
A Hypomorphic Mutant of PHD Domain Protein Male Meiocytes Death 1

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Cytology and Microscopy
2.3. Identification of the Causative Mutation
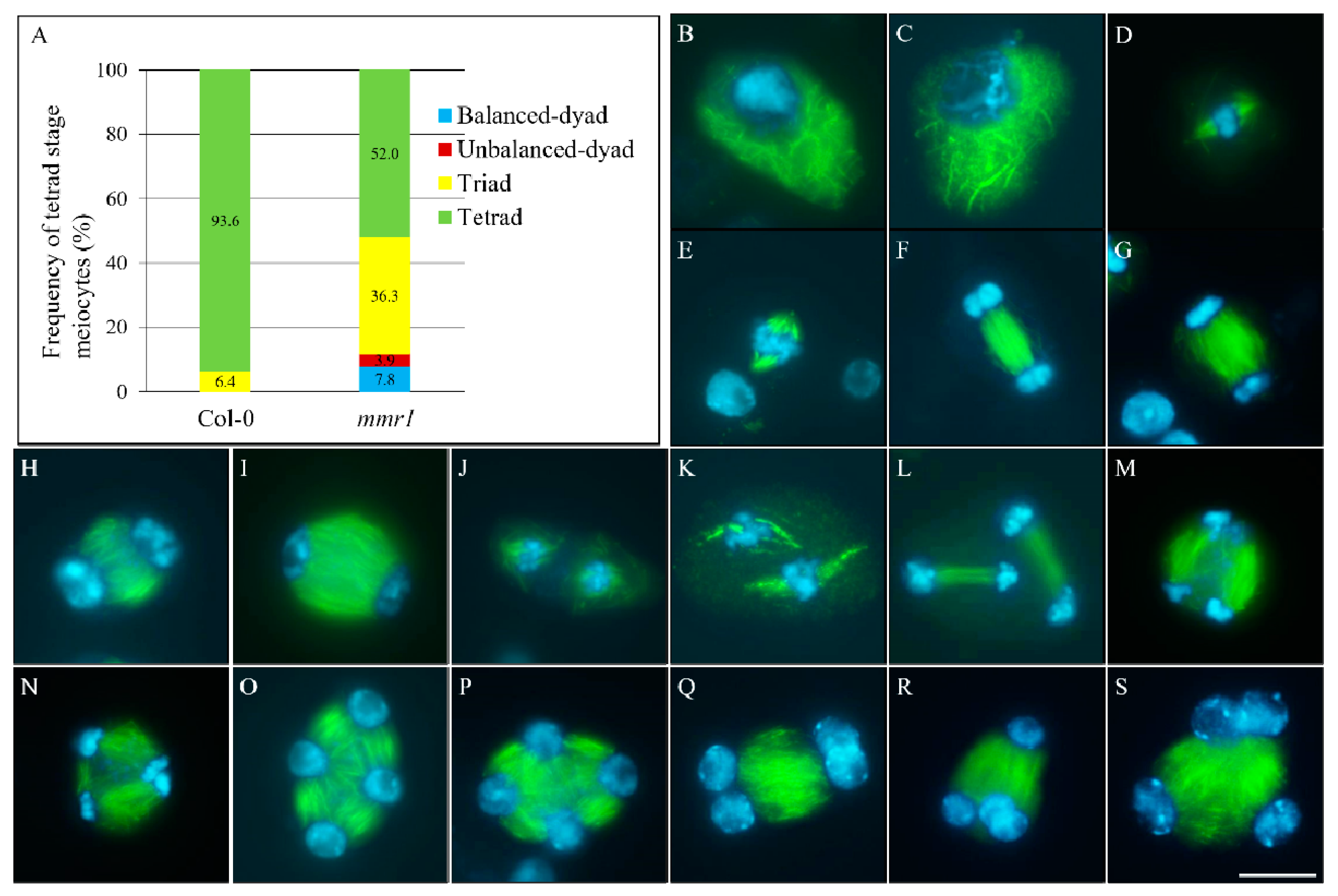
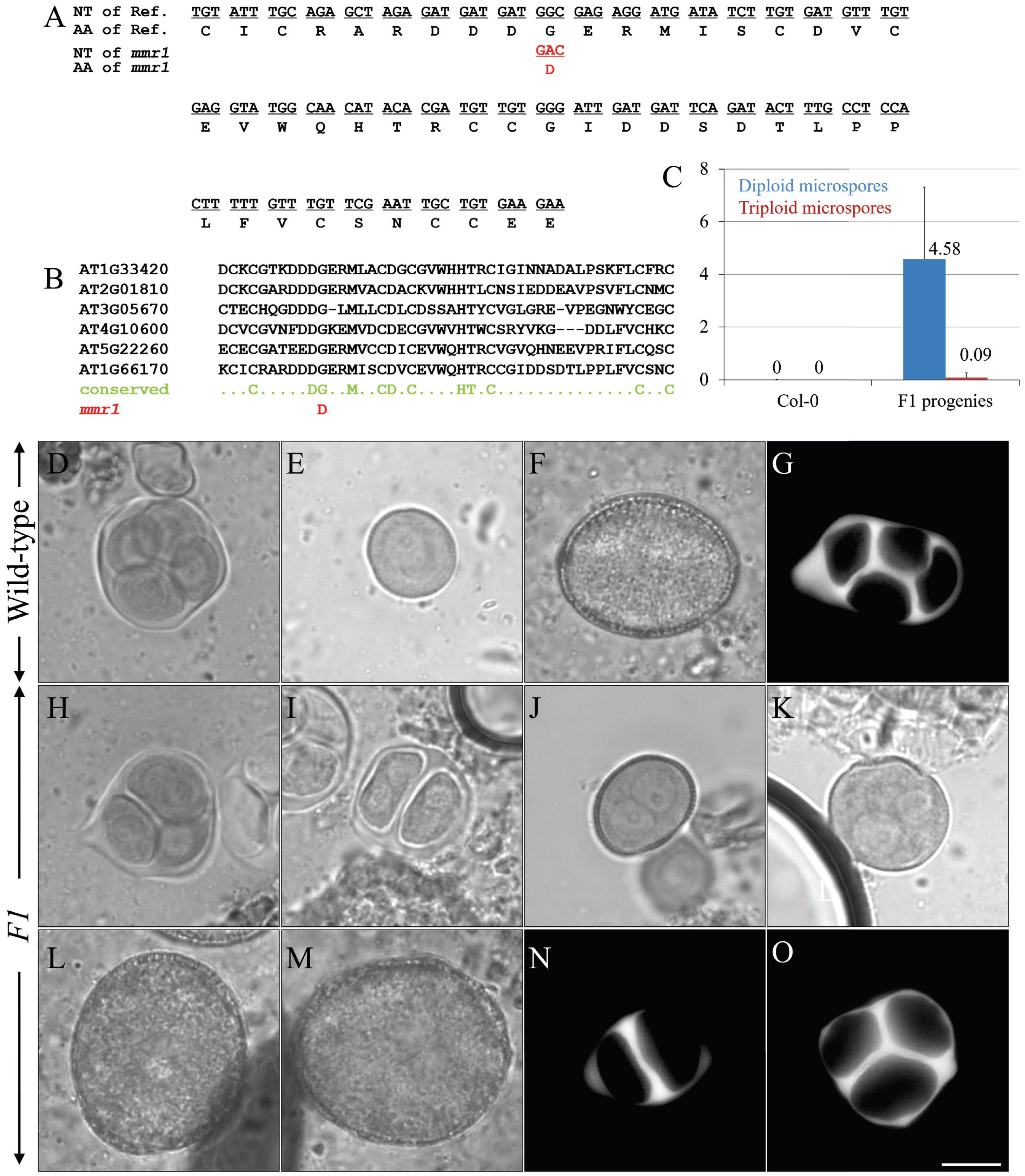
3. Results
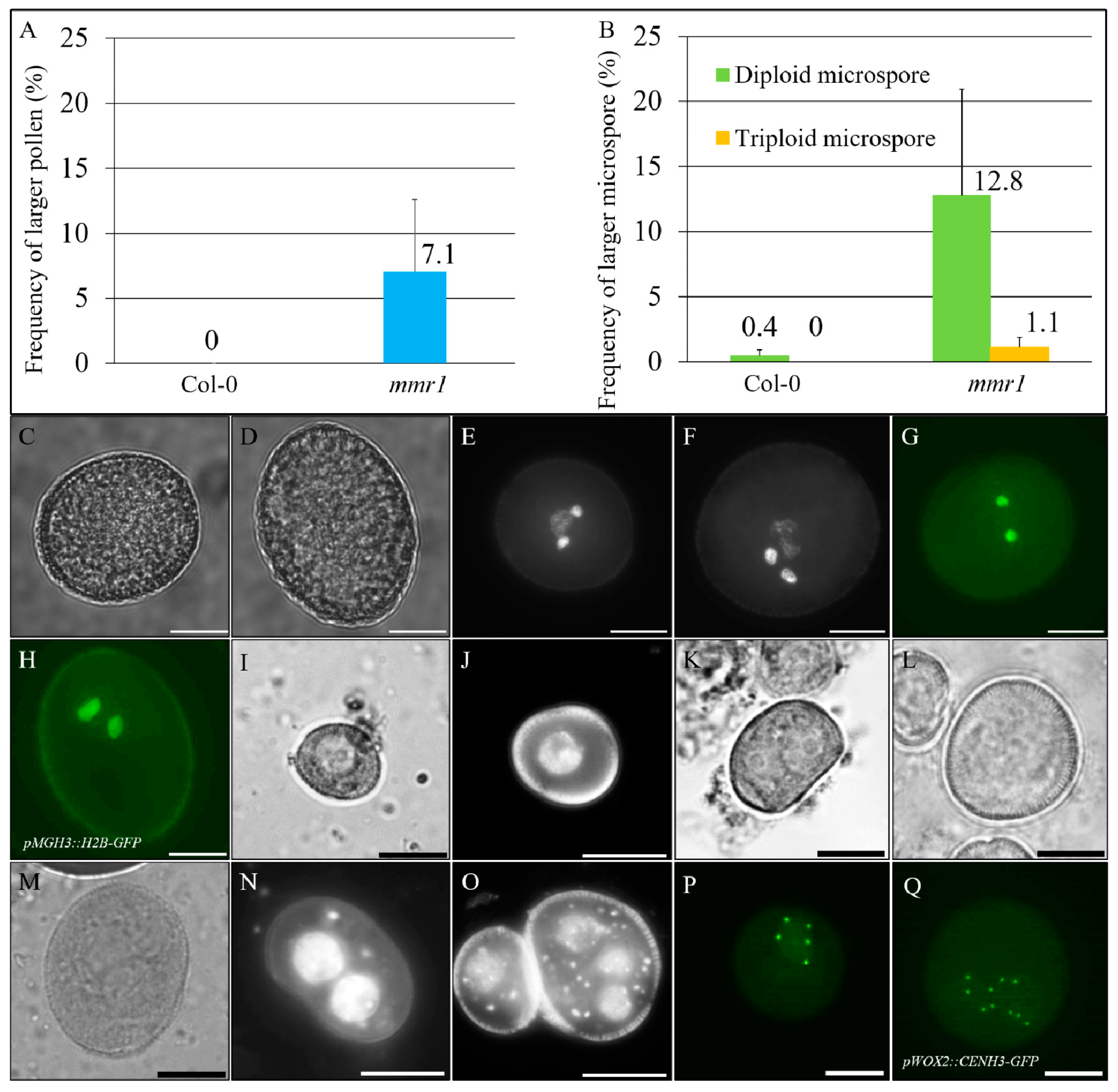
3.1. Isolation of mmr1 That Produces Diploid and/or Polyploid Pollen Grains
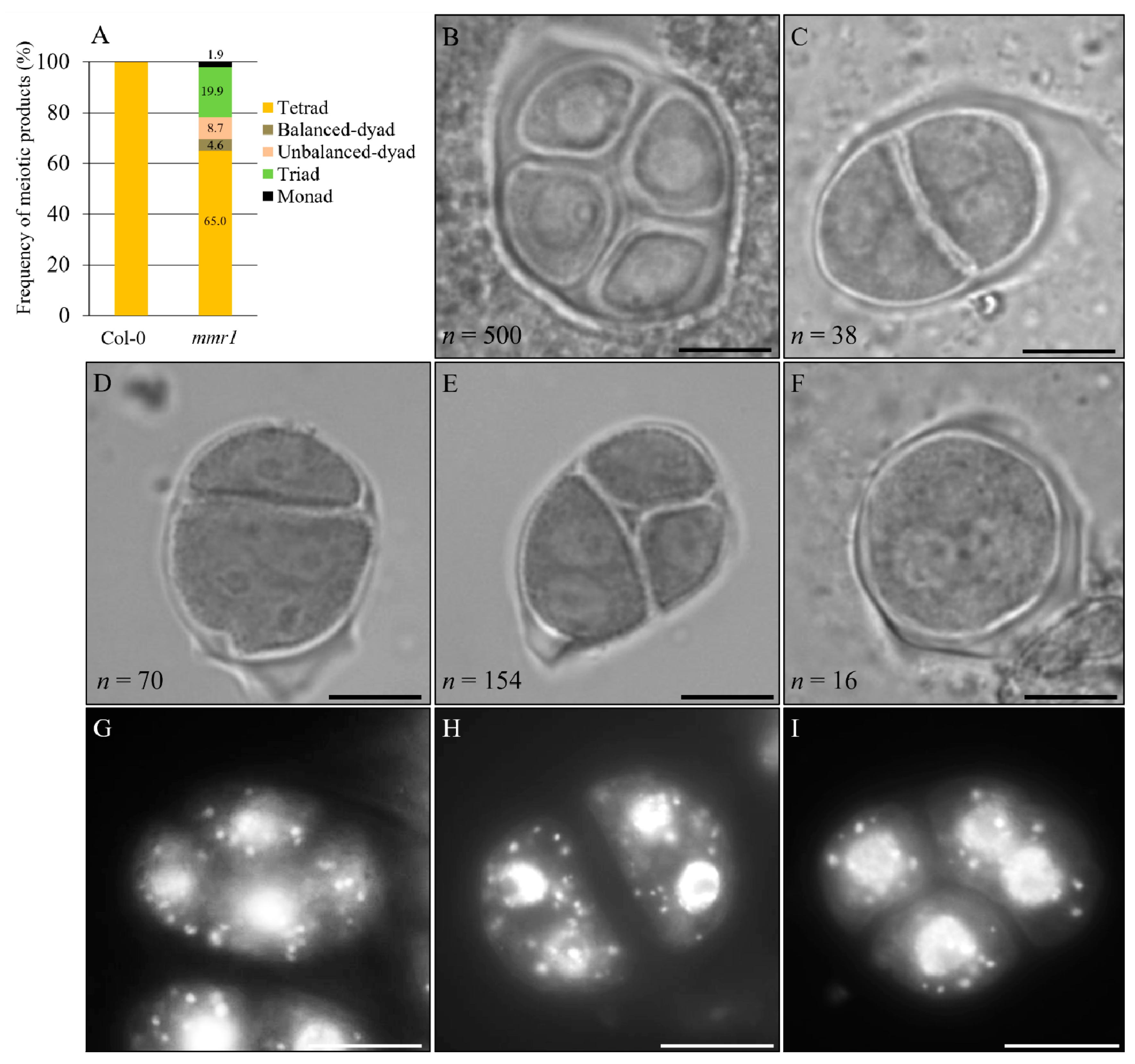
3.2. mmr1 Undergoes Meiotic Restitution
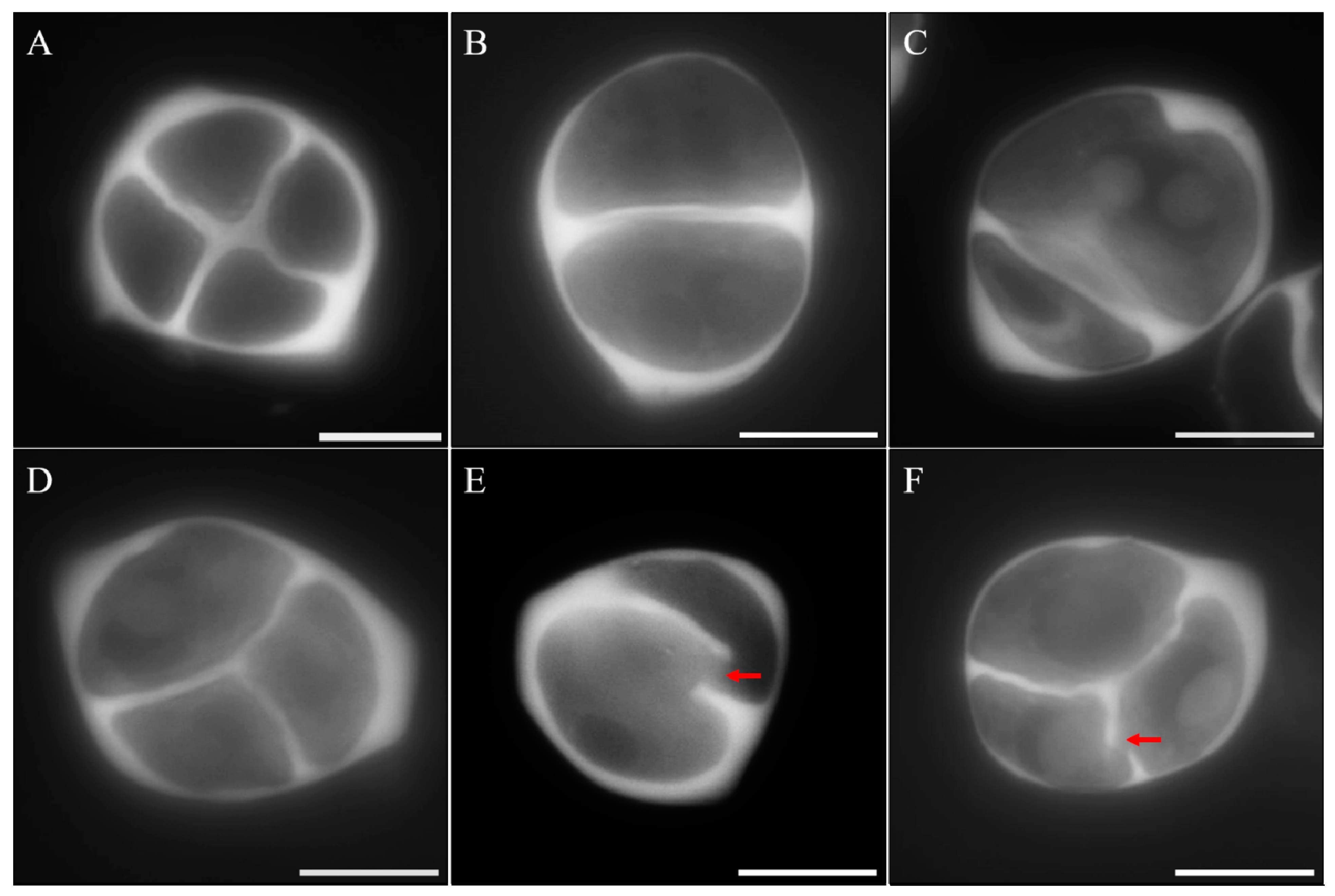
3.3. Meiotic Cytokinesis Is Defective in mmr1
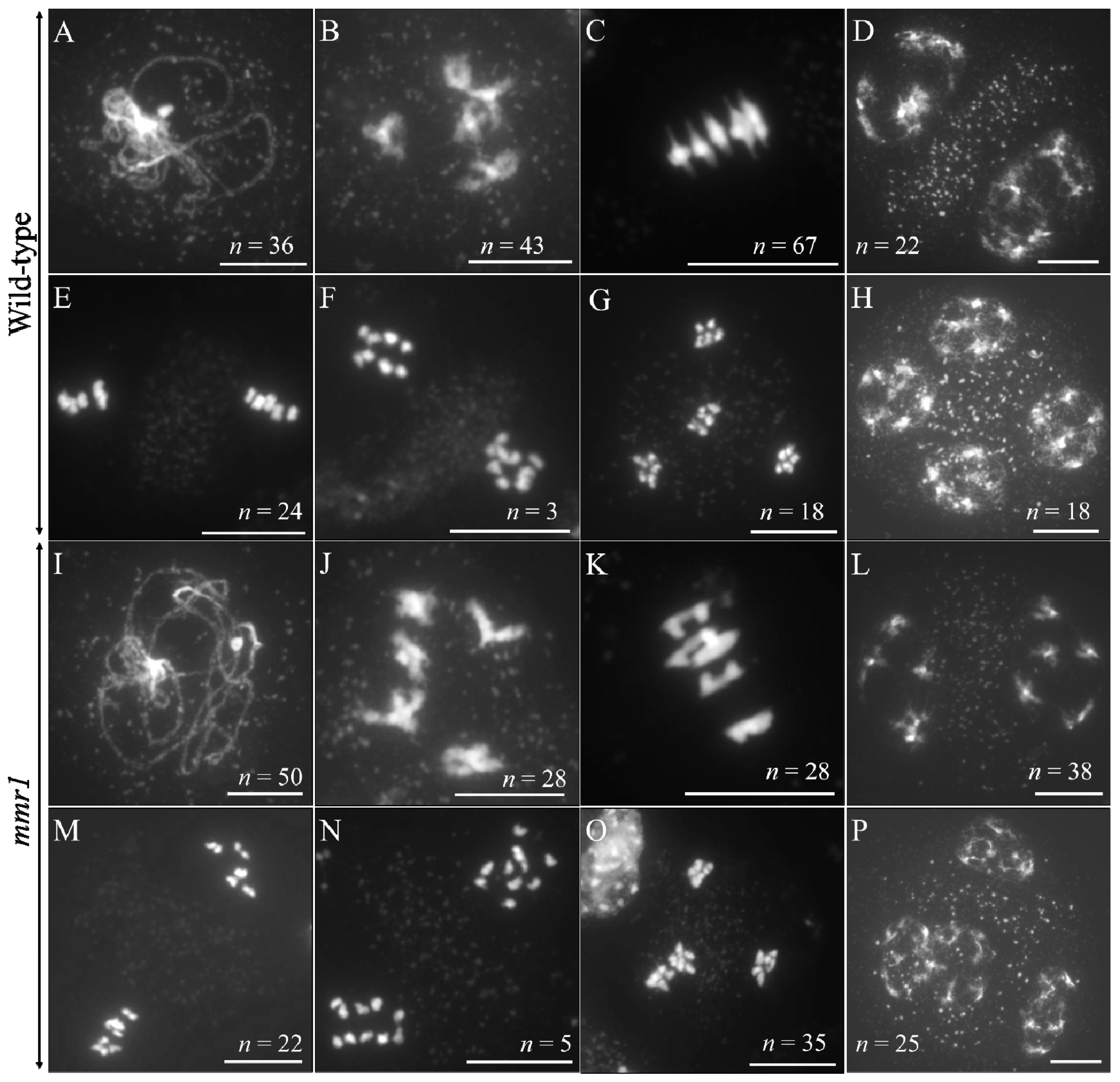
3.4. Altered Spindle Orientation and Phragmoplast Formation in mmr1
3.5. mmr1 Carries a Point Mutation in the PHD-Finger Domain of MMD1/DUET
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Del Pozo, J.C.; Ramirez-Parra, E. Whole genome duplications in plants: An overview from Arabidopsis. J. Exp. Bot. 2015, 66, 6991–7003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, R.; Wang, H.F.; Guo, C.C.; Zhang, N.; Zeng, L.P.; Chen, Y.M.; Ma, H.; Qi, J. Widespread whole genome duplications contribute to genome complexity and species diversity in angiosperms. Mol. Plant 2018, 11, 414–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bretagnolle, F.; Thompson, J.D. Gametes with the somatic chromosome number: Mechanisms of their formation and role in the evolution of autopolyploid plants. New Phytol. 1995, 129, 1–22. [Google Scholar] [CrossRef]
- Ramsey, J.; Schemske, D.W. Pathways, mechanisms, and rates of polyploid formation in flowering plants. Annu. Rev. Ecol. Syst. 1998, 29, 467–501. [Google Scholar] [CrossRef] [Green Version]
- Brownfield, L.; Kohler, C. Unreduced gamete formation in plants: Mechanisms and prospects. J. Exp. Bot. 2011, 62, 1659–1668. [Google Scholar] [CrossRef] [Green Version]
