
Chromosomal Polymorphism and Speciation: The Case of the Genus Mazama (Cetartiodactyla; Cervidae)

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Species and Samples
2.2. Whole-Chromosome Painting and Bacterial Artificial Chromosomes (BAC) Probes
2.3. FISH
2.4. Inter-Specific Hybrids (MGO × MNE) Reproductive Assessment
2.4.1. Spermiogram
2.4.2. Testicular Histology
2.5. Intra-Specific Hybrids (MAM) Reproductive Assessment
2.5.1. Semen Samples and Sperm Nuclei Preparation
2.5.2. Sperm-FISH
2.6. Statistical Analysis
3. Results
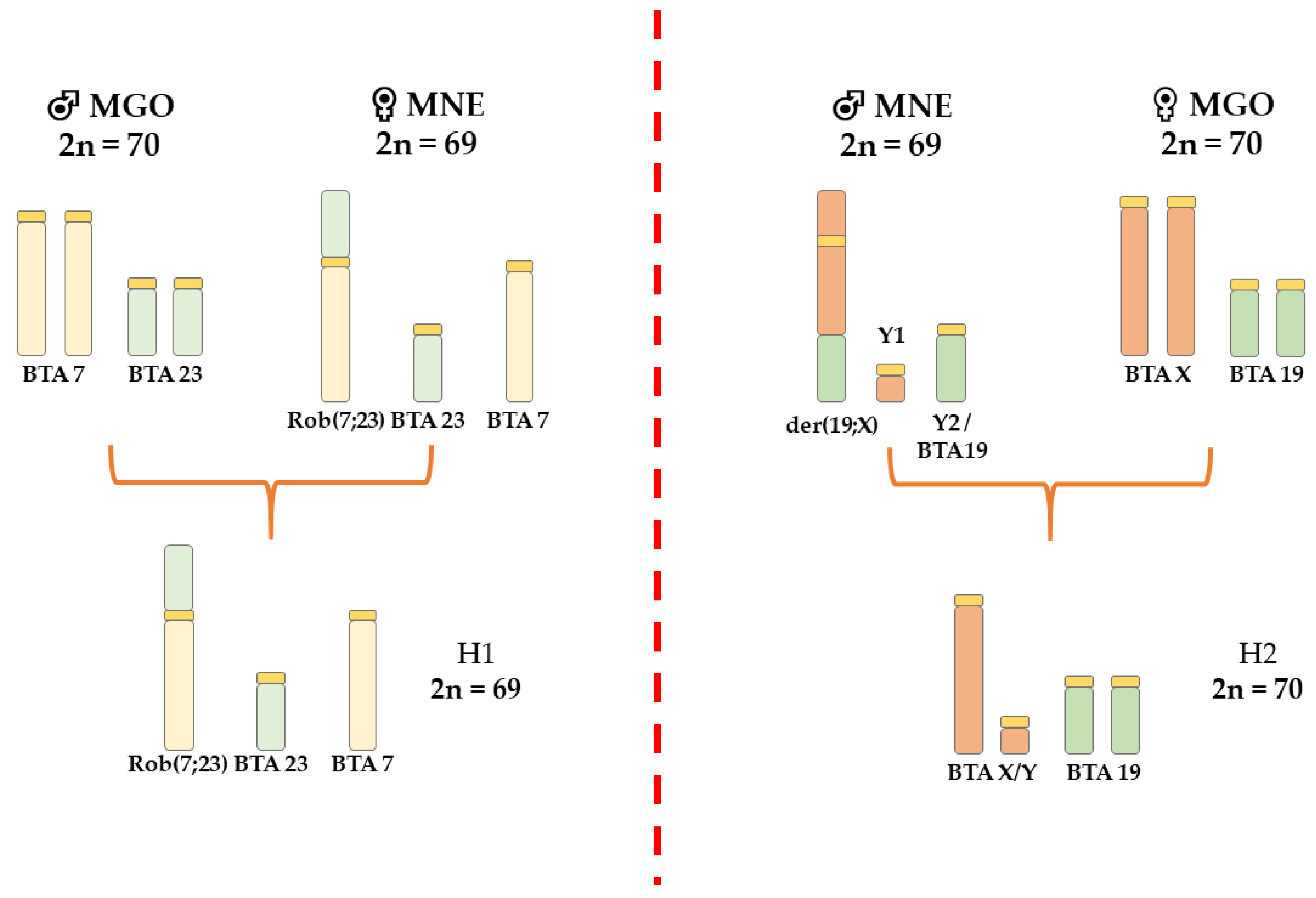
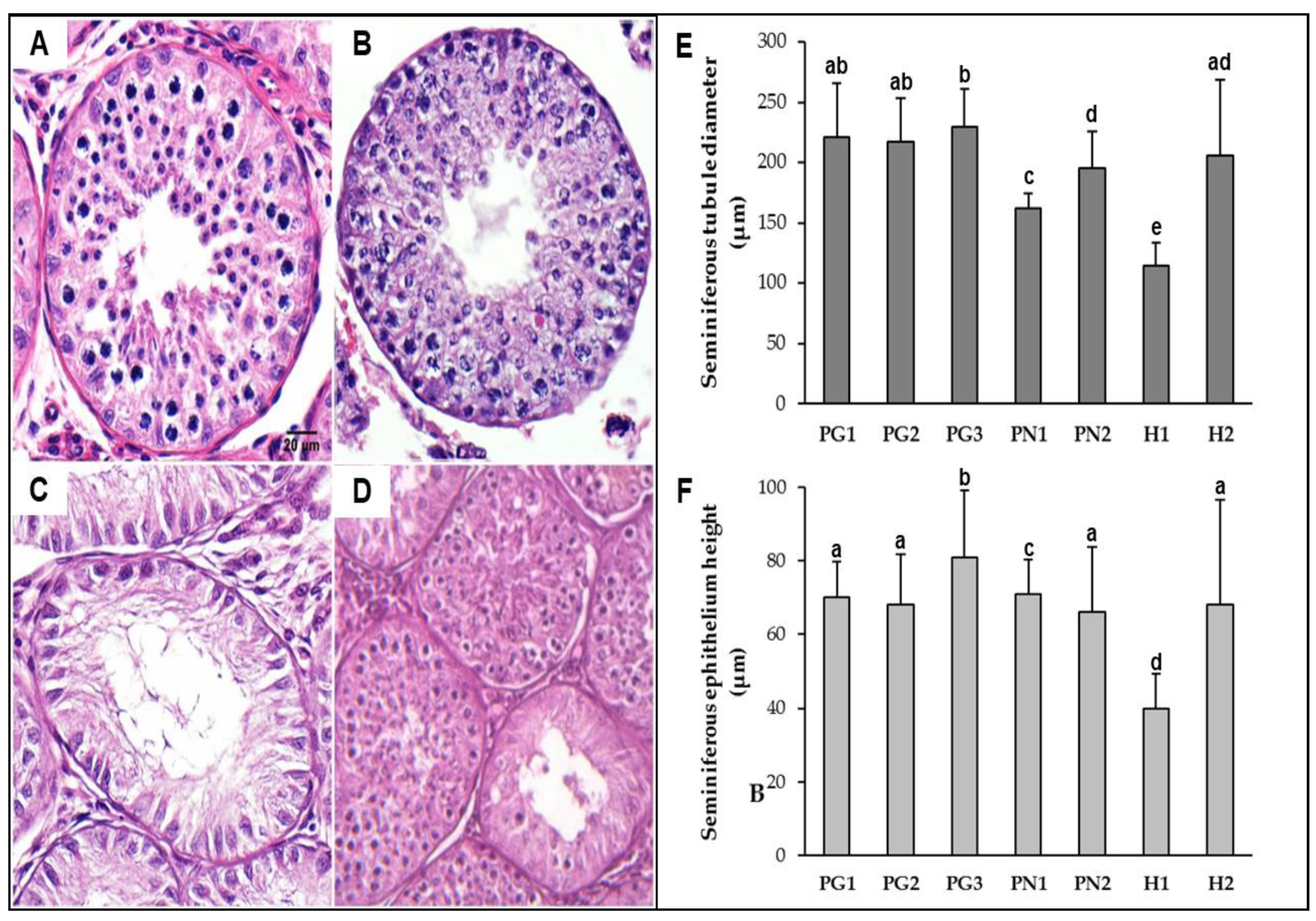
3.1. Inter-Specific Hybrids
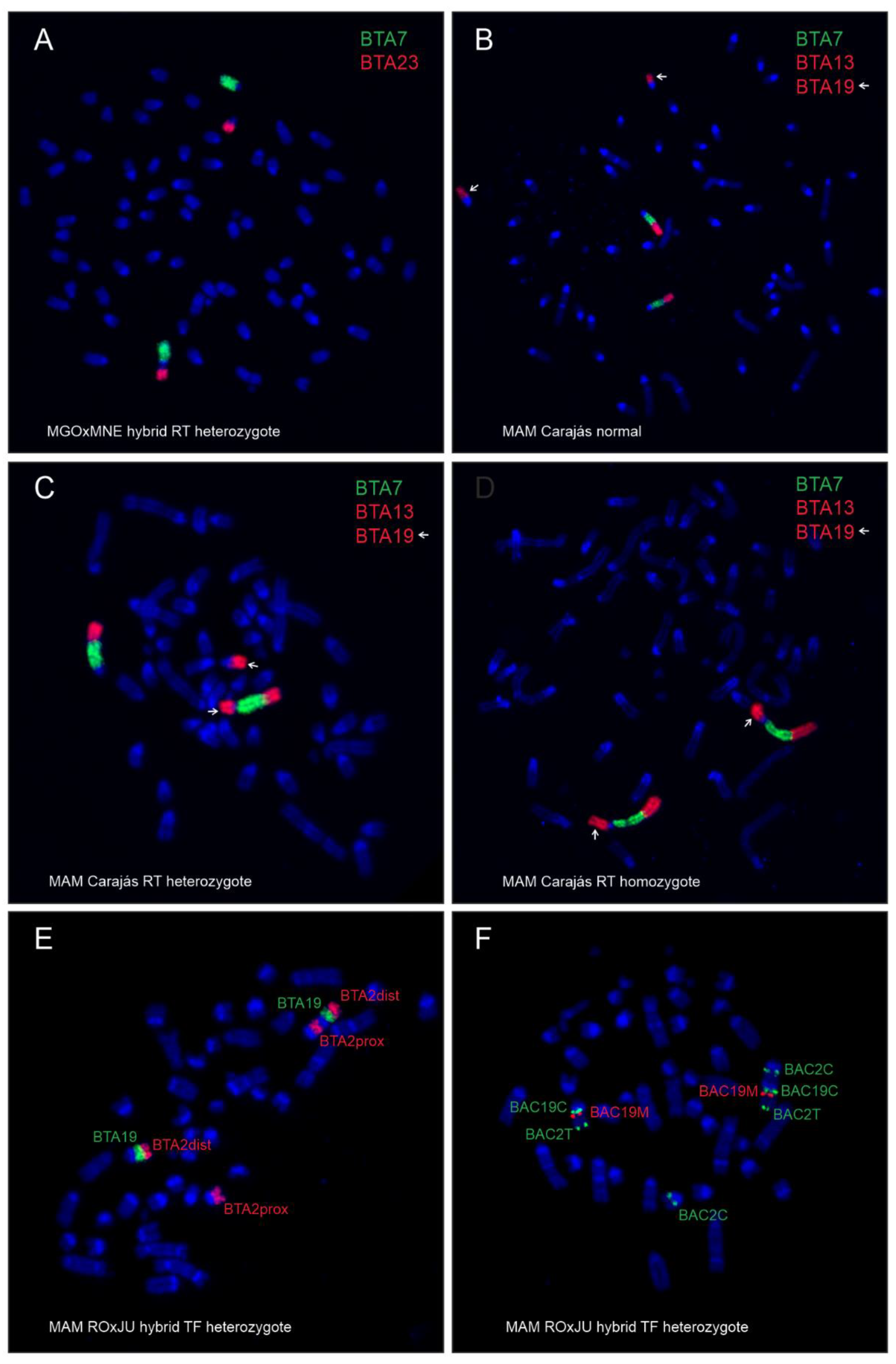
3.2. Intra-Specific Hybrids
4. Discussion
4.1. Inter-Specific Hybrids
4.2. Intra-Specific Hybrids
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobigny, G.; Britton-Davidian, J.; Robinson, T.J. Chromosomal polymorphism in mammals: An evolutionary perspective. Biol. Rev. Camb. Philos. Soc. 2017, 92, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Faria, R.; Navarro, A. Chromosomal speciation revisited: Rearranging theory with pieces of evidence. Trends Ecol. Evol. 2010, 25, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Farré, M.; Micheletti, D.; Ruiz-Herrera, A. Recombination rates and genomic shuffling in human and chimpanzee--a new twist in the chromosomal speciation theory. Mol. Biol. Evol. 2013, 30, 853–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; O’Brien, P.C.M.; Wienberg, J.; Ferguson-Smith, M.A. A reappraisal of the tandem fusion theory of karyotype evolution in Indian muntjac using chromosome painting. Chromosome Res. 1997, 5, 109–117. [Google Scholar] [CrossRef]
