
Alternative Splicing of Pre-mRNA in the Control of Immune Activity


Abstract
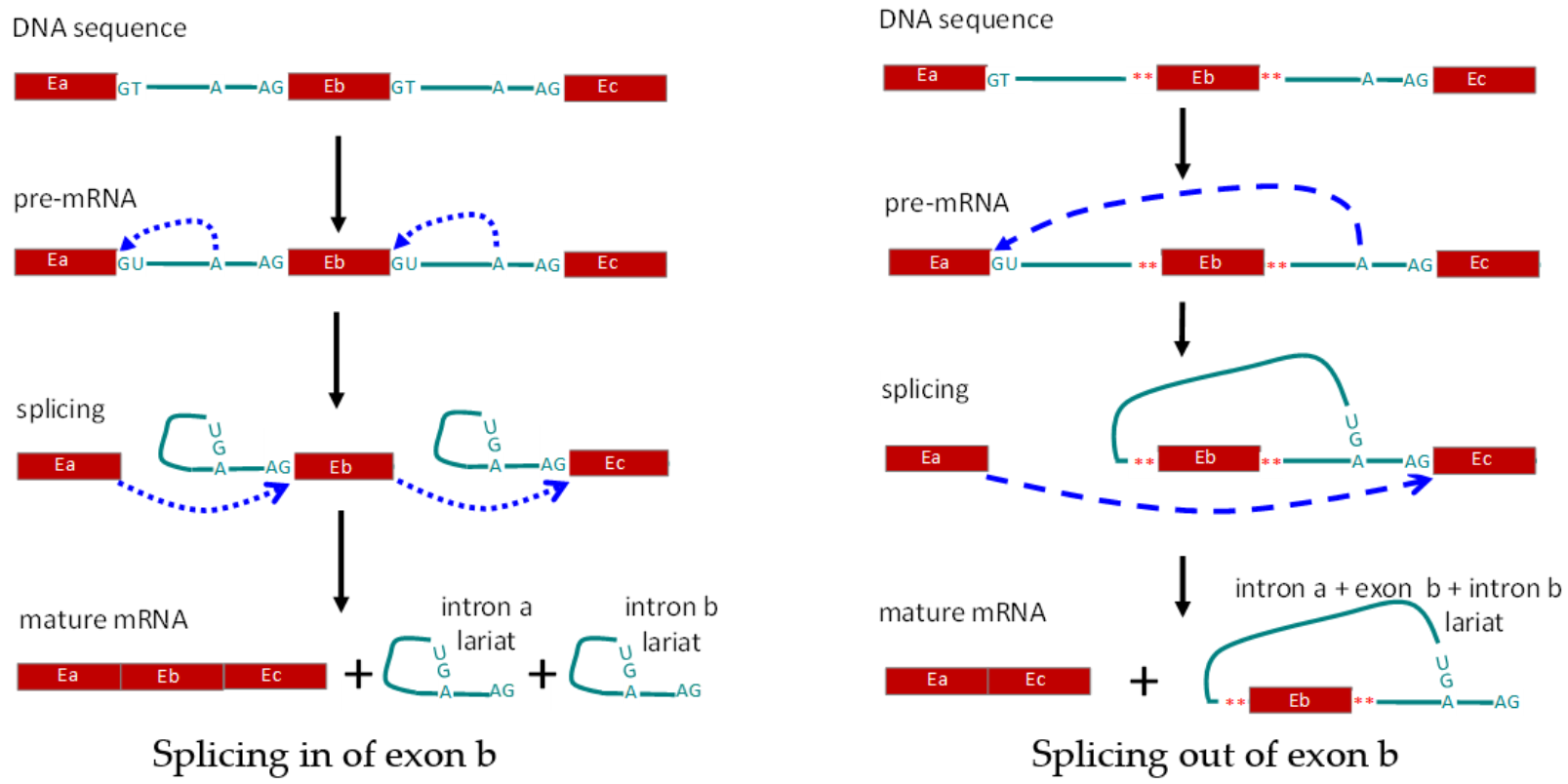
:1. Introduction
2. Influence of Alternative Splicing on Innate Immunity
3. Contribution of Alternative Splicing to Humoral Immunity
4. Control of T Cell Development and Activation Mediated by Splicing Isoforms
5. Abnormally Spliced Isoforms in Immune-Associated Diseases
6. Regulatory Mechanisms of Alternative Splicing in the Immune Response
7. Neoantigens Produced by Alternative Splicing
8. Prospect
- (1)
- Tumor cells have strategies to escape immune surveillance of neoantigens generated from the spliced variants, while for the normal somatic cells, there are also novel protein variants produced to deal with altered exogeneous surroundings or endogenous modulation. How does the immune system recognize and deal with these novel protein isoforms in the life span?