- De Storme, N.; Geelen, D. Sexual polyploidization in plants--cytological mechanisms and molecular regulation. New Phytol. 2013, 198, 670–684. [Google Scholar] [CrossRef] [Green Version]
- De Storme, N.; Geelen, D. Induction and characterization of diploid pollen grains in Arabidopsis thaliana. In Plant Meiosis: Methods and Protocols; Pradillo, M., Heckmann, S., Eds.; Springer: New York, NY, USA, 2020; pp. 331–346. [Google Scholar]
- Kreiner, J.M.; Kron, P.; Husband, B.C. Evolutionary dynamics of unreduced gametes. Trends Genet. 2017, 33, 583–593. [Google Scholar] [CrossRef]
- Loginova, D.B.; Silkova, O.G. Mechanisms of Unreduced Gamete Formation in Flowering Plants. Russ. J. Genet. 2017, 53, 741–756. [Google Scholar] [CrossRef]
- Mason, A.S.; Pires, J.C. Unreduced gametes: Meiotic mishap or evolutionary mechanism? Trends Genet. 2015, 31, 5–10. [Google Scholar] [CrossRef]
- d’Erfurth, I.; Cromer, L.; Jolivet, S.; Girard, C.; Horlow, C.; Sun, Y.; To, J.P.C.; Berchowitz, L.E.; Copenhaver, G.P.; Mercier, R. The cyclin-A CYCA1;2/TAM is required for the meiosis I to meiosis II transition and cooperates with OSD1 for the prophase to first meiotic division transition. PLoS Genet. 2010, 6, e1000989. [Google Scholar] [CrossRef] [Green Version]
- d’Erfurth, I.; Jolivet, S.; Froger, N.; Catrice, O.; Novatchkova, M.; Simon, M.; Jenczewski, E.; Mercier, R. Mutations in AtPS1 (Arabidopsis thaliana Parallel Spindle 1) Lead to the Production of Diploid Pollen Grains. PLoS Genet. 2008, 4, e1000274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Storme, N.; Geelen, D. The Arabidopsis mutant jason produces unreduced first division restitution male gametes through a parallel/fused spindle mechanism in meiosis II. Plant Physiol. 2011, 155, 1403–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hülskamp, M.; Parekh, N.S.; Grini, P.; Schneitz, K.; Zimmermann, I.; Lolle, S.J.; Pruitt, R.E. The STUD gene is required for male-specific cytokinesis after telophase II of meiosis in Arabidopsis thaliana. Dev. Biol. 1997, 187, 114–124. [Google Scholar] [CrossRef] [Green Version]
