A Forensic Detection Method for Hallucinogenic Mushrooms via High-Resolution Melting (HRM) Analysis

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Genomic DNA Extraction
2.3. Primer Selection
2.4. Specificity Studies with HRM Analysis
2.5. Reproductivity, Sensitivity, and Mixture Testing
2.6. ITS Sequencing and Data Analysis
2.7. Unknown Sample Detection
3. Results
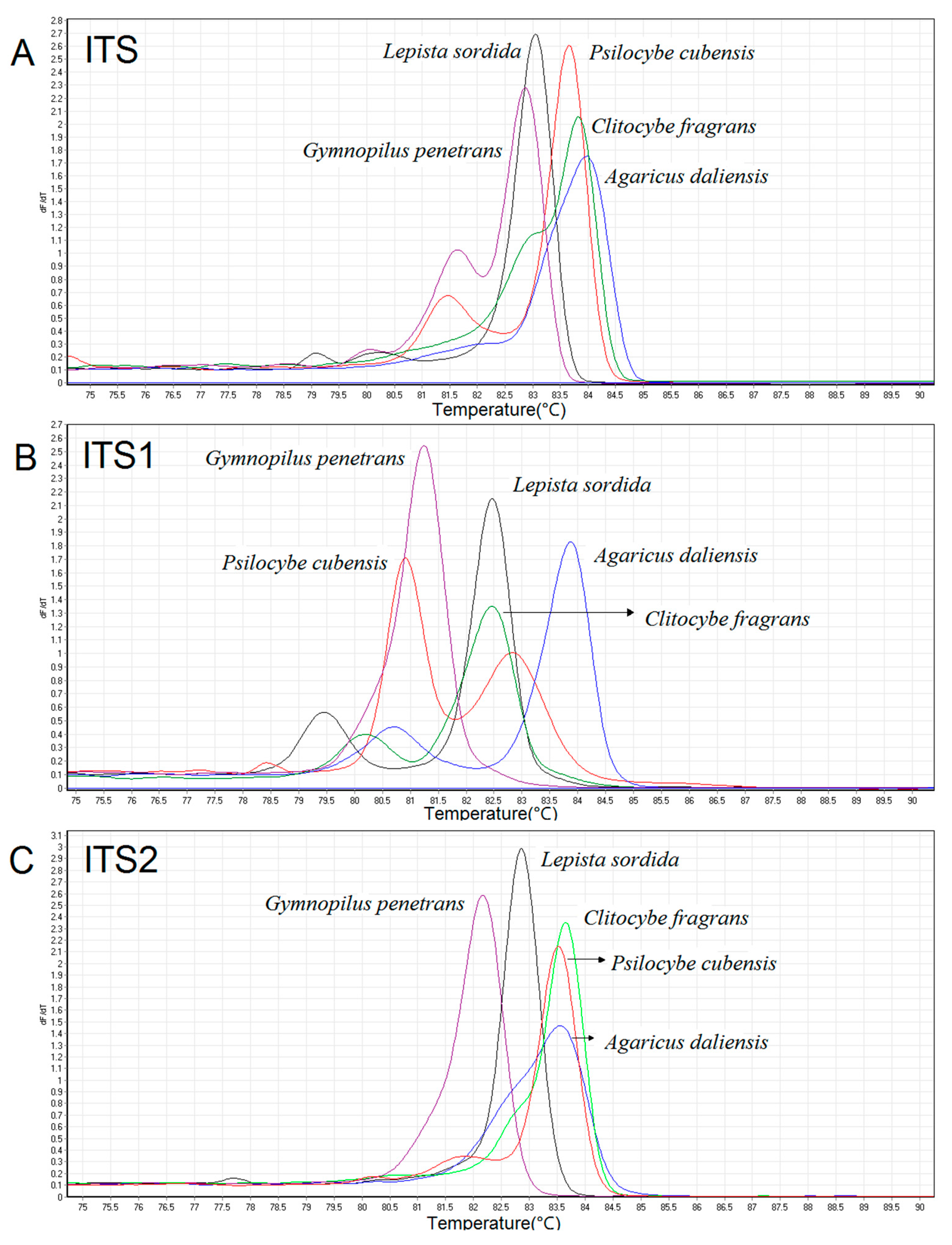
3.1. HRM Specificity Studies
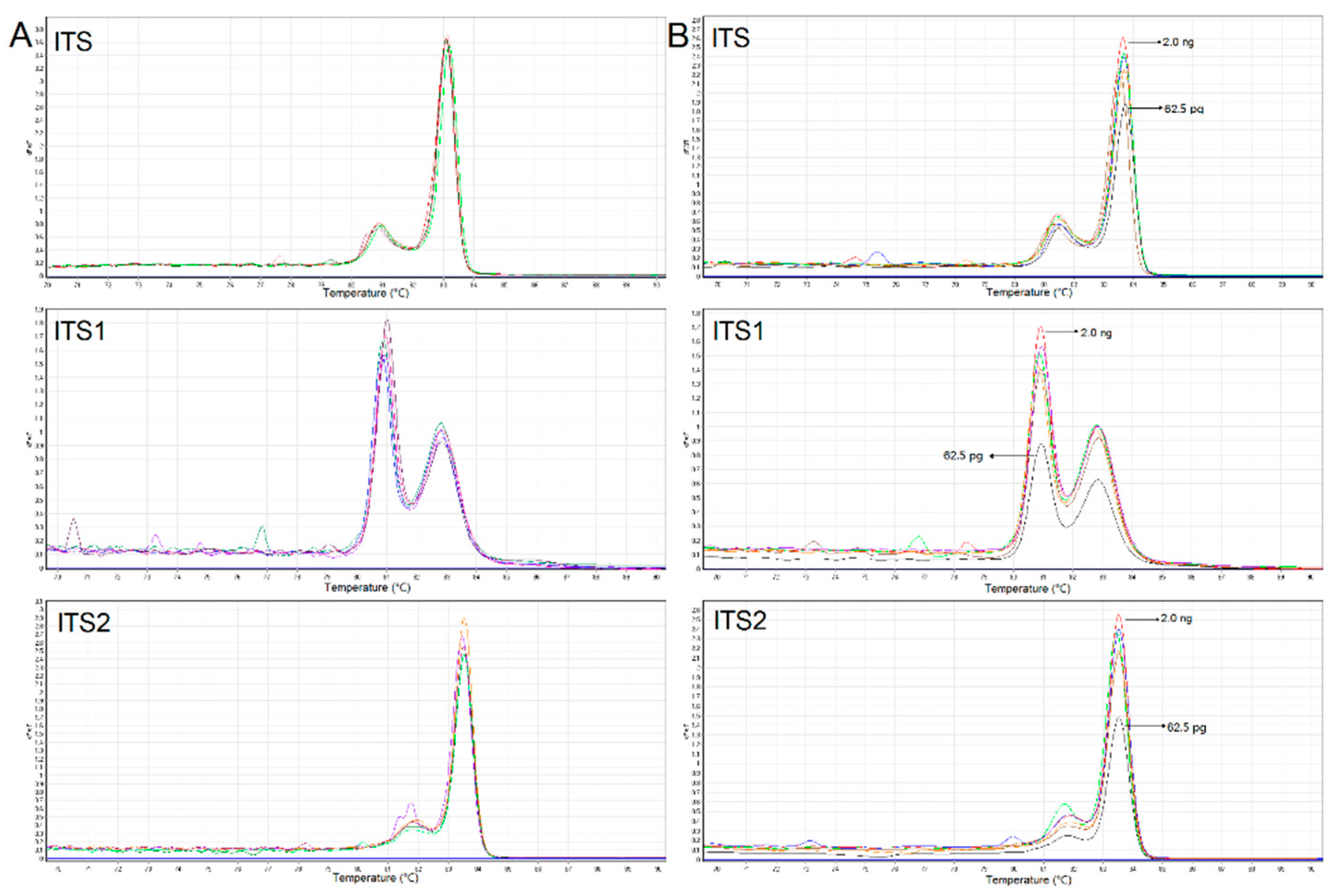
3.2. Reproductivity, Sensitivity, and Mixture Tests
3.3. Species Validation
3.4. Phylogenetic Analysis
3.5. Unknown Sample Detection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kamata, T.; Katagi, M.; Tsuchihashi, H. Metabolism and toxicological analyses of hallucinogenic tryptamine analogues being abused in Japan. Forensic Toxicol. 2010, 28, 1–8. [Google Scholar] [CrossRef]
- Pain, S.; Batisse, A.; Ingrand, I.; Fauconneau, B.; Pérault-Pochat, M.-C. Consumption of hallucinogenic plants and mushrooms by university students in France: A pilot study. Presse Med. 2018, 47, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Carod-Artal, F.J. Hallucinogenic drugs in pre-Columbian Mesoamerican cultures. Neurología 2015, 30, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Wurst, M.; Kysilka, R.; Flieger, M. Psychoactive tryptamines from basidiomycetes. Folia Microbiol. 2002, 47, 3–27. [Google Scholar] [CrossRef] [PubMed]
- International Narcotics Control Board. List of psychotropic substances under international control. Benzodiazepines 2015. [Google Scholar]
- Anastos, N.; Lewis, S.W.; Barnett, N.W.; Sims, D.N. The determination of psilocin and psilocybin in hallucinogenic mushrooms by HPLC utilizing a dual reagent acidic potassium permanganate and tris(2,2′-bipyridyl)ruthenium(II) chemiluminescence detection system. J. Forensic Sci. 2006, 51, 45–51. [Google Scholar] [CrossRef]