- (2)
- What are the physiological functions of the spliced isoforms? Most human genes have alternatively spliced RNA isoforms. Presently, only a small portion of them correlate with clinical disorders, such as cancer, heart disease, and immune disease, and have attracted the attention of scientific researchers, while the functions of most of them has not been identified. It is considered that spliced isoforms might play an important role through providing diversity transcripts in the long-term evolution of immune responses [141]. If without function, will these alternative exons be removed as junk sequences in the future evolution of humans?
- (3)
- If alternative splicing is correlated with clinical diseases, the next step will be how to target the abnormal alternative splicing to provide treatment. For refractory autoimmune diseases, besides the application of chemical medicines, whether the production of the alternative splicing be prevented or be targeted for degradation should be a question worthy of consideration and exploration.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mele, M.; Ferreira, P.G.; Reverter, F.; DeLuca, D.S.; Monlong, J.; Sammeth, M.; Young, T.R.; Goldmann, J.M.; Pervouchine, D.D.; Sullivan, T.J.; et al. Human genomics. The human transcriptome across tissues and individuals. Science 2015, 348, 660–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, E.T.; Sandberg, R.; Luo, S.; Khrebtukova, I.; Zhang, L.; Mayr, C.; Kingsmore, S.F.; Schroth, G.P.; Burge, C.B. Alternative isoform regulation in human tissue transcriptomes. Nature 2008, 456, 470–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.S.; Pinto, S.M.; Getnet, D.; Nirujogi, R.S.; Manda, S.S.; Chaerkady, R.; Madugundu, A.K.; Kelkar, D.S.; Isserlin, R.; Jain, S.; et al. A draft map of the human proteome. Nature 2014, 509, 575–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keren, H.; Lev-Maor, G.; Ast, G. Alternative splicing and evolution: Diversification, exon definition and function. Nat. Rev. Genet. 2010, 11, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Jiang, X.; Ditsiou, A.; Gao, Y.; Sun, J.; Lowenstein, E.D.; Huang, S.; Khaitovich, P.; Xi, J. Predominant patterns of splicing evolution on human, chimpanzee and macaque evolutionary lineages. Hum. Mol. Genet. 2018, 27, 1474–1485. [Google Scholar] [CrossRef]
- Berto, S.; Mendizabal, I.; Usui, N.; Toriumi, K.; Chatterjee, P.; Douglas, C.; Tamminga, C.A.; Preuss, T.M.; Yi, S.V.; Konopka, G. Accelerated evolution of oligodendrocytes in the human brain. Proc. Natl. Acad. Sci. USA 2019, 116, 24334–24342. [Google Scholar] [CrossRef]
- Baralle, F.E.; Giudice, J. Alternative splicing as a regulator of development and tissue identity. Nat. Rev. Mol. Cell Biol. 2017, 18, 437–451. [Google Scholar] [CrossRef]
- Bhadra, M.; Howell, P.; Dutta, S.; Heintz, C.; Mair, W.B. Alternative splicing in aging and longevity. Hum. Genet. 2020, 139, 357–369. [Google Scholar] [CrossRef]
- Rogers, T.F.; Palmer, D.H.; Wright, A.E. Sex-Specific selection drives the evolution of alternative splicing in birds. Mol. Biol. Evol. 2021, 38, 519–530. [Google Scholar] [CrossRef]
- Banerjee, A.K.; Blanco, M.R.; Bruce, E.A.; Honson, D.D.; Chen, L.M.; Chow, A.; Bhat, P.; Ollikainen, N.; Quinodoz, S.A.; Loney, C.; et al. SARS-CoV-2 disrupts splicing, translation, and protein trafficking to suppress host defenses. Cell 2020, 183, 1325–1339.e21. [Google Scholar] [CrossRef]