- Spielman, M.; Preuss, D.; Li, F.L.; Browne, W.E.; Scott, R.J.; Dickinson, H.G. TETRASPORE is required for male meiotic cytokinesis in Arabidopsis thaliana. Development 1997, 124, 2645–2657. [Google Scholar] [PubMed]
- Takahashi, Y.; Soyano, T.; Kosetsu, K.; Sasabe, M.; Machida, Y. HINKEL kinesin, ANP MAPKKKs and MKK6/ANQ MAPKK, which phosphorylates and activates MPK4 MAPK, constitute a pathway that is required for cytokinesis in Arabidopsis thaliana. Plant Cell Physiol. 2010, 51, 1766–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.Y.; Spielman, M.; Coles, J.P.; Li, Y.; Ghelani, S.; Bourdon, V.; Brown, R.C.; Lemmon, B.E.; Scott, R.J.; Dickinson, H.G. TETRASPORE encodes a kinesin required for male meiotic cytokinesis in Arabidopsis. Plant J. 2003, 34, 229–240. [Google Scholar] [CrossRef]
- Zeng, Q.N.; Chen, J.G.; Ellis, B.E. AtMPK4 is required for male-specific meiotic cytokinesis in Arabidopsis. Plant J. 2011, 67, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Cromer, L.; Heyman, J.; Touati, S.; Harashima, H.; Araou, E.; Girard, C.; Horlow, C.; Wassmann, K.; Schnittger, A.; De Veylder, L.; et al. OSD1 promotes meiotic progression via APC/C inhibition and forms a regulatory network with TDM and CYCA1;2/TAM. PLoS Genet. 2012, 8, e1002865. [Google Scholar] [CrossRef] [Green Version]
- Azumi, Y.; Liu, D.; Zhao, D.; Li, W.; Wang, G.; Hu, Y.; Ma, H. Homolog interaction during meiotic prophase I in Arabidopsis requires the SOLO DANCERS gene encoding a novel cyclin-like protein. EMBO J. 2002, 21, 3081–3095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Storme, N.; Copenhaver, G.P.; Geelen, D. Production of diploid male gametes in Arabidopsis by cold-induced destabilization of postmeiotic radial microtubule arrays. Plant Physiol. 2012, 160, 1808–1826. [Google Scholar] [CrossRef] [Green Version]
