Insights into the Transcriptional Regulation of Branching Hormonal Signaling Pathways Genes under Drought Stress in Arabidopsis



Abstract
:1. Introduction
2. Results
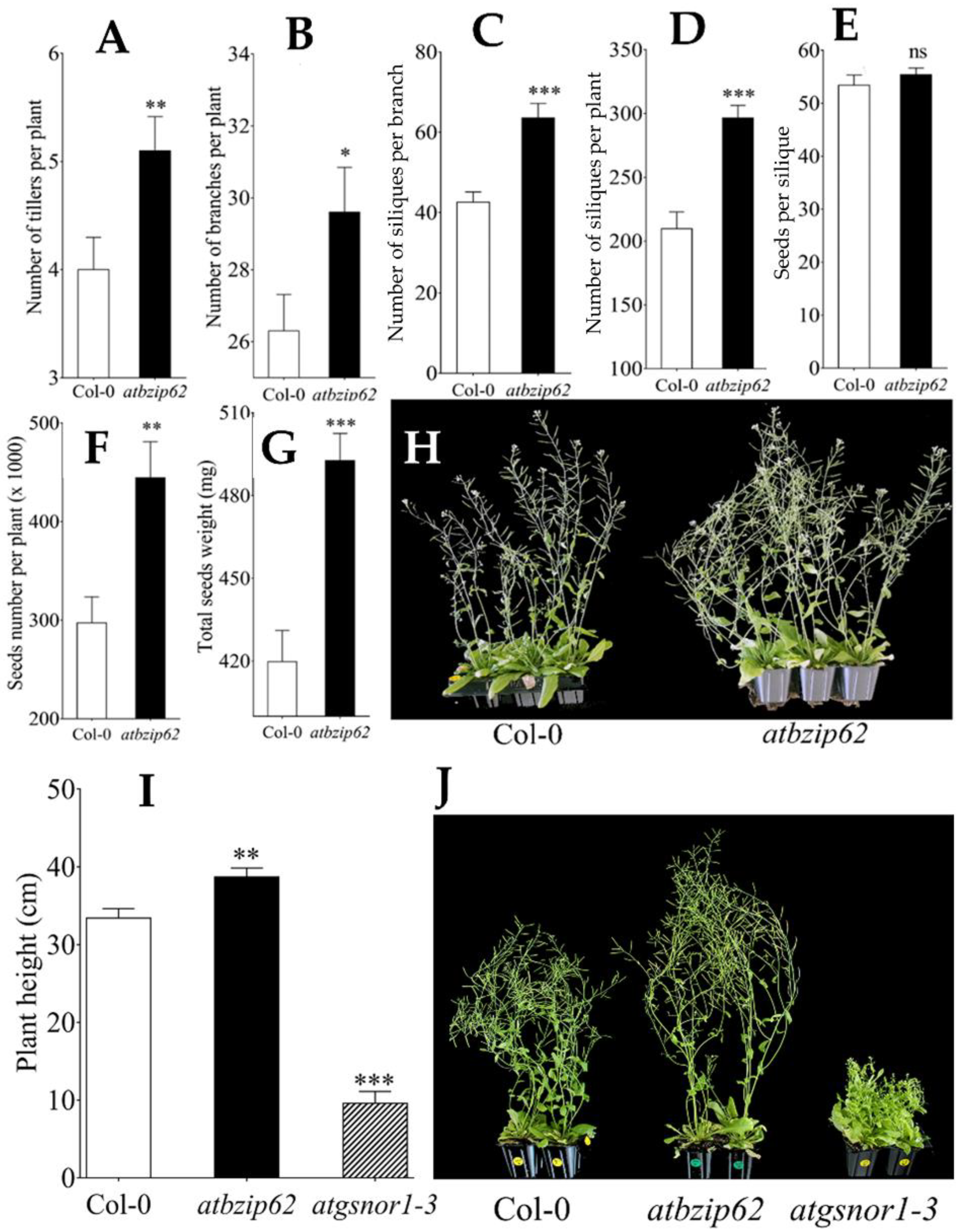
2.1. AtbZIP62 TF Could Be Involved in the Control of Bud Outgrowth in Plants
2.2. In Silico Prediction of Transcription Factor Binding Sites Identified bZIPs Cis-Regulatory Elements
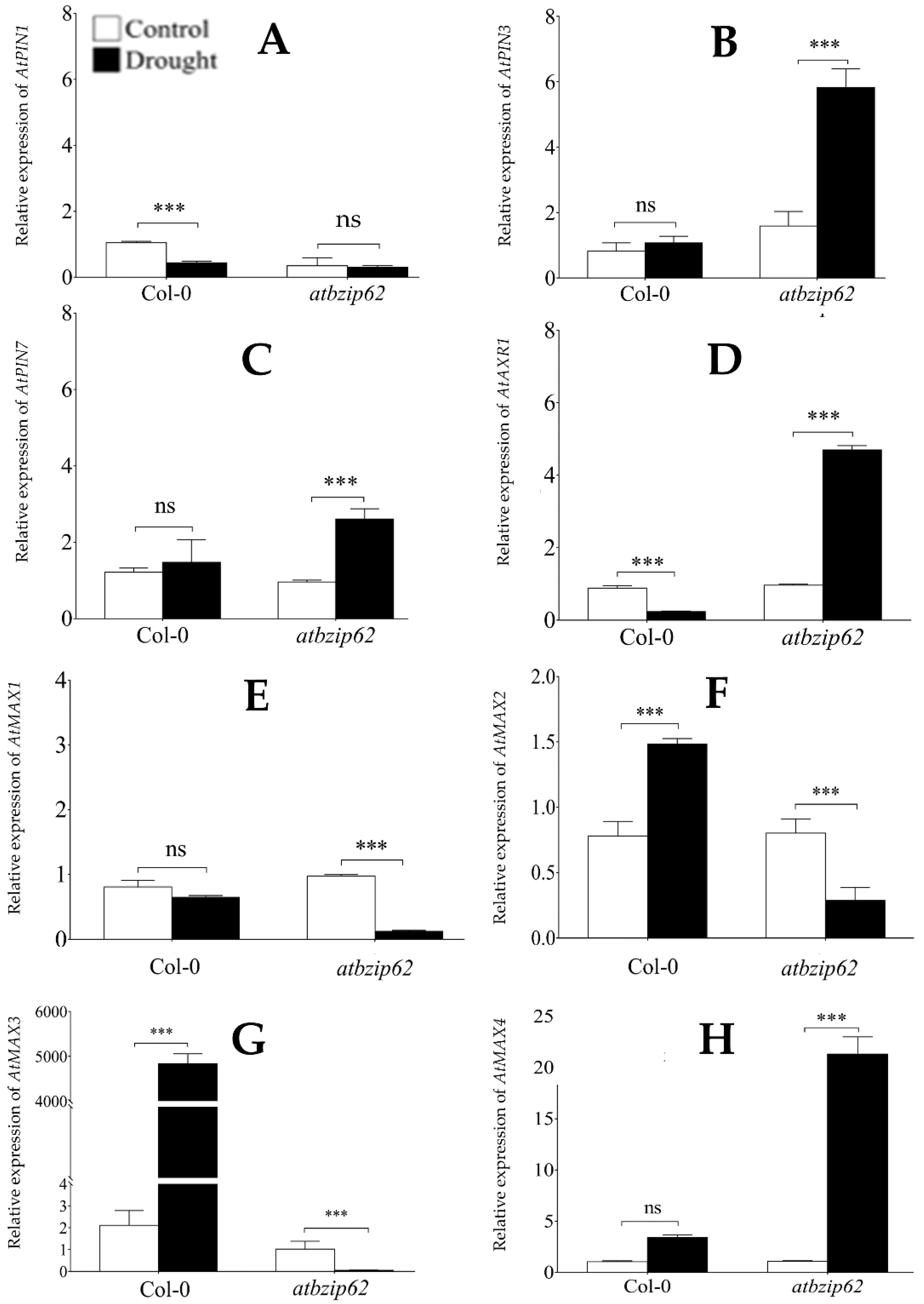
2.3. AtbZIP62 Differentially Regulated PIN-FORMED and MAX Encoding Genes in Response to Drought Stress
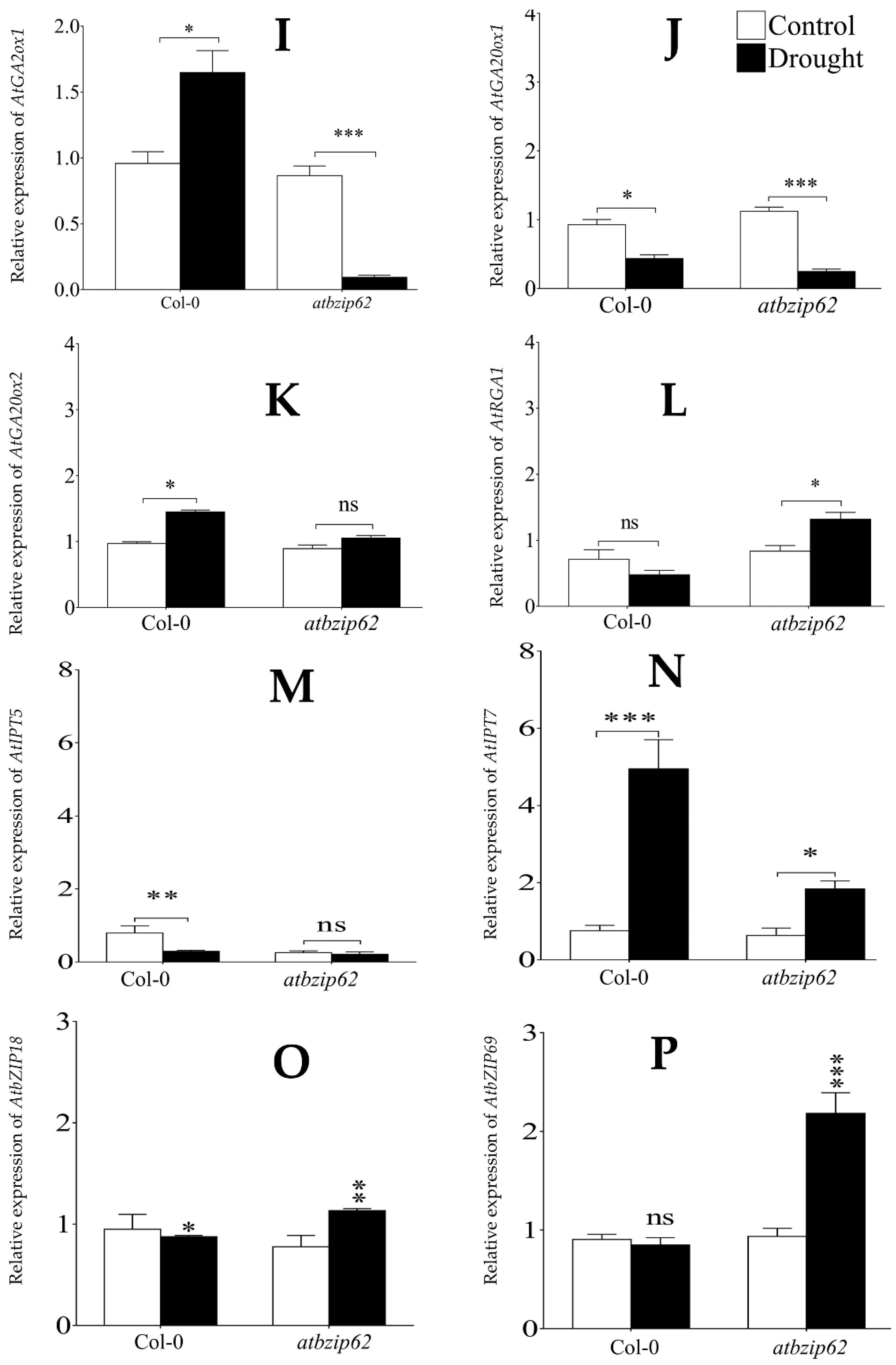
2.4. Drought Stress Differentially Expressed Gibberellic Acid Biosynthetic Genes in Col-0 and atbzip62
2.5. Drought Stress Differentially Regulated AtIPT5 and AtIPT7 in atbzip62
2.6. AtbZIP18 and AtbZIP69 Are Differentially Regulated betwen Col-0 and atbzip62
3. Discussion
3.1. The AtbZIP62 TF Differentially Regulates the Expression of the Arabidopsis PIN-FORMED Protein, MAX, IPT and GA-Biosynthetic Encoding Genes in Response to Drought Stress
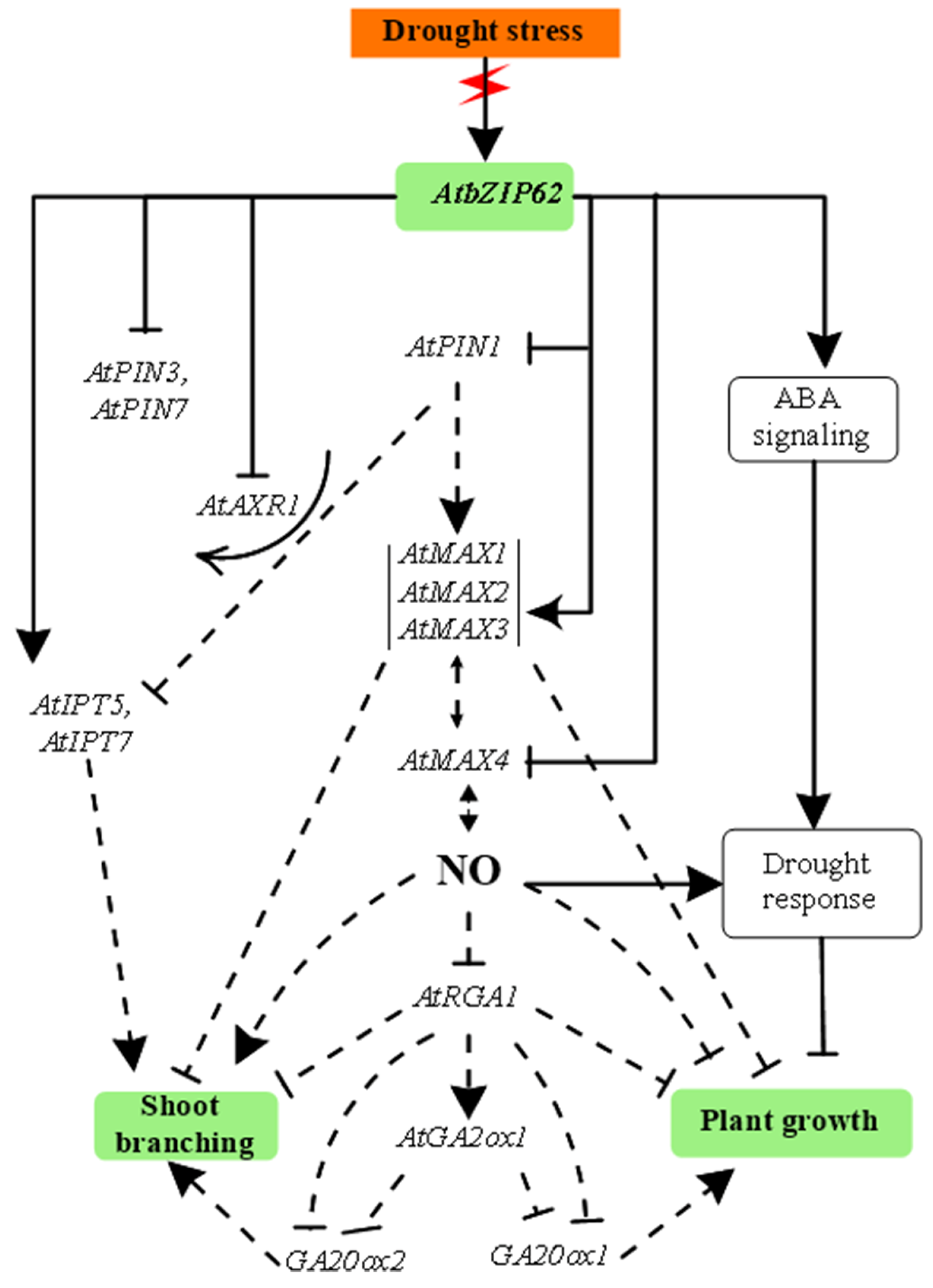
3.2. Proposed Signaling Model of AbZIP62 TF and Homonal Biosynthesic Genes under Drought Stress
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. In Silico Transcription Factor Binding Site Prediction
4.3. Drought Stress Induction by Water Withholding
4.4. Total RNA Isolation and Gene Expression Analysis by qPCR
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef] [Green Version]
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate trends and global crop production since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mafakheri, A.; Siosemardeh, A.; Bahramnejad, B.; Struik, P.; Sohrabi, Y. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 2010, 4, 580–585. [Google Scholar]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol 2009, 11, 100–105. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Borsani, O.; Valpuesta, V.; Botella, M.A. Evidence for a role of salicylic acid in the oxidative damage generated by NaCl and osmotic stress in Arabidopsis seedlings. Plant Physiol. 2001, 126, 1024–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Verma, S.; Dubey, R. Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Sci. 2003, 164, 645–655. [Google Scholar] [CrossRef]
