
Post-Transcriptional Modifications of Conserved Nucleotides in the T-Loop of tRNA: A Tale of Functional Convergent Evolution

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Intricate T-D-Loop Anatomy



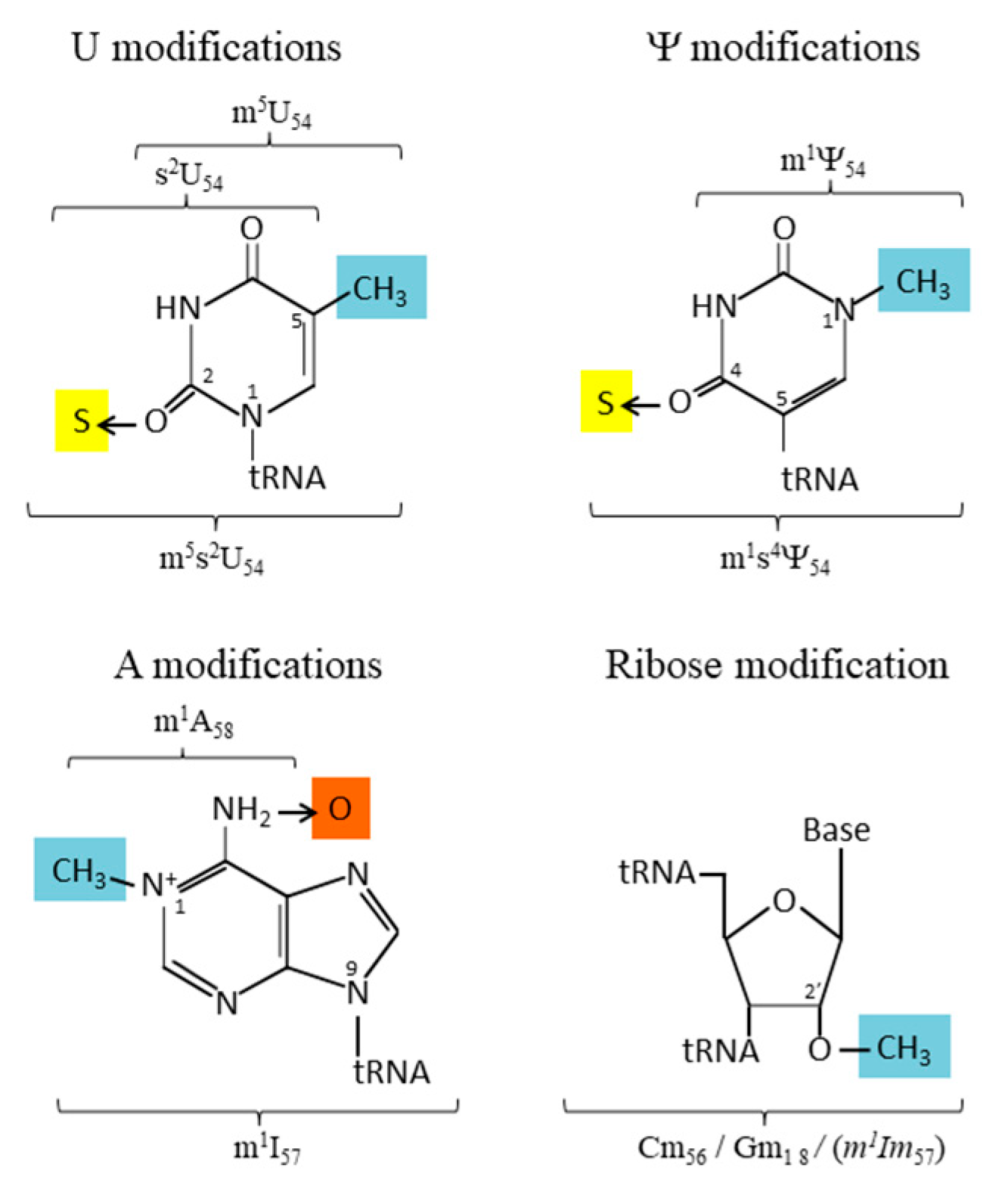
3. Modified Nucleotides within the T-Loop and G18 Stabilize the Elbow of tRNA
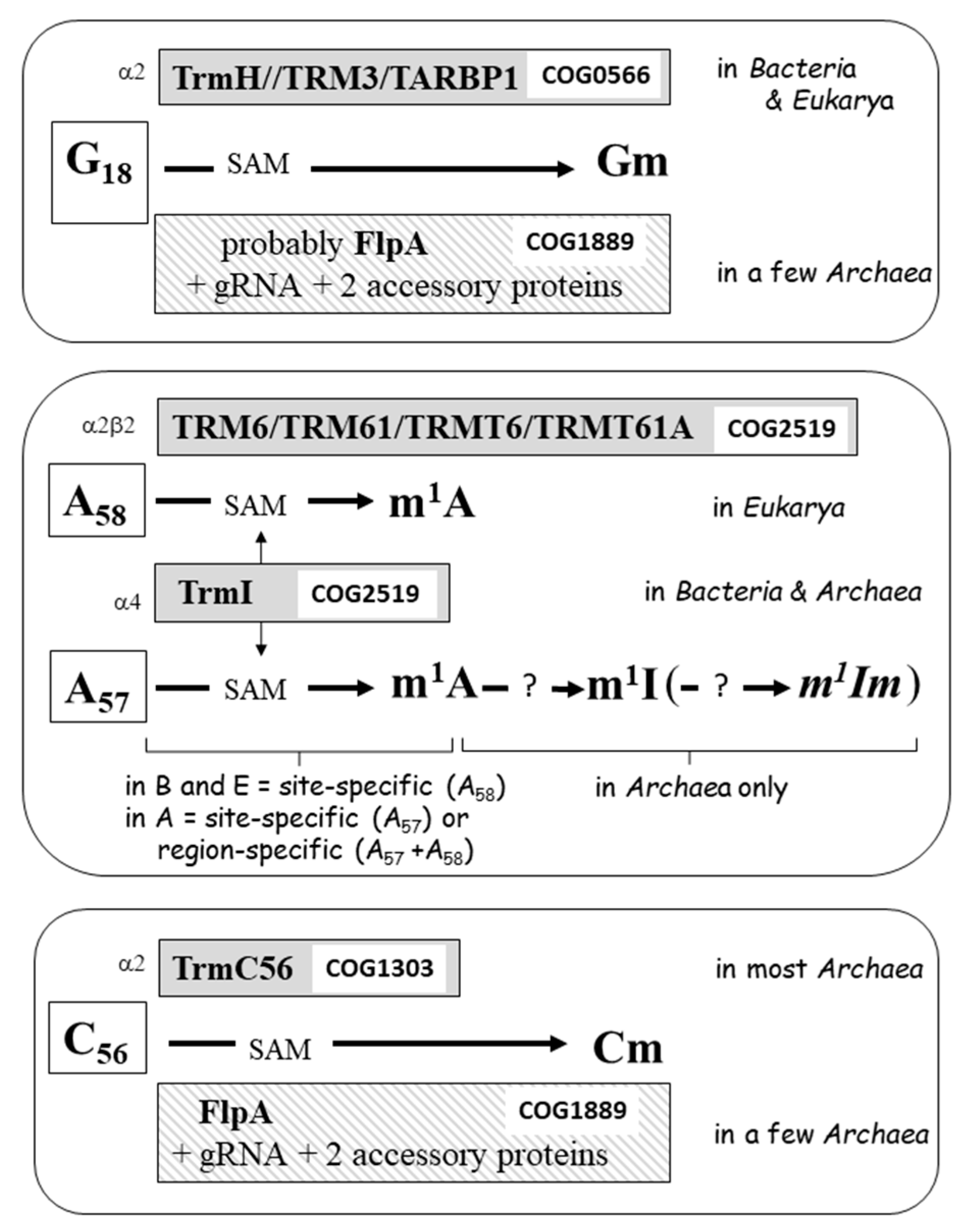
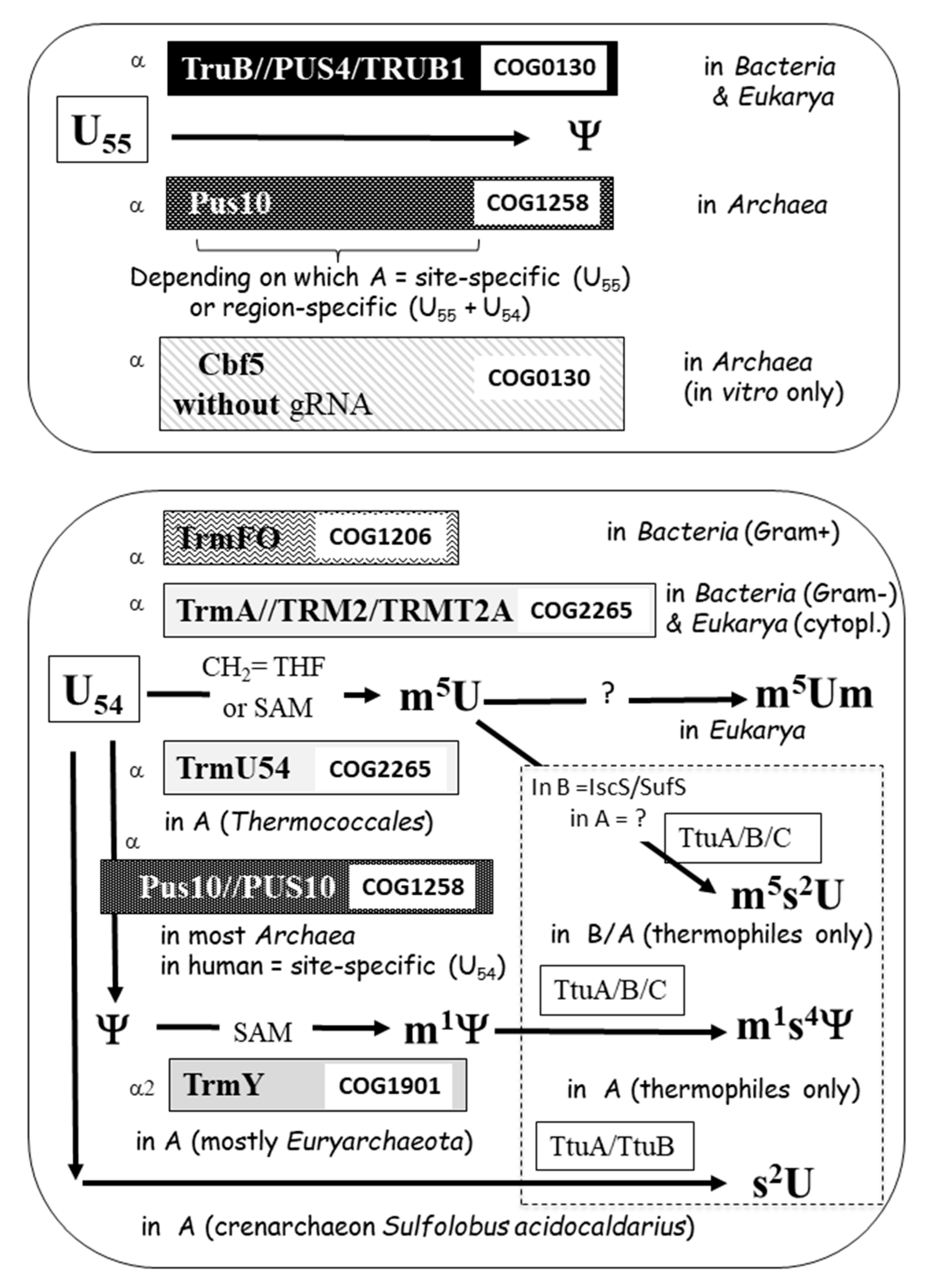
4. The Enzymatic Toolbox for Nucleotide Modifications at Positions 18 (D-Loop) and 54–58 (T-Loop) of tRNA
4.1. G18 Modification in the D-Loop

4.2. Modification of A58 and A57
4.3. Modification of C56
4.4. Modification of U55

4.5. Modification of U54
5. Temporal Order of Nucleotide Modifications and Their Interdependencies
6. tRNA T-Loop Modification Enzymes Can Modify Other T-Loop Containing RNAs
7. Conclusions and Future Prospects: Unraveling the Evolutionary Origin of tRNA Modification Machinery
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Westhof, E.; Auffinger, P. tRNA stucture. In Citable Reviews in the Life Sciences; John Wiley & Sons Ltd.: Chichester, UK, 2001; pp. 1–11. [Google Scholar] [CrossRef]
- Giegé, R.; Jühling, F.; Pütz, J.; Stadler, P.; Sauter, C.; Florentz, C. Structure of transfer RNAs: Similarity and variability. Wiley Interdiscip. Rev. RNA 2012, 3, 37–61. [Google Scholar] [CrossRef]
- Lorenz, C.; Lünse, C.E.; Mörl, M. tRNA Modifications: Impact on Structure and Thermal Adaptation. Biomolecules 2017, 7, 35. [Google Scholar] [CrossRef] [Green Version]
- Väre, V.Y.; Eruysal, E.R.; Narendran, A.; Sarachan, K.L.; Agris, P.F. Chemical and Conformational Diversity of Modified Nucleosides Affects tRNA Structure and Function. Biomolecules 2017, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, S.; Leclerc, F.; Cedergren, R. Structural rules and conformational compensations in the tRNA L-form. J. Mol. Biol. 1997, 266, 269–282. [Google Scholar] [CrossRef]
- Watanabe, Y.; Suematsu, T.; Ohtsuki, T. Losing the stem-loop structure from metazoan mitochondrial tRNAs and co-evolution of interacting factors. Front. Genet. 2014, 5, 109. [Google Scholar] [CrossRef] [Green Version]
- Dick, T.P.; Schamel, W.A. Molecular evolution of transfer RNA from two precursor hairpins: Implications for the origin of protein synthesis. J. Mol. Evol. 1995, 41, 1–9. [Google Scholar] [CrossRef]
- Ardell, D.H. Computational analysis of tRNA identity. FEBS Lett. 2010, 584, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Jühling, F.; Mörl, M.; Hartmann, R.K.; Sprinzl, M.; Stadler, P.F.; Pütz, J. tRNAdb 2009: Compilation of tRNA sequences and tRNA genes. Nucleic Acids Res. 2009, 37, D159–D162. [Google Scholar] [CrossRef] [Green Version]
- Boccaletto, P.; Machnicka, M.A.; Purta, E.; Piatkowski, P.; Baginski, B.; Wirecki, T.K.; de Crecy-Lagard, V.; Ross, R.; Limbach, P.A.; Kotter, A.; et al. MODOMICS: A database of RNA modification pathways. 2017 update. Nucleic Acids Res. 2018, 46, D303–D307. [Google Scholar] [CrossRef]
- Westhof, E.; Dumas, P.; Moras, D. Crystallographic refinement of yeast aspartic acid transfer RNA. J. Mol. Biol. 1985, 184, 119–145. [Google Scholar] [CrossRef]
- Romby, P.; Carbon, P.; Westhof, E.; Ehresmann, C.; Ebel, J.P.; Ehresmann, B.; Giegé, R. Importance of conserved residues for the conformation of the T-loop in tRNAs. J. Biomol. Struct. Dyn. 1987, 5, 669–687. [Google Scholar] [CrossRef]
- Shi, H.; Moore, P.B. The crystal structure of yeast phenylalanine tRNA at 1.93 A resolution: A classic structure revisited. RNA 2000, 6, 1091–1105. [Google Scholar] [CrossRef] [Green Version]
- Basavappa, R.; Sigler, P.B. The 3 A crystal structure of yeast initiator tRNA: Functional implications in initiator/elongator discrimination. EMBO J. 1991, 10, 3105–3111. [Google Scholar] [CrossRef]
- Quigley, G.J.; Rich, A. Structural domains of transfer RNA molecules. Science 1976, 194, 796–806. [Google Scholar] [CrossRef]
- Levitt, M. Detailed molecular model for transfer ribonucleic acid. Nature 1969, 224, 759–763. [Google Scholar] [CrossRef]
- Oliva, R.; Cavallo, L.; Tramontano, A. Accurate energies of hydrogen bonded nucleic acid base pairs and triplets in tRNA tertiary interactions. Nucleic Acids Res. 2006, 34, 865–879. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.R. Stabilization of RNA stacking by pseudouridine. Nucleic Acids Res. 1995, 23, 5020–5026. [Google Scholar] [CrossRef]
- Westhof, E.; Dumas, P.; Moras, D. Hydration of transfer RNA molecules: A crystallographic study. Biochimie 1988, 70, 145–165. [Google Scholar] [CrossRef]
- Hori, H.; Kawamura, T.; Awai, T.; Ochi, A.; Yamagami, R.; Tomikawa, C.; Hirata, A. Transfer RNA Modification Enzymes from Thermophiles and Their Modified Nucleosides in tRNA. Microorganisms 2018, 6, 110. [Google Scholar] [CrossRef] [Green Version]
- Rose, S.; Auxilien, S.; Havelund, J.F.; Kirpekar, F.; Huber, H.; Grosjean, H.; Douthwaite, S. The hyperthermophilic partners Nanoarchaeum and Ignicoccus stabilize their tRNA T-loops via different but structurally equivalent modifications. Nucleic Acids Res. 2020, 48, 6906–6918. [Google Scholar] [CrossRef]