- Yang, F.; O’Brien, P.C.M.; Wienberg, J.; Neitzel, H.; Lin, C.C.; Ferguson-Smith, M.A. Chromosomal evolution of the Chinese muntjac (Muntiacus reevesi). Chromosoma 1997, 106, 37–43. [Google Scholar] [CrossRef]
- Duarte, J.M.B.; Jorge, W. Morphologic and Cytogenetic Description of the Small Red Brocket (Mazama bororo Duarte,1996) in Brazil. Mammalia 2003, 67, 403–410. [Google Scholar] [CrossRef]
- Neitzel, H. Chromosome Evolution of Cervidae: Karyotypic and Molecular Aspects. In Cytogenetics—Basic and Applied Aspects; Obe, G., Basler, A., Eds.; Springer: Berlin/Heidelberg, Germany, 1987; pp. 90–112. ISBN 978-3-642-728004-4. [Google Scholar]
- Fontana, F.; Rubini, M. Chromosomal evolution in cervidae. BioSystems 1990, 24, 157–174. [Google Scholar] [CrossRef]
- Abril, V.V.; Duarte, J.M.B. Chromosome polymorphism in the Brazilian dwarf brocket deer, Mazama nana (Mammalia, Cervidae). Genet. Mol. Biol. 2008, 31, 53–57. [Google Scholar] [CrossRef] [Green Version]
- Abril, V.V.; Carnelossi, E.A.G.; González, S.; Duarte, J.M.B. Elucidating the Evolution of the Red Brocket Deer Mazama americana Complex (Artiodactyla; Cervidae). Cytogenet. Genome Res. 2010, 128, 177–187. [Google Scholar] [CrossRef]