- Meriga, B.; Reddy, B.K.; Rao, K.R.; Reddy, L.A.; Kishor, P.K. Aluminium-induced production of oxygen radicals, lipid peroxidation and DNA damage in seedlings of rice (Oryza sativa). J. Plant Physiol. 2004, 161, 63–68. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Riadh, K.; Gopi, R.; Manivannan, P.; Ines, J.; Al-Juburi, H.J.; Chang-Xing, Z.; Hong-Bo, S.; Panneerselvam, R. Antioxidant defense responses: Physiological plasticity in higher plants under abiotic constraints. Acta Physiol. Plant. 2009, 31, 427–436. [Google Scholar] [CrossRef]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef] [Green Version]
- Corpas, F.J. What is the role of hydrogen peroxide in plant peroxisomes? Plant Biol. 2015, 17, 1099–1103. [Google Scholar] [CrossRef]
- Hoang, X.L.T.; Du Nhi, N.H.; Binh Anh Thu, N.; Phuong Thao, N.; Phan Tran, L.-S.J.C.g. Transcription factors and their roles in signal transduction in plants under abiotic stresses. Curr. Genom. 2017, 18, 483–497. [Google Scholar] [CrossRef]
- Khan, S.-A.; Li, M.-Z.; Wang, S.-M.; Yin, H.J. Revisiting the role of plant transcription factors in the battle against abiotic stress. Int. J. Mol. Sci. 2018, 19, 1634. [Google Scholar] [CrossRef] [Green Version]
- Crawford, T.; Karamat, F.; Lehotai, N.; Rentoft, M.; Blomberg, J.; Strand, Å.; Björklund, S. Specific functions for Mediator complex subunits from different modules in the transcriptional response of Arabidopsis thaliana to abiotic stress. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Gale, M.; Devos, K. Plant comparative genetics after 10 years. Science 1998, 282, 656–659. [Google Scholar] [CrossRef] [Green Version]
- Dodeweerd, A.-M.v.; Hall, C.R.; Bent, E.G.; Johnson, S.J.; Bevan, M.W.; Bancroft, I. Identification and analysis of homoeologous segments of the genomes of rice and Arabidopsis thaliana. Genome 1999, 42, 887–892. [Google Scholar] [CrossRef]
- Liu, H.; Sachidanandam, R.; Stein, L. Comparative genomics between rice and Arabidopsis shows scant collinearity in gene order. Genome Res. 2001, 11, 2020–2026. [Google Scholar] [CrossRef] [Green Version]
- Watson, D.K.; Kitching, R.; Vary, C.; Kola, I.; Seth, A. Isolation of target gene promoter/enhancer sequences by whole genome PCR method. Methods Mol. Biol. (Clifton, N.J.) 2000, 130, 1–11. [Google Scholar]
- Perry, S.E. Plant Transcription Factors: Methods and Protocols; Springer: Berlin, Germany, 2011. [Google Scholar]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002; Volume 91, p. 401. [Google Scholar]