- Shigi, N. Biosynthesis and functions of sulfur modifications in tRNA. Front. Genet. 2014, 5, 67. [Google Scholar] [CrossRef] [Green Version]
- Kawai, G.; Yamamoto, Y.; Kamimura, T.; Masegi, T.; Sekine, M.; Hata, T.; Iimori, T.; Watanabe, T.; Miyazawa, T.; Yokoyama, S. Conformational rigidity of specific pyrimidine residues in tRNA arises from posttranscriptional modifications that enhance steric interaction between the base and the 2′-hydroxyl group. Biochemistry 1992, 31, 1040–1046. [Google Scholar] [CrossRef]
- Clouet-d’Orval, B.; Gaspin, C.; Mougin, A. Two different mechanisms for tRNA ribose methylation in Archaea: A short survey. Biochimie 2005, 87, 889–895. [Google Scholar] [CrossRef]
- Edmonds, C.G.; Crain, P.F.; Gupta, R.; Hashizume, T.; Hocart, C.H.; Kowalak, J.A.; Pomerantz, S.C.; Stetter, K.O.; McCloskey, J.A. Posttranscriptional modification of tRNA in thermophilic archaea (Archaebacteria). J. Bacteriol 1991, 173, 3138–3148. [Google Scholar] [CrossRef] [Green Version]
- Keller, P.; Freund, I.; Marchand, V.; Bec, G.; Huang, R.; Motorin, Y.; Eigenbrod, T.; Dalpke, A.; Helm, M. Double methylation of tRNA-U54 to 2′-O-methylthymidine (Tm) synergistically decreases immune response by Toll-like receptor 7. Nucleic Acids Res. 2018, 46, 9764–9775. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R. Halobacterium volcanii tRNAs. Identification of 41 tRNAs covering all amino acids, and the sequences of 33 class I tRNAs. J. Biol. Chem. 1984, 259, 9461–9471. [Google Scholar] [CrossRef]
- Grosjean, H.; Gaspin, C.; Marck, C.; Decatur, W.A.; de Crécy-Lagard, V. RNomics and Modomics in the halophilic archaea Haloferax volcanii: Identification of RNA modification genes. BMC Genom. 2008, 9, 470. [Google Scholar] [CrossRef] [Green Version]
- Wolff, P.; Villette, C.; Zumsteg, J.; Heintz, D.; Antoine, L.; Chane-Woon-Ming, B.; Droogmans, L.; Grosjean, H.; Westhof, E. Comparative patterns of modified nucleotides in individual tRNA species from a mesophilic and two thermophilic archaea. RNA 2020, 26, 1957–1975. [Google Scholar] [CrossRef]
- Nobles, K.N.; Yarian, C.S.; Liu, G.; Guenther, R.H.; Agris, P.F. Highly conserved modified nucleosides influence Mg2+-dependent tRNA folding. Nucleic Acids Res. 2002, 30, 4751–4760. [Google Scholar] [CrossRef] [Green Version]
- Ayadi, L.; Galvanin, A.; Pichot, F.; Marchand, V.; Motorin, Y. RNA ribose methylation (2′-O-methylation): Occurrence, biosynthesis and biological functions. Biochim. Biophys. Acta Gene. Regul. Mech. 2019, 1862, 253–269. [Google Scholar] [CrossRef]
- Nureki, O.; Watanabe, K.; Fukai, S.; Ishii, R.; Endo, Y.; Hori, H.; Yokoyama, S. Deep knot structure for construction of active site and cofactor binding site of tRNA modification enzyme. Structure 2004, 12, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Galperin, M.Y.; Wolf, Y.I.; Makarova, K.S.; Vera Alvarez, R.; Landsman, D.; Koonin, E.V. COG database update: Focus on microbial diversity, model organisms, and widespread pathogens. Nucleic Acids Res. 2020. [Google Scholar] [CrossRef]
- Hori, H.; Yamazaki, N.; Matsumoto, T.; Watanabe, Y.; Ueda, T.; Nishikawa, K.; Kumagai, I.; Watanabe, K. Substrate recognition of tRNA (Guanosine-2′-)-methyltransferase from Thermus thermophilus HB27. J. Biol. Chem. 1998, 273, 25721–25727. [Google Scholar] [CrossRef] [Green Version]
- Persson, B.C.; Jäger, G.; Gustafsson, C. The spoU gene of Escherichia coli, the fourth gene of the spoT operon, is essential for tRNA (Gm18) 2′-O-methyltransferase activity. Nucleic Acids Res. 1997, 25, 4093–4097. [Google Scholar] [CrossRef] [Green Version]
- Hori, H.; Kubota, S.; Watanabe, K.; Kim, J.M.; Ogasawara, T.; Sawasaki, T.; Endo, Y. Aquifex aeolicus tRNA (Gm18) methyltransferase has unique substrate specificity. TRNA recognition mechanism of the enzyme. J. Biol. Chem. 2003, 278, 25081–25090. [Google Scholar] [CrossRef] [Green Version]
- Hori, H. Transfer RNA methyltransferases with a SpoU-TrmD (SPOUT) fold and their modified nucleosides in tRNA. Biomolecules 2017, 7, 23. [Google Scholar] [CrossRef] [Green Version]
- Cavaillé, J.; Chetouani, F.; Bachellerie, J.P. The yeast Saccharomyces cerevisiae YDL112w ORF encodes the putative 2′-O-ribose methyltransferase catalyzing the formation of Gm18 in tRNAs. RNA 1999, 5, 66–81. [Google Scholar] [CrossRef] [Green Version]
- De Crécy-Lagard, V.; Boccaletto, P.; Mangleburg, C.G.; Sharma, P.; Lowe, T.M.; Leidel, S.A.; Bujnicki, J.M. Matching tRNA modifications in humans to their known and predicted enzymes. Nucleic Acids Res. 2019, 47, 2143–2159. [Google Scholar] [CrossRef]
- Omer, A.D.; Lowe, T.M.; Russell, A.G.; Ebhardt, H.; Eddy, S.R.; Dennis, P.P. Homologs of small nucleolar RNAs in Archaea. Science 2000, 288, 517–522. [Google Scholar] [CrossRef] [Green Version]