- Rehman, S.U.; Tabish, M. Alternative splicing of ACE2 possibly generates variants that may limit the entry of SARS-CoV-2: A potential therapeutic approach using SSOs. Clin. Sci. 2020, 134, 1143–1150. [Google Scholar] [CrossRef]
- Kim, C.H. Immune regulation by microbiome metabolites. Immunology 2018, 154, 220–229. [Google Scholar] [CrossRef]
- Bansal, A.; Henao-Mejia, J.; Simmons, R.A. Immune system: An emerging player in mediating effects of endocrine disruptors on metabolic health. Endocrinology 2018, 159, 32–45. [Google Scholar] [CrossRef]
- Muscatell, K.A. Social psychoneuroimmunology: Understanding bidirectional links between social experiences and the immune system. Brain Behav. Immun. 2020. [Google Scholar] [CrossRef]
- Ergun, A.; Doran, G.; Costello, J.C.; Paik, H.H.; Collins, J.J.; Mathis, D.; Benoist, C.; Blair, D.A.; Dustin, M.L.; Shinton, S.A.; et al. Differential splicing across immune system lineages. Proc. Natl. Acad. Sci. USA 2013, 110, 14324–14329. [Google Scholar] [CrossRef] [Green Version]
- Martinez, N.M.; Lynch, K.W. Control of alternative splicing in immune responses: Many regulators, many predictions, much still to learn. Immunol. Rev. 2013, 253, 216–236. [Google Scholar] [CrossRef]
- Yabas, M.; Elliott, H.; Hoyne, G.F. The role of alternative splicing in the control of immune homeostasis and cellular differentiation. Int. J. Mol. Sci. 2015, 17, 3. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, R.; Grosso, A.R.; Moita, L. Genome-Wide analysis of alternative splicing during dendritic cell response to a bacterial challenge. PLoS ONE 2013, 8, e61975. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Lorenzini, P.A.; Zhang, F.; Xu, S.; Wong, M.S.M.; Zheng, J.; Roca, X. Alternative splicing analysis in human monocytes and macrophages reveals MBNL1 as major regulator. Nucleic Acids Res. 2018, 46, 6069–6086. [Google Scholar] [CrossRef]
- Korbel, D.S.; Norman, P.J.; Newman, K.C.; Horowitz, A.; Gendzekhadze, K.; Parham, P.; Riley, E.M. Killer Ig-like receptor (KIR) genotype predicts the capacity of human KIR-positive CD56dim NK cells to respond to pathogen-associated signals. J. Immunol. 2009, 182, 6426–6434. [Google Scholar] [CrossRef]
- Bruijnesteijn, J.; van der Wiel, M.K.H.; de Groot, N.; Otting, N.; de Vos-Rouweler, A.J.M.; Lardy, N.M.; de Groot, N.G.; Bontrop, R.E. Extensive alternative splicing of KIR transcripts. Front. Immunol. 2018, 9, 2846. [Google Scholar] [CrossRef]
- Pizzolla, A.; Smith, J.M.; Brooks, A.G.; Reading, P.C. Pattern recognition receptor immunomodulation of innate immunity as a strategy to limit the impact of influenza virus. J. Leukoc. Biol. 2017, 101, 851–861. [Google Scholar] [CrossRef]
- De Arras, L.; Alper, S. Limiting of the innate immune response by SF3A-dependent control of MyD88 alternative mRNA splicing. PLoS Genet. 2013, 9, e1003855. [Google Scholar] [CrossRef]
- Savan, R. Alternative splicing in innate antiviral immunity. J. Interferon Cytokine Res. 2018, 38, 317–318. [Google Scholar] [CrossRef]
- Haque, N.; Ouda, R.; Chen, C.; Ozato, K.; Hogg, J.R. ZFR coordinates crosstalk between RNA decay and transcription in innate immunity. Nat. Commun. 2018, 9, 1145. [Google Scholar] [CrossRef]