- Jakoby, M.; Weisshaar, B.; Droge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Ali, Z.; Sarwat, S.S.; Karim, I.; Faridi, R.; Jaskani, M.J.; Khan, A.A. Functions of plant’s bZIP transcription factors. Pak. J. Agric. Sci. 2016, 53. [Google Scholar] [CrossRef]
- Ábrahám, E.; Hourton-Cabassa, C.; Erdei, L.; Szabados, L. Methods for determination of proline in plants. In Methods in Molecular Biology; Springer International Publishing: Berlin, Germany, 2010; Volume 639, pp. 317–331. [Google Scholar]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Rolly, N.K.; Imran, Q.M.; Shahid, M.; Imran, M.; Khan, M.; Lee, S.-U.; Hussain, A.; Lee, I.-J.; Yun, B.-W. Drought-induced AtbZIP62 transcription factor regulates drought stress response in Arabidopsis. Plant Physiol. Biochem. 2020, 156, 384–395. [Google Scholar] [CrossRef]
- Rolly, N.K.; Imran, Q.M.; Lee, I.-J.; Yun, B.-W. Salinity Stress-Mediated Suppression of Expression of Salt Overly Sensitive Signaling Pathway Genes Suggests Negative Regulation by AtbZIP62 Transcription Factor in Arabidopsis thaliana. Int. J. Mol. Sci. 2020, 21, 1726. [Google Scholar] [CrossRef] [Green Version]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nat. Cell Biol. 2005, 433, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Křeček, P.; Skůpa, P.; Libus, J.; Naramoto, S.; Tejos, R.; Friml, J.; Zažímalová, E. The PIN-FORMED (PIN) protein family of auxin transporters. Genome Biol. 2009, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keuskamp, D.H.; Pollmann, S.; Voesenek, L.A.; Peeters, A.J.; Pierik, R. Auxin transport through PIN-FORMED 3 (PIN3) controls shade avoidance and fitness during competition. Proc. Nat. Acad. Sci. USA 2010, 107, 22740–22744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friml, J.; Wiśniewska, J.; Benková, E.; Mendgen, K.; Palme, K. Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nat. Cell Biol. 2002, 415, 806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakeslee, J.J.; Peer, W.A.; Murphy, A.S. Auxin transport. Curr. Opin. Plant Biol. 2005, 8, 494–500. [Google Scholar] [CrossRef]