- Lui, L.M.; Uzilov, A.V.; Bernick, D.L.; Corredor, A.; Lowe, T.M.; Dennis, P.P. Methylation guide RNA evolution in archaea: Structure, function and genomic organization of 110 C/D box sRNA families across six Pyrobaculum species. Nucleic Acids Res. 2018, 46, 5678–5691. [Google Scholar] [CrossRef]
- Anderson, J.; Droogmans, L. Biosynthesis and function of 1-methyladenosine in transfer RNA. In Fine-Tuning of RNA Functions by Modification and Editing; Grosjean, H., Ed.; Springer: Heidelberg/Berlin, Germany, 2005; pp. 121–139. [Google Scholar]
- Swinehart, W.E.; Jackman, J.E. Diversity in mechanism and function of tRNA methyltransferases. RNA Biol. 2015, 12, 398–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dégut, C.; Ponchon, L.; Folly-Klan, M.; Barraud, P.; Tisné, C. The m1A(58) modification in eubacterial tRNA: An overview of tRNA recognition and mechanism of catalysis by TrmI. Biophys. Chem. 2016, 210, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Oerum, S.; Dégut, C.; Barraud, P.; Tisné, C. m1A Post-Transcriptional Modification in tRNAs. Biomolecules 2017, 7, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shima, H.; Igarashi, K. N 1-methyladenosine (m1A) RNA modification: The key to ribosome control. J. Biochem. 2020, 167, 535–539. [Google Scholar] [CrossRef]
- Droogmans, L.; Roovers, M.; Bujnicki, J.M.; Tricot, C.; Hartsch, T.; Stalon, V.; Grosjean, H. Cloning and characterization of tRNA (m1A58) methyltransferase (TrmI) from Thermus thermophilus HB27, a protein required for cell growth at extreme temperatures. Nucleic Acids Res. 2003, 31, 2148–2156. [Google Scholar] [CrossRef] [Green Version]
- Barraud, P.; Golinelli-Pimpaneau, B.; Atmanene, C.; Sanglier, S.; Van Dorsselaer, A.; Droogmans, L.; Dardel, F.; Tisné, C. Crystal structure of Thermus thermophilus tRNA m1A58 methyltransferase and biophysical characterization of its interaction with tRNA. J. Mol. Biol. 2008, 377, 535–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takuma, H.; Ushio, N.; Minoji, M.; Kazayama, A.; Shigi, N.; Hirata, A.; Tomikawa, C.; Ochi, A.; Hori, H. Substrate tRNA recognition mechanism of eubacterial tRNA (m1A58) methyltransferase (TrmI). J. Biol. Chem. 2015, 290, 5912–5925. [Google Scholar] [CrossRef] [Green Version]
- Grosjean, H.; Edqvist, J.; Stråby, K.B.; Giegé, R. Enzymatic formation of modified nucleosides in tRNA: Dependence on tRNA architecture. J. Mol. Biol. 1996, 255, 67–85. [Google Scholar] [CrossRef]
- Anderson, J.; Phan, L.; Cuesta, R.; Carlson, B.A.; Pak, M.; Asano, K.; Björk, G.R.; Tamame, M.; Hinnebusch, A.G. The essential Gcd10p-Gcd14p nuclear complex is required for 1-methyladenosine modification and maturation of initiator methionyl-tRNA. Genes Dev. 1998, 12, 3650–3662. [Google Scholar] [CrossRef] [Green Version]
- Ozanick, S.; Krecic, A.; Andersland, J.; Anderson, J.T. The bipartite structure of the tRNA m1A58 methyltransferase from S. cerevisiae is conserved in humans. RNA 2005, 11, 1281–1290. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhu, Y.; Wang, C.; Fan, X.; Jiang, X.; Ebrahimi, M.; Qiao, Z.; Niu, L.; Teng, M.; Li, X. Crystal structure of the two-subunit tRNA m(1)A58 methyltransferase TRM6-TRM61 from Saccharomyces cerevisiae. Sci. Rep. 2016, 6, 32562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guy, M.P.; Phizicky, E.M. Two-subunit enzymes involved in eukaryotic post-transcriptional tRNA modification. RNA Biol. 2014, 11, 1608–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roovers, M.; Wouters, J.; Bujnicki, J.M.; Tricot, C.; Stalon, V.; Grosjean, H.; Droogmans, L. A primordial RNA modification enzyme: The case of tRNA (m1A) methyltransferase. Nucleic Acids Res. 2004, 32, 465–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guelorget, A.; Barraud, P.; Tisné, C.; Golinelli-Pimpaneau, B. Structural comparison of tRNA m(1)A58 methyltransferases revealed different molecular strategies to maintain their oligomeric architecture under extreme conditions. BMC Struct. Biol. 2011, 11, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guelorget, A.; Roovers, M.; Guérineau, V.; Barbey, C.; Li, X.; Golinelli-Pimpaneau, B. Insights into the hyperthermostability and unusual region-specificity of archaeal Pyrococcus abyssi tRNA m1A57/58 methyltransferase. Nucleic Acids Res. 2010, 38, 6206–6218. [Google Scholar] [CrossRef] [Green Version]
- Hamdane, D.; Guelorget, A.; Guérineau, V.; Golinelli-Pimpaneau, B. Dynamics of RNA modification by a multi-site-specific tRNA methyltransferase. Nucleic Acids Res. 2014, 42, 11697–11706. [Google Scholar] [CrossRef] [Green Version]
- Grosjean, H.; Constantinesco, F.; Foiret, D.; Benachenhou, N. A novel enzymatic pathway leading to 1-methylinosine modification in Haloferax volcanii tRNA. Nucleic Acids Res. 1995, 23, 4312–4319. [Google Scholar] [CrossRef] [Green Version]
- Dixit, S.; Henderson, J.C.; Alfonzo, J.D. Multi-Substrate Specificity and the Evolutionary Basis for Interdependence in tRNA Editing and Methylation Enzymes. Front. Genet. 2019, 10, 104. [Google Scholar] [CrossRef]