- Pozzi, B.; Bragado, L.; Mammi, P.; Torti, M.F.; Gaioli, N.; Gebhard, L.G.; Solá, M.E.G.; Vaz-Drago, R.; Iglesias, N.G.; García, C.C.; et al. Dengue virus targets RBM10 deregulating host cell splicing and innate immune response. Nucleic Acids Res. 2020, 48, 6824–6838. [Google Scholar] [CrossRef]
- Ashraf, U.; Benoit-Pilven, C.; Lacroix, V.; Navratil, V.; Naffakh, N. Advances in analyzing virus-induced alterations of host cell splicing. Trends Microbiol. 2019, 27, 268–281. [Google Scholar] [CrossRef]
- Frankiw, L.; Mann, M.; Li, G.; Joglekar, A.; Baltimore, D. Alternative splicing coupled with transcript degradation modulates OAS1g antiviral activity. RNA 2020, 26, 126–136. [Google Scholar] [CrossRef] [Green Version]
- Maki, R.; Roeder, W.; Traunecker, A.; Sidman, C.; Wabl, M.; Raschke, W.; Tonegawa, S. The role of DNA rearrangement and alternative RNA processing in the expression of immunoglobulin delta genes. Cell 1981, 24, 353–365. [Google Scholar] [CrossRef]
- Zhang, K.; Saxon, A.; Max, E.E. Two unusual forms of human immunoglobulin E encoded by alternative RNA splicing of epsilon heavy chain membrane exons. J. Exp. Med. 1992, 176, 233–343. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Munoz, M.D.; Bell, S.E.; Fairfax, K.; Monzon-Casanova, E.; Cunningham, A.F.; Gonzalez-Porta, M.; Andrews, S.R.; Bunik, V.I.; Zarnack, K.; Curk, T.; et al. The RNA-binding protein HuR is essential for the B cell antibody response. Nat. Immunol. 2015, 16, 415–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilarski, L.M.; Gillitzer, R.; Zola, H.; Shortman, K.; Scollay, R. Definition of the thymic generative lineage by selective expression of high molecular weight isoforms of CD45 (T200). Eur. J. Immunol. 1989, 19, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Chui, D.; Ong, C.J.; Johnson, P.; Teh, H.S.; Marth, J.D. Specific CD45 isoforms differentially regulate T cell receptor signaling. EMBO J. 1994, 13, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Fukuhara, K.; Okumura, M.; Shiono, H.; Inoue, M.; Kadota, Y.; Miyoshi, S.; Matsuda, H. A study on CD45 isoform expression during T-cell development and selection events in the human thymus. Hum. Immunol. 2002, 63, 394–404. [Google Scholar] [CrossRef]
- Hermiston, M.L.; Xu, Z.; Weiss, A. CD45: A critical regulator of signaling thresholds in immune cells. Annu. Rev. Immunol. 2003, 21, 107–137. [Google Scholar] [CrossRef]
- Andersen, M.H.; Schrama, D.; Thor Straten, P.; Becker, J.C. Cytotoxic T cells. J. Investig. Dermatol. 2006, 126, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Zikherman, J.; Weiss, A. Alternative splicing of CD45: The tip of the iceberg. Immunity 2008, 29, 839–841. [Google Scholar] [CrossRef] [Green Version]
- Cordoba, S.P.; Choudhuri, K.; Zhang, H.; Bridge, M.; Basat, A.B.; Dustin, M.L.; Van Der Merwe, P.A. The large ectodomains of CD45 and CD148 regulate their segregation from and inhibition of ligated T-cell receptor. Blood 2013, 121, 4295–4302. [Google Scholar] [CrossRef] [Green Version]
- Chang, V.T.; Fernandes, R.A.; Ganzinger, K.A.; Lee, S.F.; Siebold, C.; McColl, J.; Jönsson, P.; Palayret, M.; Harlos, K.; Coles, C.H.; et al. Initiation of T cell signaling by CD45 segregation at ‘close contacts’. Nat. Immunol. 2016, 17, 574–582. [Google Scholar] [CrossRef] [Green Version]
- Carbone, C.B.; Kern, N.; Fernandes, R.A.; Hui, E.; Su, X.; Garcia, K.C.; Vale, R.D. In vitro reconstitution of T cell receptor-mediated segregation of the CD45 phosphatase. Proc. Natl. Acad. Sci. USA 2017, 114, E9338–E9345. [Google Scholar] [CrossRef] [Green Version]
