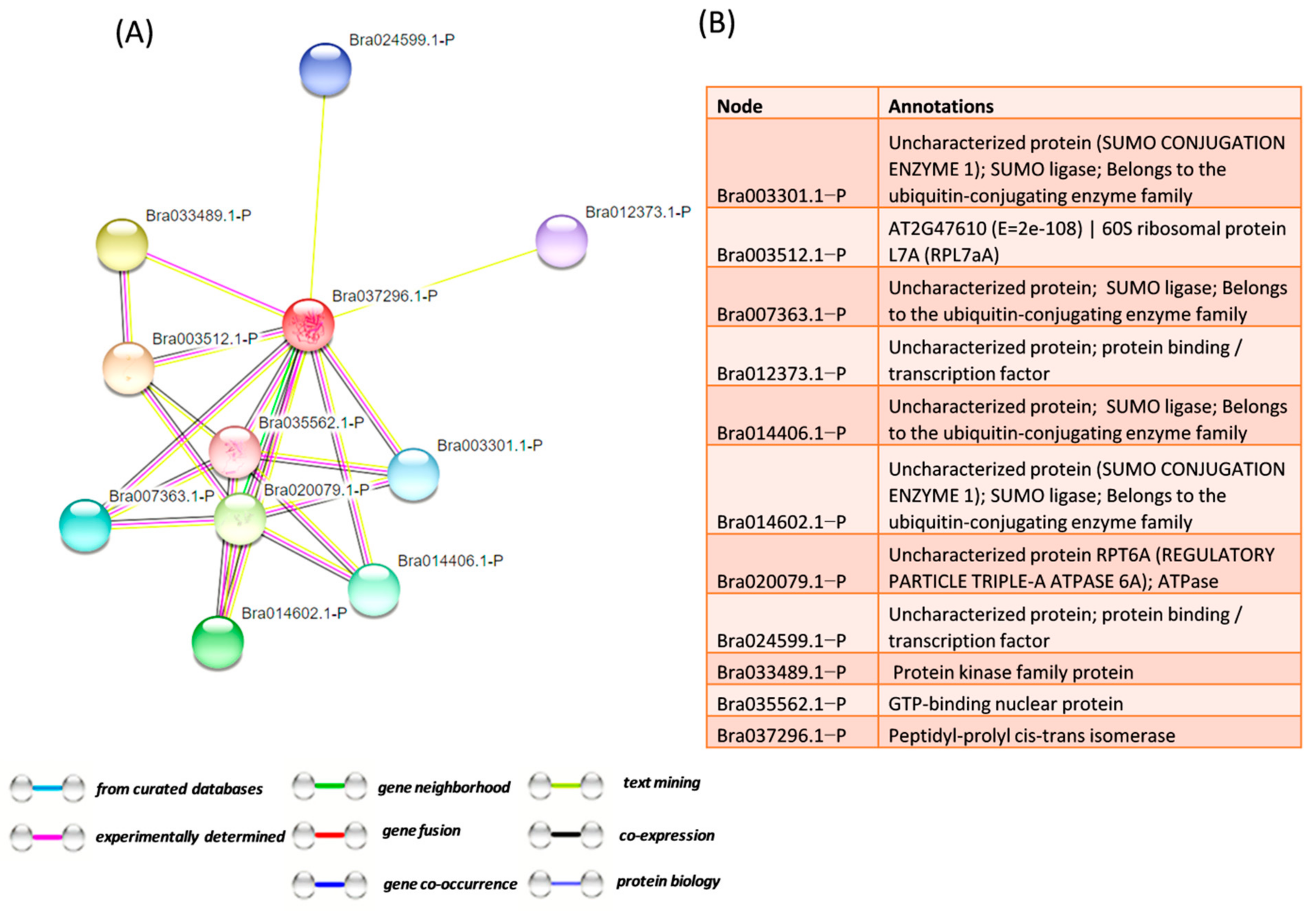
80]. Insilico analysis from this study showed that various signaling proteins are involved in pathogen resistance.
Previous studies indicated that pathogen-mediated host intracellular acidification increased the expression of PPIase.
Sclerotinia sclerotiorum releases oxalic acid inside the invaded host to cause disease in plants. Oxalic acid is a chief virulence/factor without oxalic acid, the pathogen loses its pathogenicity and becomes non-pathogenic/avirulent. Oxalic acid creates an acidic environment inside the invaded host tissue to cause direct toxicity to living cells, suppresses the host antioxidant defense system, consequently escalating the activities of various cell wall-degrading enzymes to disturb host cell wall integrity [
81]. To overcome the adverse effects of oxalic acids, PPIase may act as intracellular pH homeostatic machinery and refold several stress-related proteins to activate H+ extrusion and restore intra-cellular pH of the
S. sclerotiorum invaded host tissue [
82].
Therefore, the present study, apart from being an initial step for further investigation towards the molecular basis of Sclerotinia stem rot resistance, could be beneficial for designing operative breeding programs that might lead to Indian mustard cultivars resistant to this economically important disease.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}