- Andreuzza, S.; Nishal, B.; Singh, A.; Siddiqi, I. The chromatin protein DUET/MMD1 controls expression of the meiotic gene TDM1 during male meiosis in Arabidopsis. PLoS Genet. 2015, 11, e1005396. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Niu, B.; Huang, J.; Wang, H.; Yang, X.; Dong, A.; Makaroff, C.; Ma, H.; Wang, Y. The PHD Finger Protein MMD1/DUET Ensures the Progression of Male Meiotic Chromosome Condensation and Directly Regulates the Expression of the Condensin Gene CAP-D3. Plant Cell 2016, 28, 1894–1909. [Google Scholar] [CrossRef]
- Reddy, T.V.; Kaur, J.; Agashe, B.; Sundaresan, V.; Siddiqi, I. The DUET gene is necessary for chromosome organization and progression during male meiosis in Arabidopsis and encodes a PHD finger protein. Development 2003, 130, 5975–5987. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Makaroff, C.A.; Ma, H. The Arabidopsis MALE MEIOCYTE DEATH1 gene encodes a PHD-finger protein that is required for male meiosis. Plant Cell 2003, 15, 1281–1295. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Makaroff, C.A. Arabidopsis separase AESP is essential for embryo development and the release of cohesin during meiosis. Plant Cell 2006, 18, 1213–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Shen, Y.; Cai, C.; Zhong, C.; Zhu, L.; Yuan, M.; Ren, H. The type II Arabidopsis formin14 interacts with microtubules and microfilaments to regulate cell division. Plant Cell 2010, 22, 2710–2726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.; Hong, Z.; Sivaramakrishnan, M.; Mahfouz, M.; Verma, D.P.S. Callose synthase (CalS5) is required for exine formation during microgametogenesis and for pollen viability in Arabidopsis. Plant J. 2005, 42, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Chai, M.; Yang, J.; Ning, G.; Wang, G.; Ma, H. The Arabidopsis CALLOSE DEFECTIVE MICROSPORE1 gene is required for male fertility through regulating callose metabolism during microsporogenesis. Plant Physiol. 2014, 164, 1893–1904. [Google Scholar] [CrossRef] [Green Version]