- Saito, K.; Toyo’oka, T.; Kato, M.; Fukushima, T.; Shirota, O.; Goda, Y. Determination of psilocybin in hallucinogenic mushrooms by reversed-phase liquid chromatography with fluorescence detection. Talanta 2005, 66, 562–568. [Google Scholar] [CrossRef]
- Christiansen, A.L.; Rasmussen, K.E. Screening of hallucinogenic mushrooms with high-performance liquid chromatography and multiple detection. J. Chromatogr. A 1983, 270, 293–299. [Google Scholar] [CrossRef]
- Guzmán, G.; Allen, J.W.; Gartz, J. A worldwide geographical distribution of the neurotropic fungi, an analysis and discussion. Annali dei Museo Civico Rovereto 1998, 14, 189–280. [Google Scholar]
- Nugent, K.G.; Saville, B.J. Forensic analysis of hallucinogenic fungi: A DNA-based approach. Forensic Sci. Int. 2004, 140, 147–157. [Google Scholar] [CrossRef]
- McLaughlin, D.J.; Hibbett, D.S.; Lutzoni, F.; Spatafora, J.W.; Vilgalys, R. The search for the fungal tree of life. Trends Microbiol. 2009, 17, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badotti, F.; De Oliveira, F.S.; Garcia, C.F.; Vaz, A.B.M.; Fonseca, P.L.C.; Nahum, L.A.; Oliveira, G.; Góes-Neto, A. Effectiveness of ITS and sub-regions as DNA barcode markers for the identification of Basidiomycota (Fungi). BMC Microbiol. 2017, 17, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambaro, V.; Roda, G.; Visconti, G.L.; Arnoldi, S.; Casagni, E.; Dell’Acqua, L.; Farè, F.; Paladino, E.; Rusconi, C.; Arioli, S.; et al. DNA-based taxonomic identification of basidiospores in hallucinogenic mushrooms cultivated in “grow-kits” seized by the police: LC-UV quali-quantitative determination of psilocybin and psilocin. J. Pharm. Biomed. Anal. 2016, 125, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.-I.; Cole, M.; Linacre, A. Identification of members of the genera Panaeolus and Psilocybe by a DNA test. Forensic Sci. Int. 2000, 112, 123–133. [Google Scholar] [CrossRef]