- Fiorillo, B.F.; Sarria-Perea, J.A.; Abril, V.V.; Duarte, J.M.B. Cytogenetic description of the Amazonian brown brocket Mazama nemorivaga (Artiodactyla, Cervidae). Comp. Cytogenet. 2013, 7, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Valeri, M.P.; Tomazella, I.M.; Duarte, J.M.B. Intrapopulation Chromosomal Polymorphism in Mazama gouazoubira (Cetartiodactyla; Cervidae): The Emergence of a New Species? Cytogenet. Genome Res. 2018, 154, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Tomazella, I.M. Análise de Polimorfismo Cromossômico em Mazama Gouazoubira (Artiodactyla; Cervidae): Implicações Para a Evolução Cariotípica em Cervidae; Universidade Estadual Paulista: Ilha Solteira, Brazil, 2016. [Google Scholar]
- Vargas-Munar, D.S.F.; Sarria-Perea, J.A.; Duarte, J.M.B. Different responses to doxorubicin-induced chromosome aberrations in Brazilian deer species. Genet. Mol. Res. 2010, 9, 1545–1549. [Google Scholar] [CrossRef] [PubMed]
- Tomazella, I.M.; Abril, V.V.; Duarte, J.M.B. Identifying Mazama gouazoubira (Artiodactyla; Cervidae) chromosomes involved in rearrangements induced by doxorubicin. Genet. Mol. Biol. 2017, 40, 460–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, J.M.B.; González, S.; Maldonado, J.E. The surprising evolutionary history of South American deer. Mol. Phylogenet. Evol. 2008, 49, 17–22. [Google Scholar] [CrossRef] [PubMed]
- González, S.; Barbanti Duarte, J.M. Speciation, evolutionary history and conservation trends of Neotropical deer. Mastozoología Neotrop. 2020, 27, 35–46. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2020-2. Available online: https://www.iucnredlist.org/ (accessed on 24 July 2020).