- Friml, J.; Vieten, A.; Sauer, M.; Weijers, D.; Schwarz, H.; Hamann, T.; Offringa, R.; Jürgens, G. Efflux-dependent auxin gradients establish the apical–basal axis of Arabidopsis. Nat. Cell Biol. 2003, 426, 147. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Raman, A.S.; Ream, J.E.; Fujiwara, H.; Cerny, R.E.; Brown, S.M. Overexpression of 20-oxidase confers a gibberellin-overproduction phenotype in Arabidopsis. Plant Physiol. 1998, 118, 773–781. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, A. Effect of High Temperature on Crop Productivity and Metabolism of Macro Molecules; Elsevier BV: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Phillips, A.L.; Ward, D.A.; Uknes, S.; Appleford, N.E.; Lange, T.; Huttly, A.K.; Gaskin, P.; Graebe, J.E.; Hedden, P. Isolation and expression of three gibberellin 20-oxidase cDNA clones from Arabidopsis. Plant Physiol. 1995, 108, 1049–1057. [Google Scholar] [CrossRef] [Green Version]
- Gilmour, S.J.; Zeevaart, J.A.; Schwenen, L.; Graebe, J.E. Gibberellin metabolism in cell-free extracts from spinach leaves in relation to photoperiod. Plant Physiol. 1986, 82, 190–195. [Google Scholar] [CrossRef] [Green Version]
- Hedden, P.; Croker, S.J. Regulation of gibberellin biosynthesis in maize seedlings. In Red. Clover Science; Springer International Publishing: Berlin, Germany, 1992; pp. 534–544. [Google Scholar]
- Xu, Y.-L.; Li, L.; Wu, K.; Peeters, A.; Gage, D.A.; Zeevaart, J. The GA5 locus of Arabidopsis thaliana encodes a multifunctional gibberellin 20-oxidase: Molecular cloning and functional expression. Proc. Nat. Acad. Sci. USA 1995, 92, 6640–6644. [Google Scholar] [CrossRef] [Green Version]
- Pysh, L.D.; Wysocka-Diller, J.W.; Camilleri, C.; Bouchez, D.; Benfey, P.N. The GRAS gene family in Arabidopsis: Sequence characterization and basic expression analysis of the SCARECROW-LIKE genes. Plant J. 1999, 18, 111–119. [Google Scholar] [CrossRef]
- Silverstone, A.L.; Ciampaglio, C.N.; Sun, T.-p. The Arabidopsis RGA gene encodes a transcriptional regulator repressing the gibberellin signal transduction pathway. Plant Cell 1998, 10, 155–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakimoto, T. Identification of plant cytokinin biosynthetic enzymes as dimethylallyl diphosphate: ATP/ADP isopentenyltransferases. Plant Cell Physiol. 2001, 42, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Takei, K.; Sakakibara, H.; Sugiyama, T. Genes: Structure and Regulation-Identification of genes encoding adenylate isopentenyltransferase, a cytokinin biosynthesis enzyme, in Arabidopsis thaliana. J. Biol. Chem. 2001, 276, 26405–26410. [Google Scholar] [CrossRef] [Green Version]