- Lu, S.; Mirchevska, G.; Phatak, S.S.; Li, D.; Luka, J.; Calderone, R.A.; Fonzi, W.A. Dynamic time warping assessment of high-resolution melt curves provides a robust metric for fungal identification. PLoS ONE 2017, 12, e0173320. [Google Scholar] [CrossRef] [Green Version]
- Kallifatidis, B.; Borovička, J.; Stránská, J.; Drábek, J.; Mills, D.K. Fluorescent Random Amplified Microsatellites (F-RAMS) analysis of mushrooms as a forensic investigative tool. Forensic Sci. Int. Genet. 2014, 9, 25–32. [Google Scholar] [CrossRef]
- Mattioni, C.; Casasoli, M.; Gonzalez, M.; Ipinza, R.; Villani, F. Comparison of ISSR and RAPD markers to characterize three Chilean Nothofagus species. Theor. Appl. Genet. 2002, 104, 1064–1070. [Google Scholar] [CrossRef]
- Maeta, K.; Ochi, T.; Tokimoto, K.; Shimomura, N.; Maekawa, N.; Kawaguchi, N.; Nakaya, M.; Kitamoto, Y.; Aimi, T. Rapid species identification of cooked poisonous mushrooms by using real-time PCR. Appl. Environ. Microbiol. 2008, 74, 3306–3309. [Google Scholar] [CrossRef] [Green Version]
- Nicklas, J.A.; Noreault-Conti, T.; Buel, E. Development of a fast, simple profiling method for sample screening using high resolution melting (HRM) of STRs. J. Forensic Sci. 2012, 57, 478–488. [Google Scholar] [CrossRef]
- Elkins, K.M.; Perez, A.C.U.; Quinn, A.A. Simultaneous Identification of Four "Legal High" Plant Species in a Multiplex PCR High-Resolution Melt Assay. J. Forensic Sci. 2017, 62, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Solano, J.; Anabalón, L.; Figueroa, S.; Lizama, C.; Reyes, L.C.; Gangitano, D. Psychedelic