- Ravi, M.; Marimuthu, M.P.A.; Siddiqi, I. Gamete formation without meiosis in Arabidopsis. Nature 2008, 451, 1121. [Google Scholar] [CrossRef]
- Enns, L.C.; Kanaoka, M.M.; Torii, K.U.; Comai, L.; Okada, K.; Cleland, R.E. Two callose synthases, GSL1 and GSL5, play an essential and redundant role in plant and pollen development and in fertility. Plant Mol. Biol. 2005, 58, 333–349. [Google Scholar] [CrossRef]
- Brownfield, L.; Yi, J.; Jiang, H.; Minina, E.; Twell, D.; Köhler, C. Organelles maintain spindle position in plant meiosis. Nat. Commun. 2015, 6, 6492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Nadar, V.C.; Kozielski, F.; Kozlowska, M.; Yu, W.; Baas, P.W. Kinesin-12, a Mitotic Microtubule-Associated Motor Protein, Impacts Axonal Growth, Navigation, and Branching. J. Neurosci. 2010, 30, 14896–14906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.; Bourdon, V.; Dickinson, H.; Twell, D.; Park, S. Arabidopsis Fused kinase TWO-IN-ONE dominantly inhibits male meiotic cytokinesis. Plant Reprod. 2014, 27, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Kosetsu, K.; Matsunaga, S.; Nakagami, H.; Colcombet, J.; Sasabe, M.; Soyano, T.; Takahashi, Y.; Hirt, H.; Machida, Y. The MAP kinase MPK4 is required for cytokinesis in Arabidopsis thaliana. Plant Cell 2010, 22, 3778–3790. [Google Scholar] [CrossRef] [Green Version]
- Chaudhury, A.; Craig, S.; Dennis, E.; Lavithis, M.; Taylor, P.; Singh, M.; Knox, R.; Signer, E. Genetic control of male fertility in Arabidopsis thaliana: Structural analysis of premeiotic developmental mutants. Sex. Plant Reprod. 1994, 7, 17–28. [Google Scholar] [CrossRef]