- Leyser, H.O.; Lincoln, C.A.; Timpte, C.; Lammer, D.; Turner, J.; Estelle, M. Arabidopsis auxin-resistance gene AXR1 encodes a protein related to ubiquitin-activating enzyme E1. Nature 1993, 364, 161–164. [Google Scholar] [CrossRef]
- Bennett, T.; Sieberer, T.; Willett, B.; Booker, J.; Luschnig, C.; Leyser, O. The Arabidopsis MAX pathway controls shoot branching by regulating auxin transport. Curr. Biol. 2006, 16, 553–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, B.J.; Beveridge, C.A. Roles for auxin, cytokinin, and strigolactone in regulating shoot branching. Plant Physiol. 2009, 149, 1929–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booker, J.; Sieberer, T.; Wright, W.; Williamson, L.; Willett, B.; Stirnberg, P.; Turnbull, C.; Srinivasan, M.; Goddard, P.; Leyser, O. MAX1 encodes a cytochrome P450 family member that acts downstream of MAX3/4 to produce a carotenoid-derived branch-inhibiting hormone. Dev. Cell 2005, 8, 443–449. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Mun, B.-G.; Imran, Q.M.; Lee, S.-U.; Adamu, T.A.; Shahid, M.; Kim, K.-M.; Yun, B.-W. Nitric oxide mediated transcriptome profiling reveals activation of multiple regulatory pathways in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, D.; Leyser, O. Auxin, cytokinin and the control of shoot branching. Ann. Bot. 2011, 107, 1203–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauser, F.; Waadt, R.; Schroeder, J.I. Evolution of abscisic acid synthesis and signaling mechanisms. Curr. Biol. 2011, 21, R346–R355. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Zou, H.-F.; Wei, W.; Hao, Y.-J.; Tian, A.-G.; Huang, J.; Liu, Y.-F.; Zhang, J.-S.; Chen, S.-Y. Soybean GmbZIP44, GmbZIP62 and GmbZIP78 genes function as negative regulator of ABA signaling and confer salt and freezing tolerance in transgenic Arabidopsis. Planta 2008, 228, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Penfield, S.; Gilday, A.D.; Halliday, K.J.; Graham, I.A. DELLA-mediated cotyledon expansion breaks coat-imposed seed dormancy. Curr. Biol. 2006, 16, 2366–2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willige, B.C.; Isono, E.; Richter, R.; Zourelidou, M.; Schwechheimer, C. Gibberellin regulates PIN-FORMED abundance and is required for auxin transport–dependent growth and development in Arabidopsis thaliana. Plant Cell 2011, 23, 2184–2195. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Feng, F.; Liu, J.; Zhao, Q. The interaction between auxin and nitric oxide regulates root growth in response to iron deficiency in rice. Front. Plant Sci. 2017, 8, 2169. [Google Scholar] [CrossRef]
- Shen, Q.; Wang, Y.-T.; Tian, H.; Guo, F.-Q. Nitric oxide mediates cytokinin functions in cell proliferation and meristem maintenance in Arabidopsis. Mol. Plant 2013, 6, 1214–1225. [Google Scholar] [CrossRef] [Green Version]
- Oláh, D.; Feigl, G.; Molnár, Á.; Ördög, A.; Kolbert, Z. Strigolactones interact with nitric oxide in regulating root system architecture of Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 1019. [Google Scholar] [CrossRef] [PubMed]
- Sanz, L.; Albertos, P.; Mateos, I.; Sánchez-Vicente, I.; Lechón, T.; Fernández-Marcos, M.; Lorenzo, O. Nitric oxide (NO) and phytohormones crosstalk during early plant development. J. Exp. Bot. 2015, 66, 2857–2868. [Google Scholar] [CrossRef] [PubMed]
- Rosso, M.G.; Li, Y.; Strizhov, N.; Reiss, B.; Dekker, K.; Weisshaar, B. An Arabidopsis thaliana T-DNA mutagenized population (GABI-Kat) for flanking sequence tag-based reverse genetics. Plant Mol. Biol. 2003, 53, 247–259. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Tech. Adv. 1998, 16, 735–743. [Google Scholar]
- Harb, A.; Pereira, A. Screening Arabidopsis genotypes for drought stress resistance. Methods Mol. Biol. 2010, 678, 191–198. [Google Scholar] [CrossRef]
- Mun, B.-G.; Lee, S.-U.; Hussain, A.; Kim, H.-H.; Rolly, N.K.; Jung, K.-H.; Yun, B.-W. S-nitrosocysteine-responsive genes modulate diverse regulatory pathways in Oryza sativa: A transcriptome profiling study. Funct. Plant Biol. 2018, 45, 630–644. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Target Genes | Position | Strand | p-Value | q-Value | Matched Sequence |
|---|---|---|---|---|---|---|---|
| * | * | AtMAX1 | * | * | * | * | * |
| * | * | AT2G26170 | * | * | * | * | * |
| AtMAX2 | |||||||
| AT2G40620 | AtbZIP18 | AT2G42620 | 1723–1733 | - | 2.14 × 10−5 | 0.046 | CTCGGCTGGCC |
| AT2G40620 | AtbZIP18 | AT2G42620 | 923–933 | + | 7.04 × 10−6 | 0.0303 | TTCAGCTGTCA |
| AT1G06070 | AtbZIP69 | AT2G42620 | 923–933 | + | 1.41 × 10−5 | 0.0583 | TTCAGCTGTCA |
| AtMAX3 | |||||||