- Rossi, R.V. Taxonomia de Mazama RAFINESQUE, 1817, do Brasil (Artiodactyla, Cervidae); Universidade de São Paulo: São Paulo, Brazil, 2000. [Google Scholar]
- Morales-Donoso, J.A. Caracterização Morfológica, Citogenética e Molecular de Mazama Nemorivaga (Cuvier, 1817) a Partir de um Topótipo Atual; Universidade Estadual Paulista: Ilha Solteira, Brazil, 2017. [Google Scholar]
- Duarte, J.M.B.; Merino, M.L. Taxonomia e Evolução. In Biologia e Conservação de Cervídeos Sul—Americanos: Blastocerus, Ozotoceros e Mazama; Duarte, J.M.B., Ed.; FUNEP: Jaboticabal, Brazil, 1997; pp. 1–21. [Google Scholar]
- Figueiredo, M.G. Filogenia e Taxonomia dos Veados Cinza (Mazama gouazoubira e M. nemorivaga); Universidade Estadual Paulista: Ilha Solteira, Brazil, 2014. [Google Scholar]
- Resende, J.P.d.A. Comparação Cariotípica Entre Mazama Gouazoubira e Mazama Nemorivaga (Artiodactyla; Cervidae) por Meio de Marcadores Citogenéticos Clássicos, Fish Telomérica e Pintura Cromossômica; Universidade Estadual Paulista: Ilha Solteira, Brazil, 2012. [Google Scholar]
- King, M. Species Evolution The Role of Chromosome Change; Cambridge University Press: Cambridge, UK, 1993; ISBN 9780521484541. [Google Scholar]
- Salviano, M.B.; Cursino, M.S.; Zanetti, E.S.; Abril, V.V.; Duarte, J.M.B. Intraspecific chromosome polymorphisms can lead to reproductive isolation and speciation: An example in red brocket deer (Mazama americana). Biol. Reprod. 2017, 96, 1279–1287. [Google Scholar] [CrossRef]
- Carranza, J.; Roldán, M.; Duarte, J.M.B. Lack of mate selectivity for genetic compatibility within the red brocket deer Mazama americana complex. Mamm. Biol. 2018, 88, 168–175. [Google Scholar] [CrossRef]
- Aquino, C.I.; Abril, V.V.; Duarte, J.M.B. Meiotic pairing of B chromosomes, multiple sexual system, and Robertsonian fusion in the red brocket deer Mazama americana (Mammalia, Cervidae). Genet. Mol. Res. 2013, 12, 3566–3574. [Google Scholar] [CrossRef]
- Abril, V.V. Evolução Cromossômica no Veado-Mateiro—Mazama Americana (Mammalia; Cervidae); Universidade Estadual Paulista: Ilha Solteira, Brazil, 2009. [Google Scholar]
- Fröhlich, J.; Kubickova, S.; Musilova, P.; Cernohorska, H.; Muskova, H.; Rubes, J. A Comparative Study of Pygmy Hippopotamus (Choeropsis liberiensis) Karyotype by Cross-Species Chromosome Painting. J. Mamm. Evol. 2017, 24, 465–474. [Google Scholar] [CrossRef]
- Kubickova, S.; Cernohorska, H.; Musilova, P.; Rubes, J. The use of laser microdissection for the preparation of chromosome-specific painting probes in farm animals. Chromosome Res. 2002, 10, 571–577. [Google Scholar] [CrossRef]
- Telenius, H.; Carter, N.P.; Bebb, C.E.; Nordenskjöld, M.; Ponder, B.A.J.; Tunnacliffe, A. Degenerate oligonucleotide-primed PCR: General amplification of target DNA by a single degenerate primer. Genomics 1992, 13, 718–725. [Google Scholar] [CrossRef]
- Vozdova, M.; Kubickova, S.; Cernohorska, H.; Fröhlich, J.; Vodicka, R.; Rubes, J. Comparative Study of the Bush Dog (Speothos venaticus) Karyotype and Analysis of Satellite DNA Sequences and Their Chromosome Distribution in Six Species of Canidae. Cytogenet. Genome Res. 2019, 159, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Favoretto, S.M.; Zanetti, E.S.; Duarte, J.M.B. Cryopreservation of red brocket deer semen (Mazama americana): Comparison between three extenders. J. Zoo Wildl. Med. 2012, 43, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Duarte, J.M.B.; Garcia, J.M. Assistant Reproduction in Brazilian Cervidae. Rev. Bras. Reprod. Anim. 1995, 19, 111–121. [Google Scholar]
- Alvarez, M.C.L.; Rola, L.D.; Duarte, J.M.B. Comparison Between Three Cryoprotectants in the Freezing of Mazama americana Semen Collected by Artificial Vagina. Biopreserv. Biobank. 2020. [Google Scholar] [CrossRef] [PubMed]