- Razvag, Y.; Neve-Oz, Y.; Sajman, J.; Reches, M.; Sherman, E. Nanoscale kinetic segregation of TCR and CD45 in engaged microvilli facilitates early T cell activation. Nat. Commun. 2018, 9, 732. [Google Scholar] [CrossRef] [Green Version]
- Courtney, A.H.; Shvets, A.A.; Lu, W.; Griffante, G.; Mollenauer, M.; Horkova, V.; Lo, W.L.; Yu, S.; Stepanek, O.; Chakraborty, A.K.; et al. CD45 functions as a signaling gatekeeper in T cells. Sci. Signal. 2019, 12, eaaw8151. [Google Scholar] [CrossRef]
- Orta-Mascaro, M.; Consuegra-Fernandez, M.; Carreras, E.; Roncagalli, R.; Carreras-Sureda, A.; Alvarez, P.; Girard, L.; Simões, I.; Martínez-Florensa, M.; Aranda, F.; et al. CD6 modulates thymocyte selection and peripheral T cell homeostasis. J. Exp. Med. 2016, 213, 1387–1397. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.F.; Oliveira, L.; Carmo, A.M. Tuning T cell activation: The function of CD6 at the immunological synapse and in T cell responses. Curr. Drug Targets 2016, 17, 630–639. [Google Scholar] [CrossRef]
- Goncalves, C.M.; Henriques, S.N.; Santos, R.F.; Carmo, A.M. CD6, a Rheostat-Type signalosome that Tunes T cell activation. Front. Immunol. 2018, 9, 2994. [Google Scholar] [CrossRef]
- Castro, M.A.; Oliveira, M.I.; Nunes, R.J.; Fabre, S.; Barbosa, R.; Peixoto, A.; Brown, M.H.; Parnes, J.R.; Bismuth, G.; Moreira, A.; et al. Extracellular isoforms of CD6 generated by alternative splicing regulate targeting of CD6 to the immunological synapse. J. Immunol. 2007, 178, 4351–4361. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.F.; Oliveira, L.; Brown, M.H.; Carmo, A.M. Domain-specific CD6 monoclonal antibodies identify CD6 isoforms generated by alternative-splicing. Immunology 2019, 157, 296–303. [Google Scholar]
- Robinson, W.H.; Neuman de Vegvar, H.E.; Prohaska, S.S.; Rhee, J.W.; Parnes, J.R. Human CD6 possesses a large, alternatively spliced cytoplasmic domain. Eur. J. Immunol. 1995, 25, 2765–2769. [Google Scholar] [CrossRef]
- Kureel, A.K.; Kumari, S.; Saini, S.; Satyaprakash Singh, B.; Rai, A.K. Identification of a novel transcript variant of the human CD6 gene that lacks exon 9. Immunobiology 2019, 224, 666–671. [Google Scholar] [CrossRef]
- Bowen, M.A.; Whitney, G.S.; Neubauer, M.; Starling, G.C.; Palmer, D.; Zhang, J.; Nowak, N.J.; Shows, T.B.; Aruffo, A. Structure and chromosomal location of the human CD6 gene: Detection of five human CD6 isoforms. J. Immunol. 1997, 158, 1149–1156. [Google Scholar]
- Castro, M.A.; Nunes, R.J.; Oliveira, M.I.; Tavares, P.A.; Simoes, C.; Parnes, J.R.; Moreira, A.; Carmo, A.M. OX52 is the rat homologue of CD6: Evidence for an effector function in the regulation of CD5 phosphorylation. J. Leukoc. Biol. 2003, 73, 183–190. [Google Scholar] [CrossRef]
- Nishimura, H.; Yajima, T.; Naiki, Y.; Tsunobuchi, H.; Umemura, M.; Itano, K.; Matsuguchi, T.; Suzuki, M.; Ohashi, P.S.; Yoshikai, Y. Differential roles of interleukin 15 mRNA isoforms generated by alternative splicing in immune responses in vivo. J. Exp. Med. 2000, 191, 157–170. [Google Scholar] [CrossRef]