- Agris, P.F. The importance of being modified: Roles of modified nucleosides and Mg2+ in RNA structure and function. Prog. Nucleic Acid Res. Mol. Biol. 1996, 53, 79–129. [Google Scholar] [CrossRef]
- Auxilien, S.; Keith, G.; Le Grice, S.F.; Darlix, J.L. Role of post-transcriptional modifications of primer tRNALys,3 in the fidelity and efficacy of plus strand DNA transfer during HIV-1 reverse transcription. J. Biol. Chem. 1999, 274, 4412–4420. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Clark, W.; Luo, G.; Wang, X.; Fu, Y.; Wei, J.; Wang, X.; Hao, Z.; Dai, Q.; Zheng, G.; et al. ALKBH1-Mediated tRNA Demethylation Regulates Translation. Cell 2016, 167, 816–828.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Jia, G. Reversible RNA Modification N(1)-methyladenosine (m(1)A) in mRNA and tRNA. Genom. Proteom. Bioinform. 2018, 16, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Renalier, M.H.; Joseph, N.; Gaspin, C.; Thebault, P.; Mougin, A. The Cm56 tRNA modification in archaea is catalyzed either by a specific 2′-O-methylase, or a C/D sRNP. RNA 2005, 11, 1051–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constantinesco, F.; Motorin, Y.; Grosjean, H. Transfer RNA modification enzymes from Pyrococcus furiosus: Detection of the enzymatic activities in vitro. Nucleic Acids Res. 1999, 27, 1308–1315. [Google Scholar] [CrossRef]
- Tkaczuk, K.L.; Dunin-Horkawicz, S.; Purta, E.; Bujnicki, J.M. Structural and evolutionary bioinformatics of the SPOUT superfamily of methyltransferases. BMC Bioinform. 2007, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Tomkuvienė, M.; Ličytė, J.; Olendraitė, I.; Liutkevičiūtė, Z.; Clouet-d’Orval, B.; Klimašauskas, S. Archaeal fibrillarin-Nop5 heterodimer 2′-O-methylates RNA independently of the C/D guide RNP particle. RNA 2017, 23, 1329–1337. [Google Scholar] [CrossRef] [PubMed]
- Nurse, K.; Wrzesinski, J.; Bakin, A.; Lane, B.G.; Ofengand, J. Purification, cloning, and properties of the tRNA psi 55 synthase from Escherichia coli. RNA 1995, 1, 102–112. [Google Scholar]
- Becker, H.F.; Motorin, Y.; Planta, R.J.; Grosjean, H. The yeast gene YNL292w encodes a pseudouridine synthase (Pus4) catalyzing the formation of psi55 in both mitochondrial and cytoplasmic tRNAs. Nucleic Acids Res. 1997, 25, 4493–4499. [Google Scholar] [CrossRef]
- Zucchini, C.; Strippoli, P.; Biolchi, A.; Solmi, R.; Lenzi, L.; D’Addabbo, P.; Carinci, P.; Valvassori, L. The human TruB family of pseudouridine synthase genes, including the Dyskeratosis Congenita 1 gene and the novel member TRUB1. Int. J. Mol. Med. 2003, 11, 697–704. [Google Scholar] [CrossRef]
- Pan, H.; Agarwalla, S.; Moustakas, D.T.; Finer-Moore, J.; Stroud, R.M. Structure of tRNA pseudouridine synthase TruB and its RNA complex: RNA recognition through a combination of rigid docking and induced fit. Proc. Natl. Acad. Sci. USA 2003, 100, 12648–12653. [Google Scholar] [CrossRef] [Green Version]
- Phannachet, K.; Elias, Y.; Huang, R.H. Dissecting the roles of a strictly conserved tyrosine in substrate recognition and catalysis by pseudouridine 55 synthase. Biochemistry 2005, 44, 15488–15494. [Google Scholar] [CrossRef] [PubMed]
- Hoang, C.; Hamilton, C.S.; Mueller, E.G.; Ferré-D’Amaré, A.R. Precursor complex structure of pseudouridine synthase TruB suggests coupling of active site perturbations to an RNA-sequestering peripheral protein domain. Protein Sci. 2005, 14, 2201–2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hur, S.; Stroud, R.M.; Finer-Moore, J. Substrate recognition by RNA 5-methyluridine methyltransferases and pseudouridine synthases: A structural perspective. J. Biol. Chem. 2006, 281, 38969–38973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, X.; Yu, M.; Ivanetich, K.M.; Santi, D.V. Molecular recognition of tRNA by tRNA pseudouridine 55 synthase. Biochemistry 1998, 37, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Keffer-Wilkes, L.C.; Veerareddygari, G.R.; Kothe, U. RNA modification enzyme TruB is a tRNA chaperone. Proc. Natl. Acad. Sci. USA 2016, 113, 14306–14311. [Google Scholar] [CrossRef] [Green Version]
- Hamma, T.; Ferré-D’Amaré, A.R. The box H/ACA ribonucleoprotein complex: Interplay of RNA and protein structures in post-transcriptional RNA modification. J. Biol. Chem. 2010, 285, 805–809. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.T.; Meier, U.T. RNA-guided isomerization of uridine to pseudouridine—Pseudouridylation. RNA Biol. 2014, 11, 1483–1494. [Google Scholar] [CrossRef] [Green Version]
- Spenkuch, F.; Motorin, Y.; Helm, M. Pseudouridine: Still mysterious, but never a fake (uridine)! RNA Biol. 2014, 11, 1540–1554. [Google Scholar] [CrossRef] [Green Version]
- Roovers, M.; Hale, C.; Tricot, C.; Terns, M.P.; Terns, R.M.; Grosjean, H.; Droogmans, L. Formation of the conserved pseudouridine at position 55 in archaeal tRNA. Nucleic Acids Res. 2006, 34, 4293–4301. [Google Scholar] [CrossRef]