- BLOM, E. Interpretation of spermatic cytology in bulls. Fertil. Steril. 1950, 1, 223–238. [Google Scholar] [CrossRef]
- Rubes, J.; Vozdová, M.; Kubícková, S. Aneuploidy in pig sperm: Multicolor fluorescence in situ hybridization using probes for chromosomes 1, 10, and Y. Cytogenet. Cell Genet. 1999, 85, 200–204. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Lenguage and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Cursino, M.S.; Duarte, J.M.B. Using sperm morphometry and multivariate analysis to differentiate species of gray Mazama. R. Soc. Open Sci. 2016, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Vozdova, M.; Sebestova, H.; Kubickova, S.; Cernohorska, H.; Awadova, T.; Vahala, J.; Rubes, J. Impact of Robertsonian translocation on meiosis and reproduction: An impala (Aepyceros melampus) model. J. Appl. Genet. 2014, 55, 249–258. [Google Scholar] [CrossRef]
- Pinton, A.; Faraut, T.; Yerle, M.; Gruand, J.; Pellestor, F.; Ducos, A. Comparison of male and female meiotic segregation patterns in translocation heterozygotes: A case study in an animal model (Sus scrofa domestica L.). Hum. Reprod. 2005, 20, 2476–2482. [Google Scholar] [CrossRef] [Green Version]
- Bonnet-Garnier, A.; Lacaze, S.; Beckers, J.F.; Berland, H.M.; Pinton, A.; Yerle, M.; Ducos, A. Meiotic segregation analysis in cows carrying the t(1;29) Robertsonian translocation. Cytogenet. Genome Res. 2008, 120, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Massip, K.; Yerle, M.; Billon, Y.; Ferchaud, S.; Bonnet, N.; Calgaro, A.; Mary, N.; Dudez, A.-M.; Sentenac, C.; Plard, C.; et al. Studies of male and female meiosis in inv(4)(p1.4;q2.3) pig carriers. Chromosom. Res. 2010, 18, 925–938. [Google Scholar] [CrossRef] [PubMed]
- Zackowski, J.L.; Martin-DeLeon, P.A. Segregation products of male mice doubly heterozygous for the RB(6.16) and RB(16.17) translocations: Influence of sperm karyotype on fertilizing competence under varying mating frequencies. Gamete Res. 1989, 22, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Sánchez, R.; Gómez-Fidalgo, E.; Pérez-Garnelo, S.; Martín-Lluch, M.; De la Cruz-Vigo, P. Prevalence of chromosomal aberrations in breeding pigs in Spain. Reprod. Domest. Anim. 2019, 54, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Carter, N.P.; Shi, L.; Ferguson-Smith, M.A. A comparative study of karyotypes of muntjacs by chromosome painting. Chromosoma 1995, 103, 642–652. [Google Scholar] [CrossRef]
- Duarte, J.M.B.; Jorge, W. Chromosomal polymorphism in several populations of deer (Genus Mazama) from Brazil. Arch. Zootec. 1996, 45, 281–287. [Google Scholar]
- Grize, S.A.; Wilwert, E.; Searle, J.B.; Lindholm, A.K. Measurements of hybrid fertility and a test of mate preference for two house mouse races with massive chromosomal divergence. BMC Evol. Biol. 2019, 19, 25. [Google Scholar] [CrossRef]
- Oka, A.; Mita, A.; Takada, Y.; Koseki, H.; Shiroishi, T. Reproductive isolation in hybrid mice due to spermatogenesis defects at three meiotic stages. Genetics 2010, 186, 339–351. [Google Scholar] [CrossRef] [Green Version]
- Li, X.C.; Barringer, B.C.; Barbash, D.A. The pachytene checkpoint and its relationship to evolutionary patterns of polyploidization and hybrid sterility. Heredity 2009, 102, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Switoński, M.; Stranzinger, G. Studies of synaptonemal complexes in farm mammals—A review. J. Hered. 1998, 89, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Barrozo, L.A.; Toniolo, G.H.; Duarte, J.M.B.; Pinho, M.P.; Oliveira, J.A. Padrão anual de variação da testosterona sérica, volume testicular e aspectos seminais de veados-catingueiros (Mazama gouazoubira) em cativeiro. Rev. Bras. Reprod. Anim. 2001, 25, 210–211. [Google Scholar]