| AT2G40620 | AtbZIP18 | AT2G44990 | 1704–1714 | - | 6.59 × 10−5 | 0.295 | AATAGCTGTCG |
| AtMAX4 | |||||||
| AT2G40620 | AtbZIP18 | AT4G32810 | 2535–2545 | - | 2.48 × 10−5 | 0.158 | CACGGCTGTCT |
| AT2G40620 | AtbZIP18 | AT4G32810 | 285–295 | + | 8.64 × 10−5 | 0.276 | CAGAGCTGTAA |
| AT1G06070 | AtbZIP69 | AT4G32810 | 2535–2545 | - | 6.02 × 10−5 | 0.369 | CACGGCTGTCT |
| AtPIN1 | |||||||
| AT1G06070 | AtbZIP69 | AT1G73590 | 1935–1945 | - | 2.08 × 10−5 | 0.138 | AAGAGCTGGCA |
| AtPIN3 | |||||||
| AT2G40620 | AtbZIP18 | AT1G70940 | 2256–2266 | - | 1.97 × 10−5 | 0.131 | CCCACCTGTCG |
| AT1G06070 | AtbZIP69 | AT1G70940 | 2256–2266 | - | 5.15 × 10−5 | 0.336 | CCCACCTGTCG |
| AtPIN7 | |||||||
| AT2G40620 | AtbZIP18 | AT1G23080 | 49–59 | + | 9.48 × 10−5 | 0.628 | AAAAGCTGTAA |
| AtAXR1 | |||||||
| AT2G40620 | AtbZIP18 | AT1G05180 | 1160–1170 | + | 4.52 × 10−5 | 0.338 | ACCACCTGTCT |
| AT1G06070 | AtbZIP69 | AT1G05180 | 1035–1045 | + | 4.37 × 10−5 | 0.254 | TGCAGCTGGTG |
| AT1G06070 | AtbZIP69 | AT1G05180 | 1160–1170 | + | 6.95 × 10−5 | 0.254 | ACCACCTGTCT |
| AtIPT5 | |||||||
| AT1G06070 | AtbZIP69 | AT5G19040 | 1294–1304 | - | 8.78 × 10−5 | 0.292 | GGCGGCTGGAA |
| * | * | AtGA2ox1 | * | * | * | * | * |
| * | * | AtGA20ox1 | * | * | * | * | * |
| * | * | AtGA20ox2 | * | * | * | * | * |
| * | * | AtRGA1 | * | * | * | * | * |
| Gene Name/Genotype | Locus/ SALK | Forward Primer (5′->3′) | Reverse Primer (5′->3′) | Gene Name |
|---|---|---|---|---|
| atbzip62 | SALK_053908C | TGGCACTTTTAACTTTGTGCC | TACGTTTCCATCGAGTGAACC | Arabidopsis bzip62 loss of function mutant |
| Drought responsive gene in Arabidopsis | ||||
| AtbZIP62 | AT1G19490 | CATCGAGTTGTTGCTCGTCG | AAATCCGCCAATGCTTCTGC | Basic-leucine zipper transcription factor encoding gene 62 |
| Genes involved in strigolactone biosynthesis pathway and controlling shoot branching (bud outgrowth) | ||||
| AtMAX1 | AT2G26170 | TGGTCACTTGCCCTTGATGG | GGTTGCCTCCCCATCTGAAA | More axillary branching 1 gene |
| AtMAX2 | AT2G42620 | CCGAGCCAGAGTTTGGGTTA | GTGCGAAACCGATTGTGTCC | More axillary branching 2 gene |
| AtMAX3 | AT2G44990 | CGTTGGTGAGCCCATGTTTG | TCCACCGAAACCGCATACTC | More axillary branching 3 gene |
| AtMAX4 | AT4G32810 | TATCGGGTCGTGAGGATGGA | GCAAACGAATGGACCCAACC | More axillary branching 4 gene |
| Genes involved in Auxin polar transport (efflux carrier) and controlling shoot branching | ||||