- Gurha, P.; Joardar, A.; Chaurasia, P.; Gupta, R. Differential roles of archaeal box H/ACA proteins in guide RNA-dependent and independent pseudouridine formation. RNA Biol. 2007, 4, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Gurha, P.; Gupta, R. Archaeal Pus10 proteins can produce both pseudouridine 54 and 55 in tRNA. RNA 2008, 14, 2521–2527. [Google Scholar] [CrossRef] [Green Version]
- Tillault, A.S.; Fourmann, J.B.; Loegler, C.; Wieden, H.J.; Kothe, U.; Charpentier, B. Contribution of two conserved histidines to the dual activity of archaeal RNA guide-dependent and -independent pseudouridine synthase Cbf5. RNA 2015, 21, 1233–1239. [Google Scholar] [CrossRef] [Green Version]
- Muller, S.; Fourmann, J.B.; Loegler, C.; Charpentier, B.; Branlant, C. Identification of determinants in the protein partners aCBF5 and aNOP10 necessary for the tRNA:Psi55-synthase and RNA-guided RNA:Psi-synthase activities. Nucleic Acids Res. 2007, 35, 5610–5624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujikane, R.; Behm-Ansmant, I.; Tillault, A.S.; Loegler, C.; Igel-Bourguignon, V.; Marguet, E.; Forterre, P.; Branlant, C.; Motorin, Y.; Charpentier, B. Contribution of protein Gar1 to the RNA-guided and RNA-independent rRNA:Ψ-synthase activities of the archaeal Cbf5 protein. Sci. Rep. 2018, 8, 13815. [Google Scholar] [CrossRef] [PubMed]
- Kamalampeta, R.; Kothe, U. Archaeal proteins Nop10 and Gar1 increase the catalytic activity of Cbf5 in pseudouridylating tRNA. Sci. Rep. 2012, 2, 663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaby, I.K.; Majumder, M.; Chatterjee, K.; Jana, S.; Grosjean, H.; de Crécy-Lagard, V.; Gupta, R. Pseudouridine formation in archaeal RNAs: The case of Haloferax volcanii. RNA 2011, 17, 1367–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, Y.; Gray, M.W. Evolutionary appearance of genes encoding proteins associated with box H/ACA snoRNAs: cbf5p in Euglena gracilis, an early diverging eukaryote, and candidate Gar1p and Nop10p homologs in archaebacteria. Nucleic Acids Res. 2000, 28, 2342–2352. [Google Scholar] [CrossRef] [Green Version]
- Fitzek, E.; Joardar, A.; Gupta, R.; Geisler, M. Evolution of Eukaryal and Archaeal Pseudouridine Synthase Pus10. J. Mol. Evol. 2018, 86, 77–89. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Deogharia, M.; Gupta, R. Mammalian nuclear TRUB1, mitochondrial TRUB2 and cytoplasmic PUS10 produce conserved pseudouridine 55 in different sets of tRNA. RNA 2020. [Google Scholar] [CrossRef]
- Ny, T.; Björk, G.R. Cloning and restriction mapping of the trmA gene coding for transfer ribonucleic acid (5-methyluridine)-methyltransferase in Escherichia coli K-12. J. Bacteriol. 1980, 142, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Persson, B.C.; Gustafsson, C.; Berg, D.E.; Björk, G.R. The gene for a tRNA modifying enzyme, m5U54-methyltransferase, is essential for viability in Escherichia coli. Proc. Natl. Acad. Sci. USA 1992, 89, 3995–3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kealey, J.T.; Gu, X.; Santi, D.V. Enzymatic mechanism of tRNA (m5U54)methyltransferase. Biochimie 1994, 76, 1133–1142. [Google Scholar] [CrossRef]
- Keffer-Wilkes, L.C.; Soon, E.F.; Kothe, U. The methyltransferase TrmA facilitates tRNA folding through interaction with its RNA-binding domain. Nucleic Acids Res. 2020, 48, 7981–7990. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, M.E.; Johansson, J.O.; von Pawel-Rammingen, U.; Byström, A.S. Identification of the TRM2 gene encoding the tRNA(m5U54)methyltransferase of Saccharomyces cerevisiae. RNA 2000, 6, 844–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, J.M.; Emmett, W.; Mozos, I.R.; Kotter, A.; Helm, M.; Ule, J.; Hussain, S. FICC-Seq: A method for enzyme-specified profiling of methyl-5-uridine in cellular RNA. Nucleic Acids Res. 2019, 47, e113. [Google Scholar] [CrossRef]
- Urbonavicius, J.; Brochier-Armanet, C.; Skouloubris, S.; Myllykallio, H.; Grosjean, H. In Vitro detection of the enzymatic activity of folate-dependent tRNA (Uracil-54,-C5)-methyltransferase: Evolutionary implications. Methods Enzymol. 2007, 425, 103–119. [Google Scholar] [CrossRef]
- Urbonavicius, J.; Skouloubris, S.; Myllykallio, H.; Grosjean, H. Identification of a novel gene encoding a flavin-dependent tRNA:m5U methyltransferase in bacteria—Evolutionary implications. Nucleic Acids Res. 2005, 33, 3955–3964. [Google Scholar] [CrossRef]
- Hamdane, D.; Grosjean, H.; Fontecave, M. Flavin-Dependent Methylation of RNAs: Complex Chemistry for a Simple Modification. J. Mol. Biol. 2016, 428, 4867–4881. [Google Scholar] [CrossRef]
- Grosjean, H.; Breton, M.; Sirand-Pugnet, P.; Tardy, F.; Thiaucourt, F.; Citti, C.; Barré, A.; Yoshizawa, S.; Fourmy, D.; de Crécy-Lagard, V.; et al. Predicting the minimal translation apparatus: Lessons from the reductive evolution of mollicutes. PLoS Genet. 2014, 10, e1004363. [Google Scholar] [CrossRef]