- Benirschke, K. Sterility and Fertility of Interspecific Mammalian Hybrids. In Comparative Aspects of Reproductive Failure; Springer: Berlin/Heidelberg, Germany, 1967; pp. 218–234. ISBN 978-3-642-48949-5. [Google Scholar]
- Ashley, T. An integration of old and new perspectives of mammalian meiotic sterility. Results Probl. Cell Differ. 2000, 28, 131–173. [Google Scholar] [CrossRef] [PubMed]
- Wishart, W.D.; Hrudka, F.; Schmutz, S.M.; Flood, P.F. Observations on spermatogenesis, sperm phenotype, and fertility in white-tailed × mule deer hybrids and a yak × cow hybrid. Can. J. Zool. 1988, 66, 1664–1671. [Google Scholar] [CrossRef]
- McEntee, K. Reproductive Pathology of Domestic Mammals; Academic Press: San Diego, CA, USA, 1990; ISBN 9780124833753. [Google Scholar]
- Britton-Davidian, J.; Fel-Clair, F.; Lopez, J.; Alibert, P.; Boursot, P. Postzygotic isolation between the two European subspecies of the house mouse: Estimates from fertility patterns in wild and laboratory-bred hybrids. Biol. J. Linn. Soc. 2005, 84, 379–393. [Google Scholar] [CrossRef] [Green Version]
- McGovern, P.T. The barriers to interspecific hybridization in domestic and laboratory mammals. II. Hybrid sterility. Br. Vet. J. 1976, 132, 68–75. [Google Scholar] [CrossRef]
- Jadwiszczak, K.A.; Banaszek, A. Fertility in the male common shrews, Sorex araneus, from the extremely narrow hybrid zone between chromosome races. Mamm. Biol. 2006, 71, 257–267. [Google Scholar] [CrossRef]
- Trujillo, J.M.; Ohno, S.; Jardine, J.H.; Atkins, N.B. Spermatogenesis in a male hinny: Histological and cytological studies. J. Hered. 1969, 60, 79–84. [Google Scholar] [CrossRef]
- Chandley, A.C.; Jones, R.C.; Dott, H.M.; Allen, W.R.; Short, R.V. Meiosis in interspecific equine hybrids. I. The male mule (Equus asinus X E. caballus) and hinny (E. caballus X E. asinus). Cytogenet. Cell Genet. 1974, 13, 330–341. [Google Scholar] [CrossRef]
- Gray, A.P. Mammalian Hybrids. A Check-List with Bibliography; Commonwealth Agricultural Bureaux International: Buckinghamshire, UK, 1954. [Google Scholar]
- Carranza, J.; Roldán, M.; de Fátima Carvalho Peroni, E.; Duarte, J.M.B. Weak premating isolation between two parapatric brocket deer species. Mamm. Biol. 2017, 87, 17–26. [Google Scholar] [CrossRef]
- Bonnet-Garnier, A.; Pinton, A.; Berland, H.M.; Khireddine, B.; Eggen, A.; Yerle, M.; Darré, R.; Ducos, A. Sperm nuclei analysis of 1/29 Robertsonian translocation carrier bulls using fluorescence in situ hybridization. Cytogenet. Genome Res. 2006, 112, 241–247. [Google Scholar] [CrossRef]
- Barasc, H.; Mouney-Bonnet, N.; Peigney, C.; Calgaro, A.; Revel, C.; Mary, N.; Ducos, A.; Pinton, A. Analysis of Meiotic Segregation Pattern and Interchromosomal Effects in a Bull Heterozygous for a 3/16 Robertsonian Translocation. Cytogenet. Genome Res. 2018, 156, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Pinton, A.; Calgaro, A.; Bonnet, N.; Ferchaud, S.; Billoux, S.; Dudez, A.M.; Mary, N.; Massip, K.; Bonnet-Garnier, A.; Yerle, M.; et al. Influence of sex on the meiotic segregation of a t(13;17) Robertsonian translocation: A case study in the pig. Hum. Reprod. 2009, 24, 2034–2043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manieu, C.; González, M.; López-Fenner, J.; Page, J.; Ayarza, E.; Fernández-Donoso, R.; Berríos, S. Aneuploidy in spermatids of Robertsonian (Rb) chromosome heterozygous mice. Chromosome Res. 2014, 22, 545–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiland, E.; Olszewska, M.; Woźniak, T.; Kurpisz, M. How much, if anything, do we know about sperm chromosomes of Robertsonian translocation carriers? Cell. Mol. Life Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Switoński, M.; Gustavsson, I.; Plöen, L. The nature of the 1;29 translocation in cattle as revealed by synaptonemal complex analysis using electron microscopy. Cytogenet. Cell Genet. 