fungus (Psilocybe sp.) authentication in a case of illegal drug traffic: Sporological, molecular analysis and identification of the psychoactive substance. Sci. Justice 2019, 59, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Cowan, A.F.; Elkins, K.M. Detection and Identification of Psilocybe cubensis DNA Using a Real-Time Polymerase Chain Reaction High Resolution Melt Assay. J. Forensic Sci. 2018, 63, 1500–1505. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. ISBN 9780123721808. [Google Scholar]
- Libert, X.; Chasseur, C.; Bladt, S.; Packeu, A.; Bureau, F.; Roosens, N.H.; De Keersmaecker, S.C.J. Development and performance assessment of a qualitative SYBR® green real-time PCR assay for the detection of Aspergillus versicolor in indoor air. Appl. Microbiol. Biotechnol. 2015, 99, 7267–7282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiser, D.M.; Klich, M.A.; Frisvad, J.C.; Peterson, S.W.; Varga, J.; Samson, R.A. The current status of species recognition and identification in Aspergillus. Stud. Mycol. 2007, 59, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Skouboe, P.; Frisvad, J.C.; Taylor, J.W.; Lauritsen, D.; Boysen, M.; Rossen, L. Phylogenetic analysis of nucleotide sequences from the ITS region of terverticillate Penicillium species. Mycol. Res. 1999, 103, 873–881. [Google Scholar] [CrossRef]
- Borovička, J.; Noordeloos, M.E.; Gryndler, M.; Oborník, M. Molecular phylogeny of Psilocybe cyanescens complex in Europe, with reference to the position of the secotioid Weraroa novae-zelandiae. Mycol. Prog. 2011, 10, 149–155. [Google Scholar] [CrossRef]
- Mao, F.; Leung, W.-Y.; Xin, X. Characterization of EvaGreen and the implication of its physicochemical properties for qPCR applications. BMC Biotechnol. 2007, 7, 76. [Google Scholar] [CrossRef] [Green Version]