- Sirand-Pugnet, P.; Brégeon, D.; Béven, L.; Goyenvalle, C.; Blanchard, A.; Rose, S.; Grosjean, H.; Douthwaite, S.; Hamdane, D.; Crécy-Lagard, V. Reductive Evolution and Diversification of C5-Uracil Methylation in the Nucleic Acids of Mollicutes. Biomolecules 2020, 10, 587. [Google Scholar] [CrossRef] [Green Version]
- Urbonavicius, J.; Auxilien, S.; Walbott, H.; Trachana, K.; Golinelli-Pimpaneau, B.; Brochier-Armanet, C.; Grosjean, H. Acquisition of a bacterial RumA-type tRNA(uracil-54, C5)-methyltransferase by Archaea through an ancient horizontal gene transfer. Mol. Microbiol. 2008, 67, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.T.; Agarwalla, S.; Stroud, R.M. Crystal structure of RumA, an iron-sulfur cluster containing E. coli ribosomal RNA 5-methyluridine methyltransferase. Structure 2004, 12, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auxilien, S.; Rasmussen, A.; Rose, S.; Brochier-Armanet, C.; Husson, C.; Fourmy, D.; Grosjean, H.; Douthwaite, S. Specificity shifts in the rRNA and tRNA nucleotide targets of archaeal and bacterial m5U methyltransferases. RNA 2011, 17, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengupta, R.; Vainauskas, S.; Yarian, C.; Sochacka, E.; Malkiewicz, A.; Guenther, R.H.; Koshlap, K.M.; Agris, P.F. Modified constructs of the tRNA TPsiC domain to probe substrate conformational requirements of m(1)A(58) and m(5)U(54) tRNA methyltransferases. Nucleic Acids Res. 2000, 28, 1374–1380. [Google Scholar] [CrossRef] [Green Version]
- Alian, A.; Lee, T.T.; Griner, S.L.; Stroud, R.M.; Finer-Moore, J. Structure of a TrmA-RNA complex: A consensus RNA fold contributes to substrate selectivity and catalysis in m5U methyltransferases. Proc. Natl. Acad. Sci. USA 2008, 105, 6876–6881. [Google Scholar] [CrossRef] [Green Version]
- Walbott, H.; Leulliot, N.; Grosjean, H.; Golinelli-Pimpaneau, B. The crystal structure of Pyrococcus abyssi tRNA (uracil-54, C5)-methyltransferase provides insights into its tRNA specificity. Nucleic Acids Res. 2008, 36, 4929–4940. [Google Scholar] [CrossRef]
- Yamagami, R.; Yamashita, K.; Nishimasu, H.; Tomikawa, C.; Ochi, A.; Iwashita, C.; Hirata, A.; Ishitani, R.; Nureki, O.; Hori, H. The tRNA recognition mechanism of folate/FAD-dependent tRNA methyltransferase (TrmFO). J. Biol. Chem. 2012, 287, 42480–42494. [Google Scholar] [CrossRef] [Green Version]
- Gross, H.J.; Simsek, M.; Raba, M.; Limburg, K.; Heckman, J.; Raj Bhandary, U.L. 2′-O-methyl ribothymidine: A component of rabbit liver lysine transfer RNA. Nucleic Acids Res. 1974, 1, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Shigi, N.; Sakaguchi, Y.; Asai, S.; Suzuki, T.; Watanabe, K. Common thiolation mechanism in the biosynthesis of tRNA thiouridine and sulphur-containing cofactors. EMBO J. 2008, 27, 3267–3278. [Google Scholar] [CrossRef] [Green Version]
- Čavužić, M.; Liu, Y. Biosynthesis of Sulfur-Containing tRNA Modifications: A Comparison of Bacterial, Archaeal, and Eukaryotic Pathways. Biomolecules 2017, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Shigi, N.; Sakaguchi, Y.; Suzuki, T.; Watanabe, K. Identification of two tRNA thiolation genes required for cell growth at extremely high temperatures. J. Biol. Chem. 2006, 281, 14296–14306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigi, N.; Suzuki, T.; Tamakoshi, M.; Oshima, T.; Watanabe, K. Conserved bases in the TPsi C loop of tRNA are determinants for thermophile-specific 2-thiouridylation at position 54. J. Biol. Chem. 2002, 277, 39128–39135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomikawa, C.; Yokogawa, T.; Kanai, T.; Hori, H. N7-Methylguanine at position 46 (m7G46) in tRNA from Thermus thermophilus is required for cell viability at high temperatures through a tRNA modification network. Nucleic Acids Res. 2010, 38, 942–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joardar, A.; Jana, S.; Fitzek, E.; Gurha, P.; Majumder, M.; Chatterjee, K.; Geisler, M.; Gupta, R. Role of forefinger and thumb loops in production of Ψ54 and Ψ55 in tRNAs by archaeal Pus10. RNA 2013, 19, 1279–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deogharia, M.; Mukhopadhyay, S.; Joardar, A.; Gupta, R. The human ortholog of archaeal Pus10 produces pseudouridine 54 in select tRNAs where its recognition sequence contains a modified residue. RNA 2019, 25, 336–351. [Google Scholar] [CrossRef] [Green Version]
- McCleverty, C.J.; Hornsby, M.; Spraggon, G.; Kreusch, A. Crystal structure of human Pus10, a novel pseudouridine synthase. J. Mol. Biol. 2007, 373, 1243–1254. [Google Scholar] [CrossRef]
- Aravind, L.; Koonin, E.V. THUMP—A predicted RNA-binding domain shared by 4-thiouridine, pseudouridine synthases and RNA methylases. Trends Biochem. Sci. 2001, 26, 215–217. [Google Scholar] [CrossRef]
- Gabant, G.; Auxilien, S.; Tuszynska, I.; Locard, M.; Gajda, M.J.; Chaussinand, G.; Fernandez, B.; Dedieu, A.; Grosjean, H.; Golinelli-Pimpaneau, B.; et al. THUMP from archaeal tRNA:m22G10 methyltransferase, a genuine autonomously folding domain. Nucleic Acids Res. 2006, 34, 2483–2494. [Google Scholar] [CrossRef]