- Vudattu, N.K.; Magalhaes, I.; Hoehn, H.; Pan, D.; Maeurer, M.J. Expression analysis and functional activity of interleukin-7 splice variants. Genes Immun. 2009, 10, 132–140. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Zhang, S.; Luo, H.; Zhang, X.; Geng, G.; Li, J.; Guo, X.; Cai, W.; Li, L.; Liu, C.; et al. Interleukin 7 up-regulates CD95 protein on CD4+ T cells by affecting mRNA alternative splicing: Priming for a synergistic effect on HIV-1 reservoir maintenance. J. Biol. Chem. 2015, 290, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Bose, D.; Neumann, A.; Timmermann, B.; Meinke, S.; Heyd, F. Differential Interleukin-2 transcription kinetics render mouse but not human T cells vulnerable to splicing inhibition early after activation. Mol. Cell. Biol. 2019, 39, 39. [Google Scholar] [CrossRef] [Green Version]
- Drake, L.Y.; Kita, H. IL-33: Biological properties, functions, and roles in airway disease. Immunol. Rev. 2017, 278, 173–184. [Google Scholar] [CrossRef]
- Gordon, E.D.; Simpson, L.J.; Rios, C.L.; Ringel, L.; Lachowicz-Scroggins, M.E.; Peters, M.C.; Wesolowska-Andersen, A.; Gonzalez, J.R.; MacLeod, H.J.; Christian, L.S.; et al. Alternative splicing of interleukin-33 and type 2 inflammation in asthma. Proc. Natl. Acad. Sci. USA 2016, 113, 8765–8770. [Google Scholar] [CrossRef] [Green Version]
- Silva-Neta, H.L.; Brelaz-de-Castro, M.C.A.; Chagas, M.B.O.; Mariz, H.A.; de Arruda, R.G.; de Vasconcelos, V.F.; Pereira, M.C.; Romano, A.; Pitta, I.R.; Marques, C.D.L.; et al. CD4(+)CD45RA(-)FOXP3(low) Regulatory T Cells as potential biomarkers of disease activity in systemic lupus erythematosus brazilian patients. BioMed Res. Int. 2018, 2018, 3419565. [Google Scholar] [CrossRef] [Green Version]
- De Jager, P.L.; International MS Genetics Consortium; Jia, X.; Wang, J.; de Bakker, P.I.; Ottoboni, L.; Aggarwal, N.T.; Piccio, L.; Raychaudhuri, S.; Tran, D.; et al. Meta-Analysis of genome scans and replication identify CD6, IRF8 and TNFRSF1A as new multiple sclerosis susceptibility loci. Nat. Genet. 2009, 41, 776–782. [Google Scholar] [CrossRef] [Green Version]
- Swaminathan, B.; Matesanz, F.; Cavanillas, M.L.; Alloza, I.; Otaegui, D.; Olascoaga, J.; Cenit, M.C.; Heras, V.D.L.; Barcina, M.G.; Arroyo, R.; et al. Validation of the CD6 and TNFRSF1A loci as risk factors for multiple sclerosis in Spain. J. Neuroimmunol. 2010, 223, 100–103. [Google Scholar] [CrossRef]
- International Multiple Sclerosis Genetics Consortium. The genetic association of variants in CD6, TNFRSF1A and IRF8 to multiple sclerosis: A multicenter case-control study. PLoS ONE 2011, 6, e18813. [Google Scholar]
- Li, Y.; Singer, N.G.; Whitbred, J.; Bowen, M.A.; Fox, D.A.; Lin, F. CD6 as a potential target for treating multiple sclerosis. Proc. Natl. Acad. Sci. USA 2017, 114, 2687–2692. [Google Scholar] [CrossRef] [Green Version]
- Consuegra-Fernandez, M.; Isamat, M.; Lozano, F. Commentary: CD6 as a potential target for treating multiple sclerosis. Front. Immunol. 2017, 8, 1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, M.B.; Goerg, S.; Shen, L.; Prodeus, A.P.; Goodnow, C.C.; Kelsoe, G.; Carroll, M.C. Dependence of germinal center B cells on expression of CD21/CD35 for survival. Science 1998, 280, 582–585. [Google Scholar] [CrossRef] [PubMed]
- Kozono, Y.; Abe, R.; Kozono, H.; Kelly, R.G.; Azuma, T.; Holers, V.M. Cross-linking CD21/CD35 or CD19 increases both B7-1 and B7-2 expression on murine splenic B cells. J. Immunol. 1998, 160, 1565–1572. [Google Scholar]