- Mahmoudi, E.; Faizi, M.; Hajiaghaee, R.; Razmi, A. Alteration of Depressive-like Behaviors by Psilocybe cubensis Alkaloid Extract in Mice: The Role of Glutamate Pathway. Res. J. Pharmacogn. 2018, 5, 17–24. [Google Scholar]
- Reeb, V.; Lutzoni, F.; Roux, C. Contribution of RPB2 to multilocus phylogenetic studies of the euascomycetes (Pezizomycotina, Fungi) with special emphasis on the lichen-forming Acarosporaceae and evolution of polyspory. Mol. Phylogenet. Evol. 2004, 32, 1036–1060. [Google Scholar] [CrossRef]
- Frøslev, T.G.; Matheny, P.B.; Hibbett, D.S. Lower level relationships in the mushroom genus Cortinarius (Basidiomycota, Agaricales): A comparison of RPB1, RPB2, and ITS phylogenies. Mol. Phylogenet. Evol. 2005, 37, 602–618. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scheme 5 | Primer | Sequences (5′→3′) | Gene | Tm (°C) | Size of Fragments |
|---|---|---|---|---|---|
| ITS | ITS4 | TCCTCCGCTTATTGATATGC | ITS1-5.8S-ITS2 | 51.7 | ~680 bp |
| ITS5 | GGAAGTAAAAGTCGTAACAAGG | 50.7 | |||
| ITS1 | ITS2 | GCTGCGTTCTTCATCGATGC | ITS1 | 57.4 | ~275 bp |
| ITS5 | GGAAGTAAAAGTCGTAACAAGG | 50.7 | |||
| ITS2 | ITS4 | TCCTCCGCTTATTGATATGC | ITS2 | 51.7 | ~310 bp |
| ITS86 | GTGAATCATCGAATCTTTGAAC | 48.9 |
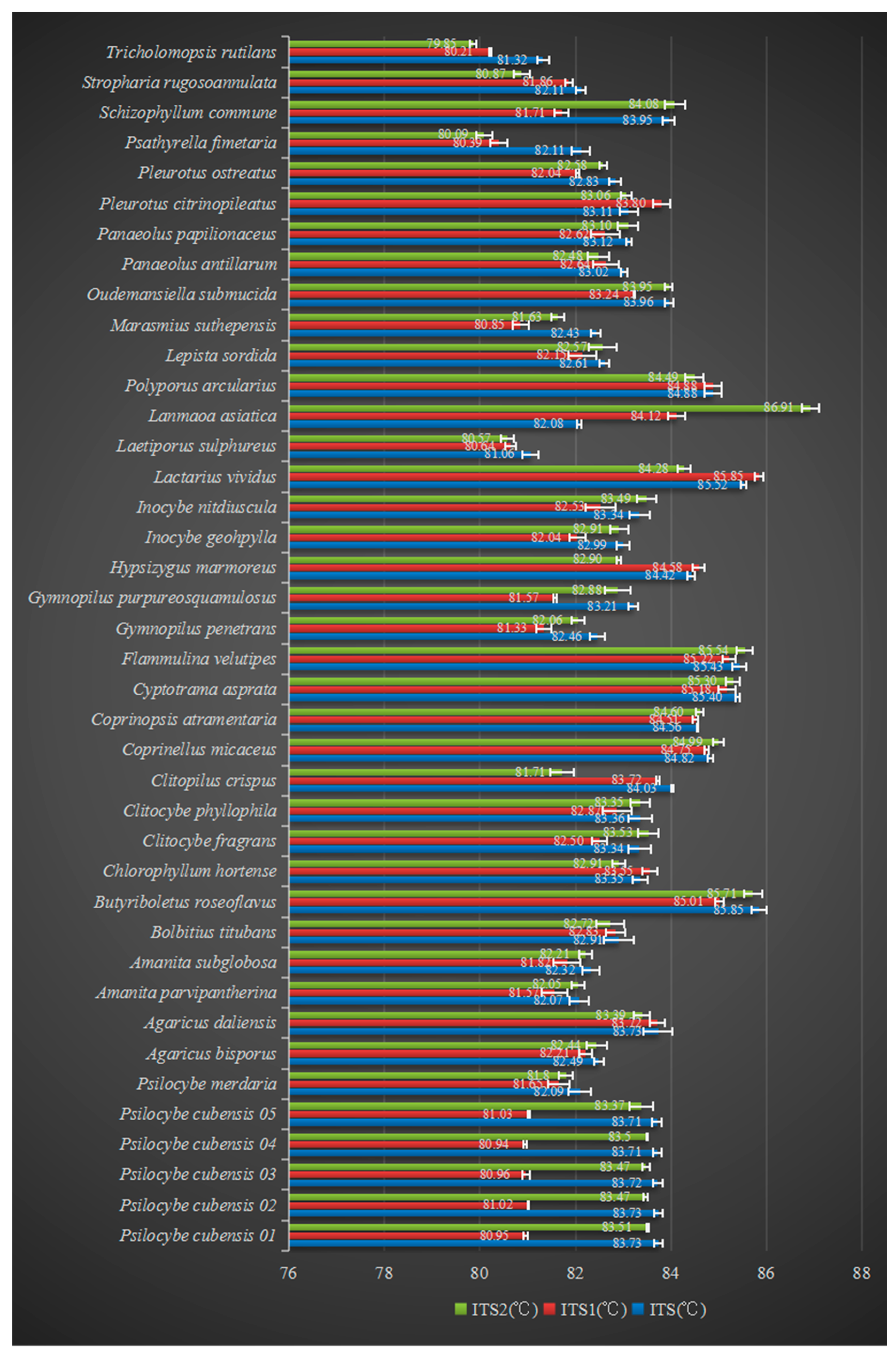
| Species | ITS (°C) | ITS1 (°C) | ITS2 (°C) |
|---|---|---|---|
| Psilocybe cubensis | 83.72 ± 0.01 | 80.98 ± 0.06 | 83.46 ± 0.08 |
| Agaricus daliensis | 83.73 ± 0.30 | 83.72 ± 0.16 | 83.39 ± 0.17 |
| Clitopilus crispus | 84.03 ± 0.03 | 83.72 ± 0.04 | 81.71 ± 0.25 |
| Gymnopilus purpureosquamulosus | 83.21 ± 0.11 | 81.57 ± 0.04 | 82.88 ± 0.27 |
| Hypsizygus marmoreus | 84.42 ± 0.08 | 84.58 ± 0.13 | 82.90 ± 0.05 |
| Lactarius vividus | 85.52 ± 0.05 | 85.85 ± 0.09 | 84.28 ± 0.13 |