- Waterman, D.G.; Ortiz-Lombardía, M.; Fogg, M.J.; Koonin, E.V.; Antson, A.A. Crystal structure of Bacillus anthracis ThiI, a tRNA-modifying enzyme containing the predicted RNA-binding THUMP domain. J. Mol. Biol. 2006, 356, 97–110. [Google Scholar] [CrossRef]
- Fislage, M.; Roovers, M.; Tuszynska, I.; Bujnicki, J.M.; Droogmans, L.; Versées, W. Crystal structures of the tRNA:m2G6 methyltransferase Trm14/TrmN from two domains of life. Nucleic Acids Res. 2012, 40, 5149–5161. [Google Scholar] [CrossRef] [Green Version]
- Neumann, P.; Lakomek, K.; Naumann, P.T.; Erwin, W.M.; Lauhon, C.T.; Ficner, R. Crystal structure of a 4-thiouridine synthetase-RNA complex reveals specificity of tRNA U8 modification. Nucleic Acids Res. 2014, 42, 6673–6685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirata, A.; Nishiyama, S.; Tamura, T.; Yamauchi, A.; Hori, H. Structural and functional analyses of the archaeal tRNA m2G/m22G10 methyltransferase aTrm11 provide mechanistic insights into site specificity of a tRNA methyltransferase that contains common RNA-binding modules. Nucleic Acids Res. 2016, 44, 6377–6390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, K.; Blaby, I.K.; Thiaville, P.C.; Majumder, M.; Grosjean, H.; Yuan, Y.A.; Gupta, R.; de Crécy-Lagard, V. The archaeal COG1901/DUF358 SPOUT-methyltransferase members, together with pseudouridine synthase Pus10, catalyze the formation of 1-methylpseudouridine at position 54 of tRNA. RNA 2012, 18, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Wurm, J.P.; Griese, M.; Bahr, U.; Held, M.; Heckel, A.; Karas, M.; Soppa, J.; Wöhnert, J. Identification of the enzyme responsible for N1-methylation of pseudouridine 54 in archaeal tRNAs. RNA 2012, 18, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.Y.; Yuan, Y.A. Crystal structure of Mj1640/DUF358 protein reveals a putative SPOUT-class RNA methyltransferase. J. Mol. Cell Biol. 2010, 2, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Forterre, P.; Gribaldo, S.; Brochier-Armanet, C. Happy together: Genomic insights into the unique Nanoarchaeum/Ignicoccus association. J. Biol. 2009, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Kuchino, Y.; Ihara, M.; Yabusaki, Y.; Nishimura, S. Initiator tRNAs from archaebacteria show common unique sequence characteristics. Nature 1982, 298, 684–685. [Google Scholar] [CrossRef]
- Tang, T.H.; Polacek, N.; Zywicki, M.; Huber, H.; Brugger, K.; Garrett, R.; Bachellerie, J.P.; Hüttenhofer, A. Identification of novel non-coding RNAs as potential antisense regulators in the archaeon Sulfolobus solfataricus. Mol. Microbiol. 2005, 55, 469–481. [Google Scholar] [CrossRef]
- Zago, M.A.; Dennis, P.P.; Omer, A.D. The expanding world of small RNAs in the hyperthermophilic archaeon Sulfolobus solfataricus. Mol. Microbiol. 2005, 55, 1812–1828. [Google Scholar] [CrossRef]
- Nishikura, K.; De Robertis, E.M. RNA processing in microinjected Xenopus oocytes. Sequential addition of base modifications in the spliced transfer RNA. J. Mol. Biol. 1981, 145, 405–420. [Google Scholar] [CrossRef]
- Koski, R.A.; Clarkson, S.G. Synthesis and maturation of Xenopus laevis methionine tRNA gene transcripts in homologous cell-free extracts. J. Biol Chem. 1982, 257, 4514–4521. [Google Scholar] [CrossRef]
- Jiang, H.Q.; Motorin, Y.; Jin, Y.X.; Grosjean, H. Pleiotropic effects of intron removal on base modification pattern of yeast tRNAPhe: An in vitro study. Nucleic Acids Res. 1997, 25, 2694–2701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barraud, P.; Gato, A.; Heiss, M.; Catala, M.; Kellner, S.; Tisné, C. Time-resolved NMR monitoring of tRNA maturation. Nat. Commun. 2019, 10, 3373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, K.; Kunibayashi, T.; Tomikawa, C.; Ochi, A.; Kanai, T.; Hirata, A.; Iwashita, C.; Hori, H. Pseudouridine at position 55 in tRNA controls the contents of other modified nucleotides for low-temperature adaptation in the extreme-thermophilic eubacterium Thermus thermophilus. Nucleic Acids Res. 2011, 39, 2304–2318. [Google Scholar] [CrossRef] [Green Version]
- Schultz, S.K.; Kothe, U. tRNA elbow modifications affect the tRNA pseudouridine synthase TruB and the methyltransferase TrmA. RNA 2020, 26, 1131–1142. [Google Scholar] [CrossRef]
- Barraud, P.; Tisné, C. To be or not to be modified: Miscellaneous aspects influencing nucleotide modifications in tRNAs. IUBMB Life 2019, 71, 1126–1140. [Google Scholar] [CrossRef]
- Hori, H. Regulatory Factors for tRNA Modifications in Extreme- Thermophilic Bacterium Thermus thermophilus. Front. Genet. 2019, 10, 204. [Google Scholar] [CrossRef] [Green Version]
- Sokołowski, M.; Klassen, R.; Bruch, A.; Schaffrath, R.; Glatt, S. Cooperativity between different tRNA modifications and their modification pathways. Biochim. Biophys. Acta Gene Regul. Mech. 2018, 1861, 409–418. [Google Scholar] [CrossRef]
- Han, L.; Phizicky, E.M. A rationale for tRNA modification circuits in the anticodon loop. RNA 2018, 24, 1277–1284. [Google Scholar] [CrossRef] [Green Version]