- Liu, B.; De Storme, N.; Geelen, D. Gibberellin induces diploid pollen formation by interfering with meiotic cytokinesis. Plant Physiol. 2017, 173, 338–353. [Google Scholar] [CrossRef] [Green Version]
- Millar, A.A.; Gubler, F. The Arabidopsis GAMYB-like genes, MYB33 and MYB65, are microRNA-regulated genes that redundantly facilitate anther development. Plant Cell 2005, 17, 705–721. [Google Scholar] [CrossRef] [Green Version]
- d’Erfurth, I.; Jolivet, S.; Froger, N.; Catrice, O.; Novatchkova, M.; Mercier, R. Turning Meiosis into Mitosis. PLoS Biol. 2009, 7, e1000124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamariola, L.; De Storme, N.; Vannerum, K.; Vandepoele, K.; Armstrong, S.J.; Franklin, F.C.H.; Geelen, D. SHUGOSHINs and PATRONUS protect meiotic centromere cohesion in Arabidopsis thaliana. Plant J. 2014, 77, 782–794. [Google Scholar] [CrossRef]
- Yang, X.; Boateng, K.A.; Yuan, L.; Wu, S.; Baskin, T.I.; Makaroff, C.A. The Radially Swollen 4 separase mutation of Arabidopsis thaliana blocks chromosome disjunction and disrupts the radial microtubule system in meiocytes. PLoS ONE 2011, 6, e19459. [Google Scholar] [CrossRef] [Green Version]
- Zamariola, L.; De Storme, N.; Tiang, C.; Armstrong, S.J.; Franklin, F.C.H.; Geelen, D. SGO1 but not SGO2 is required for maintenance of centromere cohesion in Arabidopsis thaliana meiosis. Plant Reprod. 2013, 26, 197–208. [Google Scholar] [CrossRef] [PubMed]
- De Storme, N.; Keceli, B.N.; Zamariola, L.; Angenon, G.; Geelen, D. CENH3-GFP: A visual marker for gametophytic and somatic ploidy determination in Arabidopsis thaliana. BMC Plant Biol. 2016, 16, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; De Storme, N.; Geelen, D. Cold-induced male meiotic restitution in Arabidopsis thaliana is not mediated by GA-DELLA signaling. Front. Plant Sci. 2018, 9, 91. [Google Scholar] [CrossRef] [PubMed]