1987, 44, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Kingswood, S.C.; Kumamoto, A.T.; Sudman, P.D.; Fletcher, K.C.; Greenbaum, I.F. Meiosis in chromosomally heteromorphic goitered gazelle, Gazella subgutturosa (Artiodactyla, Bovidae). Chromosome Res. 1994, 2, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Moritz, C. The Population Biology of Gehyra (Gekkonidae): Chromosome Change and Speciation. Syst. Biol. 1986, 35, 46–67. [Google Scholar] [CrossRef]
- Long, S.E. Tandem 1;30 translocation: A new structural abnormality in the horse (Equus caballus). Cytogenet. Genome Res. 1996, 72, 162–163. [Google Scholar] [CrossRef]
- Pillay, N.; Willan, K.; Meester, J. Post-zygotic reproductive isolation in two populations of the African vlei rat Otomys irroratus. Acta Theriol. 1995, 40, 69–76. [Google Scholar] [CrossRef] [Green Version]
- White, M.; Blackith, R.; Blackith, R.; Cheney, J. Cytogenetics of the viatica group morabine grasshoppers. I. The coastal species. Aust. J. Zool. 1967, 15, 263. [Google Scholar] [CrossRef]
- Cursino, M.S.; Salviano, M.B.; Abril, V.V.; dos Santos Zanetti, E.; Duarte, J.M.B. The role of chromosome variation in the speciation of the red brocket deer complex: The study of reproductive isolation in females. BMC Evol. Biol. 2014, 14, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Animal | Species | 2n | FN | Translocations | B | |
|---|---|---|---|---|---|---|
| RT | Multiple Sexual System | |||||
| PG1 | M. gouazoubira | 70 | 70 | - | No | 0–2 |
| PG2 | M. gouazoubira | 70 | 70 | - | No | 0–2 |
| PG3 | M. gouazoubira | 70 | 70 | - | No | 0–2 |
| PN1 | M. nemorivaga | 68 | 72 | rob(4;32)(4;32) a | No | 1–9 |
| PN2 | M. nemorivaga | 67 | 70 | rob(4;32)(4;32) a | Yes a | 2–5 |
| H1 | MGO  × MNE × MNE  | 69 | 72 | rob(4;32) a | No | 0–3 |
| H2 | MNE × MGO | 70 | 70 | - | No | 0–2 |
| Animal | Cytotypes | 2n | FN | Translocations | B | |
|---|---|---|---|---|---|---|
| RT | TF | |||||
| T297 a | Carajás | 51 | 54 | - | - | 2–3 |
| T274 | Carajás | 50 | 54 | rob(5;11) b | - | 3 |
| T326 | Carajás | 49 | 54 | rob(5;11)(5;11) b | - | 3–4 |
| T269 | Rondônia | 42 | 46 | rob(7;20) c | - | 3–5 |
| T343 | Juína × Rondônia | 43 | 47 | rob(7;20) c | der(7;10) d | 2–3 |
| T347 | Rondônia × Juína | 44 | 47 | - | der(7;10) d | 2–4 |
| T421 | Paraná × Carajás | 52 | 55 | - | der(5;10) e | - |
| Animal | Spermatogonia A (%) | Spermatogonia B (%) | Leptotenes/Zygotenes (%) | Pachytenes (%) | Round Spermatids (%) | Sertoli Cells (%) | SSR a |
|---|---|---|---|---|---|---|---|
| PG1 | 17.36 ± 12.70 | 22.63 ± 10.49 | 9.61 ± 4.30 | 10.53 ± 2.75 | 21.45 ± 12.34 | 18.42 ± 2.91 | 2.03 |
| PG2 | 6.34 ± 2.20 | 12.67 ± 7.08 | 21.42 ± 22.00 | 16.16 ± 5.81 | 33.40 ± 15.23 | 10.01 ± 2.46 | 2.06 |
| PG3 | 11.03 ± 7.98 | 12.76 ± 7.44 | 16.77 ± 31.65 | 14.54 ± 14.46 | 33.36 ± 23.08 | 11.53 ± 4.00 | 2.29 |
| PN1 | 15.61 ± 4.69 | 13.33 ± 6.81 | 22.28 ± 10.90 | 15.70 ± 4.43 | 23.86 ± 14.72 | 9.22 ± 1.43 | 1.50 |
| PN2 | 9.42 ± 7.04 | 8.22 ± 2.45 | 16.38 ± 13.49 | 10.47 ± 5.72 | 43.95 ± 20.33 | 11.56 ± 4.12 | 4.19 |
| H1 | 41.64 ± 4.55 | 29.79 ± 2.39 | 9.42 ± 0.00 | 0.00 | 0.00 | 19.15 ± 3.65 | 0.00 |
| H2 | 15.52 ± 3.39 | 15.64 ± 6.82 | 22.79 ± 8.36 | 18.18 ± 5.23 | 15.27 ± 6.17 | 12.60 ± 2.01 | 0,84 |
| Animal | Volume (μL) | Concentration (109/mL) | Color | Motility (%) | Vigor (0–5) | Defects (%) | Normal Sperm (%) | |
|---|---|---|---|---|---|---|---|---|
| Primary | Secondary | |||||||
| PG1 | 375 | 0.57 | White | 40 | 2 | 37.0 | 5.0 | 58.0 |
| PG2 | 240 | 3.34 | White | 65 | 4 | 29.5 | 13.5 | 57.0 |