- Perret, V.; Garcia, A.; Puglisi, J.; Grosjean, H.; Ebel, J.P.; Florentz, C.; Giegé, R. Conformation in solution of yeast tRNA(Asp) transcripts deprived of modified nucleotides. Biochimie 1990, 72, 735–743. [Google Scholar] [CrossRef]
- Byrne, R.T.; Konevega, A.L.; Rodnina, M.V.; Antson, A.A. The crystal structure of unmodified tRNAPhe from Escherichia coli. Nucleic Acids Res. 2010, 38, 4154–4162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaswamy, U.; Fox, G.E. Frequent occurrence of the T-loop RNA folding motif in ribosomal RNAs. RNA 2002, 8, 1112–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fechter, P.; Rudinger-Thirion, J.; Florentz, C.; Giegé, R. Novel features in the tRNA-like world of plant viral RNAs. Cell Mol. Life Sci. 2001, 58, 1547–1561. [Google Scholar] [CrossRef] [PubMed]
- Barends, S.; Björk, K.; Gultyaev, A.P.; de Smit, M.H.; Pleij, C.W.; Kraal, B. Functional evidence for D- and T-loop interactions in tmRNA. FEBS Lett. 2002, 514, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Becker, H.F.; Motorin, Y.; Florentz, C.; Giegé, R.; Grosjean, H. Pseudouridine and ribothymidine formation in the tRNA-like domain of turnip yellow mosaic virus RNA. Nucleic Acids Res. 1998, 26, 3991–3997. [Google Scholar] [CrossRef] [Green Version]
- Ranaei-Siadat, E.; Fabret, C.; Seijo, B.; Dardel, F.; Grosjean, H.; Nonin-Lecomte, S. RNA-methyltransferase TrmA is a dual-specific enzyme responsible for C5-methylation of uridine in both tmRNA and tRNA. RNA Biol. 2013, 10, 572–578. [Google Scholar] [CrossRef] [Green Version]
- Zagryadskaya, E.I.; Kotlova, N.; Steinberg, S.V. Key elements in maintenance of the tRNA L-shape. J. Mol. Biol. 2004, 340, 435–444. [Google Scholar] [CrossRef]
- Zagryadskaya, E.I.; Doyon, F.R.; Steinberg, S.V. Importance of the reverse Hoogsteen base pair 54-58 for tRNA function. Nucleic Acids Res. 2003, 31, 3946–3953. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, C.D. RNA versatility governs tRNA function: Why tRNA flexibility is essential beyond the translation cycle. Bioessays 2016, 38, 465–473. [Google Scholar] [CrossRef]
- Levy, M.; Miller, S.L. The prebiotic synthesis of modified purines and their potential role in the RNA world. J. Mol. Evol. 1999, 48, 631–637. [Google Scholar] [CrossRef]
- Cermakian, N.; Cedergren, R. Modified Nucleosides Always Were: An Evolutionary Model. In Modification and Editing of RNA; Grosjean, H., Benne, R., Eds.; ASM Press: Washington, DC, USA, 1998; pp. 535–541. [Google Scholar]
- McKenney, K.M.; Alfonzo, J.D. From Prebiotics to Probiotics: The Evolution and Functions of tRNA Modifications. Life Basel 2016, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anantharaman, V.; Koonin, E.V.; Aravind, L. Comparative genomics and evolution of proteins involved in RNA metabolism. Nucleic Acids Res. 2002, 30, 1427–1464. [Google Scholar] [CrossRef] [PubMed]
- McKenney, K.M.; Rubio, M.A.T.; Alfonzo, J.D. The Evolution of Substrate Specificity by tRNA Modification Enzymes. Enzymes 2017, 41, 51–88. [Google Scholar] [CrossRef] [PubMed]
- Galperin, M.Y.; Walker, D.R.; Koonin, E.V. Analogous enzymes: Independent inventions in enzyme evolution. Genome Res. 1998, 8, 779–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soucy, S.M.; Huang, J.; Gogarten, J.P. Horizontal gene transfer: Building the web of life. Nat. Rev. Genet. 2015, 16, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Sieber, K.B.; Bromley, R.E.; Dunning Hotopp, J.C. Lateral gene transfer between prokaryotes and eukaryotes. Exp. Cell Res. 2017, 358, 421–426. [Google Scholar] [CrossRef]
- Galperin, M.Y.; Koonin, E.V. Sources of systematic error in functional annotation of genomes: Domain rearrangement, non-orthologous gene displacement and operon disruption. Silico Biol. 1998, 1, 55–67. [Google Scholar]
- Zallot, R.; Harrison, K.J.; Kolaczkowski, B.; de Crécy-Lagard, V. Functional Annotations of Paralogs: A Blessing and a Curse. Life 2016, 6, 39. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roovers, M.; Droogmans, L.; Grosjean, H. Post-Transcriptional Modifications of Conserved Nucleotides in the T-Loop of tRNA: A Tale of Functional Convergent Evolution. Genes 2021, 12, 140. https://doi.org/10.3390/genes12020140
Roovers M, Droogmans L, Grosjean H. Post-Transcriptional Modifications of Conserved Nucleotides in the T-Loop of tRNA: A Tale of Functional Convergent Evolution. Genes. 2021; 12(2):140. https://doi.org/10.3390/genes12020140
Chicago/Turabian StyleRoovers, Martine, Louis Droogmans, and Henri Grosjean. 2021. "Post-Transcriptional Modifications of Conserved Nucleotides in the T-Loop of tRNA: A Tale of Functional Convergent Evolution" Genes 12, no. 2: 140. https://doi.org/10.3390/genes12020140
APA StyleRoovers, M., Droogmans, L., & Grosjean, H. (2021). Post-Transcriptional Modifications of Conserved Nucleotides in the T-Loop of tRNA: A Tale of Functional Convergent Evolution. Genes, 12(2), 140. https://doi.org/10.3390/genes12020140