- De Storme, N.; Zamariola, L.; Mau, M.; Sharbel, T.F.; Geelen, D. Volume-based pollen size analysis: An advanced method to assess somatic and gametophytic ploidy in flowering plants. Plant Reprod. 2013, 26, 65–81. [Google Scholar] [CrossRef]
- Brownfield, L.; Hafidh, S.; Borg, M.; Sidorova, A.; Mori, T.; Twell, D. A plant germline-specific integrator of sperm specification and cell cycle progression. PLoS Genet. 2009, 5, e1000430. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yu, C.; Zhang, S.; Ye, J.; Dai, H.; Wang, H.; Huang, J.; Cao, X.; Ma, J.; Ma, H.; et al. Cell-type-dependent histone demethylase specificity promotes meiotic chromosome condensation in Arabidopsis. Nat. Plants 2020, 6, 823–837. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Gene Function | Reference |
|---|---|---|
| AESP | Proper disjunction of chromosomes | (Liu and Makaroff, 2006) [26] |
| AFH14 | Organization of microtubule and microfilament arrays | (Li et al., 2010) [27] |
| ANQ1/MKK6 | Male meiotic cytokinesis | (Takahashi et al., 2010) [16] |
| CALS5 | Callose synthase | (Dong et al., 2005) [28] |
| CDM1 | Callose metabolism | (Lu et al., 2014) [29] |
| CYCA1;2/TAM | Meiosis II cell cycle transition | (d’Erfurth et al., 2010) [11] |
| MMD1/DUET | Chromatin condensation, spindle and phragmoplast formation | (Andreuzza et al., 2015; Wang et al., 2016) [22,23] |
| DYAD/SWITCH1 | Meiotic chromosome organization | (Ravi et al., 2008) [30] |
| GSL1 and 5 | Callose wall formation | (Enns et al., 2005) [31] |
| JASON | Meiosis II spindle orientation | (Brownfield et al., 2015; De Storme and Geelen, 2011) [13,32] |
| KINESIN-12 | Polymerization of phragmoplast microtubules | (Liu et al., 2010; Oh et al., 2014) [33,34] |
| MPK4 | Male meiotic cytokinesis | (Kosetsu et al., 2010; Zeng et al., 2011) [18,35] |
| MS4 | Meiosis progression | (Chaudhury et al., 1994) [36] |