| AtPIN1 | AT1G73590 | ACGACAACCAGTACGTGGAG | TATGTTGTTCCCACCGTCCG | PIN-FORMED (PIN) protein encoding gene1 |
| AtPIN3 | AT1G70940 | TGGCCATGATCCTCGCTTAC | CGAAGATGGCGACAAAACGG | PIN-FORMED (PIN) protein encoding gene 3 |
| AtPIN7 | AT1G23080 | AGCCATGATCCTCGCTTACG | AGAGGGACGGCGAAAATAGC | PIN-FORMED (PIN) protein encoding gene 7 |
| Cytokinin biosynthetic pathway genes | ||||
| AtIPT5 | AT5G19040 | CGACGGAGGTTTTTCTCCGA | GAACTTTTCGACGGCGAGTG | Isopentenyl transferase encoding gene 5 |
| AtIPT7 | AT3G23630 | GACGCCACTGAGGTGTTCTT | CGACGATTCTCTCGCTTGGT | Isopentenyl transferase encoding gene 7 |
| Genes involved in Gibberellic acid biosynthesis pathway | ||||
| AtGA2ox1 | AT1G78440 | CTCGTTGCCCAAGTCAGAGA | TACTCAACCCAACCCACGTC | Gibberellic acid 2 oxidase 1 |
| AtGA20ox1 | AT4G25420 | GTGAGAGTGTTGGCTACGCA | CTCATGTCGTCGCAAAACCG | Gibberellic acid 20 oxidase 1 |
| AtGA20ox2 | AT5G51810 | TGGCCAGACGAAGAGAAACC | TTGACGACGAGGAAGAAGCC | Gibberellic acid 20 oxidase 2 |
| AtAXR1 | AT1G05180 | CGGACAGATTTGCTGCCAAC | ATCTGGGAGTACTGAGCCGT | Arabidopsis Auxin repressor 1 |
| AtRGA1 | AT2G01570 | TTGTCCAACCACGGGACTTC | AGCTCGTCGTCCATGTTACC | Arabidopsis Repressor of GA 1 |
| Arabidopsis housekeeping gene | ||||
| AtACT2 | AT3G18780 | AGGTTCTGTTCCAGCCATC | TTAGAAGCATTTCCTGTGAAC | Arabidopsis Actin coding gene 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rolly, N.K.; Mun, B.-G.; Yun, B.-W. Insights into the Transcriptional Regulation of Branching Hormonal Signaling Pathways Genes under Drought Stress in Arabidopsis. Genes 2021, 12, 298. https://doi.org/10.3390/genes12020298
Rolly NK, Mun B-G, Yun B-W. Insights into the Transcriptional Regulation of Branching Hormonal Signaling Pathways Genes under Drought Stress in Arabidopsis. Genes. 2021; 12(2):298. https://doi.org/10.3390/genes12020298
Chicago/Turabian StyleRolly, Nkulu Kabange, Bong-Gyu Mun, and Byung-Wook Yun. 2021. "Insights into the Transcriptional Regulation of Branching Hormonal Signaling Pathways Genes under Drought Stress in Arabidopsis" Genes 12, no. 2: 298. https://doi.org/10.3390/genes12020298
APA StyleRolly, N. K., Mun, B. -G., & Yun, B. -W. (2021). Insights into the Transcriptional Regulation of Branching Hormonal Signaling Pathways Genes under Drought Stress in Arabidopsis. Genes, 12(2), 298. https://doi.org/10.3390/genes12020298