| PG3 | 270 | 2.32 | White | 60 | 3 | 13.5 | 13.0 | 73.0 |
| PN1 | 375 | 2.25 | Reddish a | 70 | 3 | 35.0 | 13.5 | 51.5 |
| PN2 | 60 | 2.71 | Reddish a | 90 | 4 | 4.0 | 39.0 | 57.0 |
| H1 b | 160 | - | Clear | - | - | - | - | - |
| H2 | 50 | 0.02 | Watery | <1 | 0 | 66.0 | 24.0 | 10.0 |
| Robertsonian Translocation (A;B) (%) | ||||||||
|---|---|---|---|---|---|---|---|---|
| FISH Phenotype | T269 rob(7;20) a,d | T274 rob(5;11) a,e | T326 rob(5;11) b,e | T297 c,e | ||||
| Normal/Balanced | 4890 | (97.80) | 4884 | (97.68) | 4949 | (98.98) | 4926 | (98.52) |
| Nullisomy A | 36 | (0.72) | 21 | (0.42) | 18 | (0.36) | 19 | (0.38) |
| Disomy A | 7 | (0.14) | 10 | (0.20) | 6 | (0.12) | 3 | (0.06) |
| Nullisomy B | 25 | (0.50) | 33 | (0.66) | 8 | (0.16) | 17 | (0.34) |
| Disomy B | 29 | (0.58) | 19 | (0.38) | 7 | (0.14) | 7 | (0.14) |
| Total adjacent | 97 | (1.94) | 83 | (1.66) | 39 | (0.78) | 46 | (0.92) |
| Disomy A + B | 1 | (0.02) | 9 | (0.18) | 0 | (0.00) | 7 | (0.14) |
| Nullisomy A + B | 1 | (0.02) | 9 | (0.18) | 6 | (0.12) | 2 | (0.04) |
| Diploidy | 7 | (0.14) | 10 | (0.20) | 5 | (0.10) | 19 | (0.38) |
| Others f | 4 | (0.08) | 5 | (0.10) | 1 | (0.02) | 0 | (0.00) |
| Total unbalanced | 110 | (2.20) | 116 | (2.32) | 51 | (1.02) | 74 | (1.48) |
| TOTAL | 5000 | (100) | 5000 | (100) | 5000 | (100) | 5000 | (100) |
| Tandem Fusion (A;B) (%) | ||||||||
|---|---|---|---|---|---|---|---|---|
| FISH Phenotype | T343 der(7;10) a | T347 der(7;10) a | T421 der(5;10) b | T297 b,c | ||||
| Normal/Balanced | 1139 | (56.95) | 1390 | (69.50) | 1505 | (75.25) | 1970 | (98.50) |
| Nullisomy A | 328 | (16.40) | 203 | (10.15) | 166 | (8.30) | 6 | (0.30) |
| Disomy A | 249 | (12.45) | 170 | (8.50) | 144 | (7.20) | 5 | (0.25) |
| Nullisomy B | 124 | (6.20) | 109 | (5.45) | 56 | (2.80) | 8 | (0.40) |
| Disomy B | 82 | (4.10) | 67 | (3.35) | 46 | (2.30) | 4 | (0.20) |
| Total adjacent | 783 | (39.15) | 549 | (27.45) | 412 | (20.60) | 23 | (1.15) |
| Disomy A + B | 18 | (0.90) | 13 | (0.65) | 26 | (1.30) | 4 | (0.20) |
| Nullisomy A + B | 11 | (0.55) | 12 | (0.60) | 11 | (0.55) | 3 | (0.15) |
| Diploidy | 15 | (0.75) | 11 | (0.55) | 12 | (0.60) | 0 | (0.00) |
| Others d | 34 | (1.70) | 25 | (1.25) | 34 | (1.70) | 0 | (0.00) |
| Total unbalanced | 861 | (43.05) | 610 | (30.50) | 495 | (24.75) | 30 | (1.50) |
| TOTAL | 2000 | (100) | 2000 | (100) | 2000 | (100) | 2000 | (100) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galindo, D.J.; Martins, G.S.; Vozdova, M.; Cernohorska, H.; Kubickova, S.; Bernegossi, A.M.; Kadlcikova, D.; Rubes, J.; Duarte, J.M.B. Chromosomal Polymorphism and Speciation: The Case of the Genus Mazama (Cetartiodactyla; Cervidae). Genes 2021, 12, 165. https://doi.org/10.3390/genes12020165
Galindo DJ, Martins GS, Vozdova M, Cernohorska H, Kubickova S, Bernegossi AM, Kadlcikova D, Rubes J, Duarte JMB. Chromosomal Polymorphism and Speciation: The Case of the Genus Mazama (Cetartiodactyla; Cervidae). Genes. 2021; 12(2):165. https://doi.org/10.3390/genes12020165
Chicago/Turabian StyleGalindo, David Javier, Gabriela Siqueira Martins, Miluse Vozdova, Halina Cernohorska, Svatava Kubickova, Agda Maria Bernegossi, Dita Kadlcikova, Jiri Rubes, and José Maurício Barbanti Duarte. 2021. "Chromosomal Polymorphism and Speciation: The Case of the Genus Mazama (Cetartiodactyla; Cervidae)" Genes 12, no. 2: 165. https://doi.org/10.3390/genes12020165
APA StyleGalindo, D. J., Martins, G. S., Vozdova, M., Cernohorska, H., Kubickova, S., Bernegossi, A. M., Kadlcikova, D., Rubes, J., & Duarte, J. M. B. (2021). Chromosomal Polymorphism and Speciation: The Case of the Genus Mazama (Cetartiodactyla; Cervidae). Genes, 12(2), 165. https://doi.org/10.3390/genes12020165