- Liu, Y.J.; Xu, J.; de Bouteiller, O.; Parham, C.L.; Grouard, G.; Djossou, O.; De Saint-Vis, B.; Lebecque, S.; Banchereau, J.; Moore, K.W. Follicular dendritic cells specifically express the long CR2/CD21 isoform. J. Exp. Med. 1997, 185, 165–170. [Google Scholar] [CrossRef]
- Ogawa, S.; Yamaguchi, M.; Oka, K.; Taniguchi, M.; Ito, M.; Nishii, K.; Nakase, K.; Ohno, T.; Kita, K.; Kobayashi, T.; et al. CD21S antigen expression in tumour cells of diffuse large B-cell lymphomas is an independent prognostic factor indicating better overall survival. Br. J. Haematol. 2004, 125, 180–186. [Google Scholar] [CrossRef]
- McKelvey, K.J.; Millier, M.J.; Doyle, T.C.; Stamp, L.K.; Highton, J.; Hessian, P.A. Co-expression of CD21L and IL17A defines a subset of rheumatoid synovia, characterised by large lymphoid aggregates and high inflammation. PLoS ONE 2018, 13, e0202135. [Google Scholar] [CrossRef]
- Marzese, D.M.; Manughian-Peter, A.O.; Orozco, J.I.J.; Hoon, D.S.B. Alternative splicing and cancer metastasis: Prognostic and therapeutic applications. Clin. Exp. Metastasis 2018, 35, 393–402. [Google Scholar] [CrossRef]
- Frankiw, L.; Baltimore, D.; Li, G. Alternative mRNA splicing in cancer immunotherapy. Nat. Rev. Immunol. 2019, 19, 675–687. [Google Scholar] [CrossRef]
- Bonnal, S.C.; Lopez-Oreja, I.; Valcarcel, J. Roles and mechanisms of alternative splicing in cancer—Implications for care. Nat. Rev. Clin. Oncol. 2020, 17, 457–474. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Jiao, B.; Nie, J.; Li, X.; Wang, W.; Wang, H. The roles of alternative splicing in tumor-immune cell interactions. Curr. Cancer Drug Targets 2020, 20, 729–740. [Google Scholar] [CrossRef]
- Mishra, S.K.; Thakran, P. Intron specificity in pre-mRNA splicing. Curr. Genet. 2018, 64, 777–784. [Google Scholar] [CrossRef]
- Mishra, S.K.; Muthye, V.; Kandoi, G. Computational methods for predicting functions at the mRNA isoform level. Int. J. Mol. Sci. 2020, 21, 5686. [Google Scholar] [CrossRef]
- Fu, X.D.; Ares, M., Jr. Context-dependent control of alternative splicing by RNA-binding proteins. Nat. Rev. Genet. 2014, 15, 689–701. [Google Scholar] [CrossRef]
- Tchilian, E.Z.; Wallace, D.L.; Imami, N.; Liao, H.X.; Burton, C.; Gotch, F.; Martinson, J.; Haynes, B.F.; Beverley, P.C.L. The exon A (C77G) mutation is a common cause of abnormal CD45 splicing in humans. J. Immunol. 2001, 166, 6144–6148. [Google Scholar] [CrossRef] [Green Version]
- Dawes, R.; Petrova, S.; Liu, Z.; Wraith, D.; Beverley, P.C.; Tchilian, E.Z. Combinations of CD45 isoforms are crucial for immune function and disease. J. Immunol. 2006, 176, 3417–3425. [Google Scholar] [CrossRef] [Green Version]
- Windhagen, A.; Sonmez, D.; Hornig-Do, H.T.; Kalinowsky, A.; Schwinzer, R. Altered CD45 isoform expression in C77G carriers influences cytokine responsiveness and adhesion properties of T cells. Clin. Exp. Immunol. 2007, 150, 509–517. [Google Scholar] [CrossRef]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N(6)-methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature 2015, 518, 560–564. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Klukovich, R.; Peng, H.; Wang, Z.; Yu, T.; Zhang, Y.; Zheng, H.; Klungland, A.; Yan, W. ALKBH5-dependent m6A demethylation controls splicing and stability of long 3’-UTR mRNAs in male germ cells. Proc. Natl. Acad. Sci. USA 2018, 115, E325–E333.e8. [Google Scholar] [CrossRef] [Green Version]