| MYB33/65 | GA signaling downstream transcription regulators | (Liu et al., 2017; Millar and Gubler, 2005) [37,38] |
| OSD1 | Meiosis cycle transition | (Cromer et al., 2012; d’Erfurth et al., 2009) [19,39] |
| PANS1 | Centromeric cohesion maintenance of sister chromatid in meiosis II | (Zamariola et al., 2014) [40] |
| AtPS1 | Meiosis II spindle orientation | (Brownfield et al., 2015; d’Erfurth et al., 2008) [12,32] |
| RGA and GAI | GA signaling repressors | (Liu et al., 2017) [37] |
| RSW4 | RMA organization | (Yang et al., 2011) [41] |
| SDS | Homolog synapsis and recombination | (Azumi et al., 2002) [20] |
| SHUGOSHIN 1/2 | Centromeric cohesion maintenance at anaphase I | (Zamariola et al., 2013; Zamariola et al., 2014) [40,42] |
| TES2 | Male meiotic cytokinesis | (Spielman et al., 1997; Yang et al., 2003a) [15,17] |
| TIO | Male meiotic cytokinesis | (Oh et al., 2014) [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.; Jin, C.; De Storme, N.; Schotte, S.; Schindfessel, C.; De Meyer, T.; Geelen, D. A Hypomorphic Mutant of PHD Domain Protein Male Meiocytes Death 1. Genes 2021, 12, 516. https://doi.org/10.3390/genes12040516
Liu B, Jin C, De Storme N, Schotte S, Schindfessel C, De Meyer T, Geelen D. A Hypomorphic Mutant of PHD Domain Protein Male Meiocytes Death 1. Genes. 2021; 12(4):516. https://doi.org/10.3390/genes12040516
Chicago/Turabian StyleLiu, Bing, Chunlian Jin, Nico De Storme, Sébastien Schotte, Cédric Schindfessel, Tim De Meyer, and Danny Geelen. 2021. "A Hypomorphic Mutant of PHD Domain Protein Male Meiocytes Death 1" Genes 12, no. 4: 516. https://doi.org/10.3390/genes12040516
APA StyleLiu, B., Jin, C., De Storme, N., Schotte, S., Schindfessel, C., De Meyer, T., & Geelen, D. (2021). A Hypomorphic Mutant of PHD Domain Protein Male Meiocytes Death 1. Genes, 12(4), 516. https://doi.org/10.3390/genes12040516