| Laetiporus sulphureus | 81.06 ± 0.17 | 80.64 ± 0.11 | 80.57 ± 0.14 |
| Lanmaoa asiatica | 82.08 ± 0.05 | 84.12 ± 0.18 | 86.91 ± 0.18 |
| Marasmius suthepensis | 82.43 ± 0.10 | 80.85 ± 0.17 | 81.63 ± 0.14 |
| Oudemansiella submucida | 83.96 ± 0.08 | 83.24 ± 0.01 | 83.95 ± 0.07 |
| Pleurotus ostreatus | 82.83 ± 0.12 | 82.04 ± 0.03 | 82.58 ± 0.08 |
| Psathyrella fimetaria | 82.11 ± 0.19 | 80.39 ± 0.18 | 80.09 ± 0.17 |
| Schizophyllum commune | 83.95 ± 0.12 | 81.71 ± 0.14 | 84.08 ± 0.21 |
| Stropharia rugosoannulata | 82.11 ± 0.09 | 81.86 ± 0.08 | 80.87 ± 0.17 |
| Tricholomopsis rutilans | 81.32 ± 0.12 | 80.21 ± 0.02 | 79.85 ± 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Yu, H.; Yang, Q.; Wang, Z.; Xia, R.; Chen, C.; Qu, Y.; Tan, R.; Shi, Y.; Xiang, P.; et al. A Forensic Detection Method for Hallucinogenic Mushrooms via High-Resolution Melting (HRM) Analysis. Genes 2021, 12, 199. https://doi.org/10.3390/genes12020199
Zhang X, Yu H, Yang Q, Wang Z, Xia R, Chen C, Qu Y, Tan R, Shi Y, Xiang P, et al. A Forensic Detection Method for Hallucinogenic Mushrooms via High-Resolution Melting (HRM) Analysis. Genes. 2021; 12(2):199. https://doi.org/10.3390/genes12020199
Chicago/Turabian StyleZhang, Xiaochun, Huan Yu, Qi Yang, Ziwei Wang, Ruocheng Xia, Chong Chen, Yiling Qu, Rui Tan, Yan Shi, Ping Xiang, and et al. 2021. "A Forensic Detection Method for Hallucinogenic Mushrooms via High-Resolution Melting (HRM) Analysis" Genes 12, no. 2: 199. https://doi.org/10.3390/genes12020199
APA StyleZhang, X., Yu, H., Yang, Q., Wang, Z., Xia, R., Chen, C., Qu, Y., Tan, R., Shi, Y., Xiang, P., Zhang, S., & Li, C. (2021). A Forensic Detection Method for Hallucinogenic Mushrooms via High-Resolution Melting (HRM) Analysis. Genes, 12(2), 199. https://doi.org/10.3390/genes12020199