- Gokhale, N.S.; McIntyre, A.B.R.; Mattocks, M.D.; Holley, C.L.; Lazear, H.M.; Mason, C.E.; Horner, S.M. Altered m(6)A modification of specific cellular transcripts affects flaviviridae infection. Mol. Cell 2020, 77, 542–555.e8. [Google Scholar] [CrossRef]
- Ule, J.; Blencowe, B.J. Alternative splicing regulatory networks: Functions, mechanisms, and evolution. Mol. Cell 2019, 76, 329–345. [Google Scholar] [CrossRef]
- Rothrock, C.R.; House, A.E.; Lynch, K.W. HnRNP L represses exon splicing via a regulated exonic splicing silencer. EMBO J. 2005, 24, 2792–2802. [Google Scholar] [CrossRef] [Green Version]
- Oberdoerffer, S.; Moita, L.F.; Neems, D.; Freitas, R.P.; Hacohen, N.; Rao, A. Regulation of CD45 alternative splicing by heterogeneous ribonucleoprotein, hnRNPLL. Science 2008, 321, 686–691. [Google Scholar] [CrossRef] [Green Version]
- Preussner, M.; Schreiner, S.; Hung, L.H.; Porstner, M.; Jack, H.M.; Benes, V.; Rätsch, G.; Bindereif, A.; Preussner, M. HnRNP L and L-like cooperate in multiple-exon regulation of CD45 alternative splicing. Nucleic Acids Res. 2012, 40, 5666–5678. [Google Scholar] [CrossRef]
- Katsuyama, T.; Li, H.; Comte, D.; Tsokos, G.C.; Moulton, V.R. Splicing factor SRSF1 controls T cell hyperactivity and systemic autoimmunity. J. Clin. Investig. 2019, 129, 5411–5423. [Google Scholar] [CrossRef] [Green Version]
- Katsuyama, T.; Martin-Delgado, I.J.; Krishfield, S.M.; Kyttaris, V.C.; Moulton, V.R. Splicing factor SRSF1 controls T cell homeostasis and its decreased levels are linked to lymphopenia in systemic lupus erythematosus. Rheumatology 2020, 59, 2146–2155. [Google Scholar] [CrossRef]
- Paz, S.; Ritchie, A.; Mauer, C.; Caputi, M. The RNA binding protein SRSF1 is a master switch of gene expression and regulation in the immune system. Cytokine Growth Factor Rev. 2021, 57, 19–26. [Google Scholar] [CrossRef] [PubMed]
- da Gloria, V.G.; Martins de Araujo, M.; Mafalda Santos, A.; Leal, R.; de Almeida, S.F.; Carmo, A.M.; Moreira, A. T cell activation regulates CD6 alternative splicing by transcription dynamics and SRSF1. J. Immunol. 2014, 193, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.J.; Luo, S.; Ho, J.X.J.; Ly, P.T.; Goh, E.; Roca, X. Characterization of the regulation of CD46 RNA alternative splicing. J. Biol. Chem. 2016, 291, 14311–14323. [Google Scholar] [CrossRef] [Green Version]
- La Porta, J.; Matus-Nicodemos, R.; Valentin-Acevedo, A.; Covey, L.R. The RNA-binding protein, Polypyrimidine Tract-Binding Protein 1 (PTBP1) is a key regulator of CD4 T cell activation. PLoS ONE 2016, 11, e0158708. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Qian, H.; Xue, Y.; Fu, X.D. PTB/nPTB: Master regulators of neuronal fate in mammals. Biophys. Rep. 2018, 4, 204–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monzon-Casanova, E.; Screen, M.; Diaz-Munoz, M.D.; Coulson, R.M.R.; Bell, S.E.; Lamers, G.; Solimena, M.; Smith, C.W.J.; Turner, M. The RNA-binding protein PTBP1 is necessary for B cell selection in germinal centers. Nat. Immunol. 2018, 19, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Sasanuma, H.; Ozawa, M.; Yoshida, N. RNA-binding protein Ptbp1 is essential for BCR-mediated antibody production. Int. Immunol. 2019, 31, 157–166. [Google Scholar] [CrossRef]
- Monzon-Casanova, E.; Matheson, L.S.; Tabbada, K.; Zarnack, K.; Smith, C.W.; Turner, M. Polypyrimidine tract-binding proteins are essential for B cell development. eLife 2020, 9, e53557. [Google Scholar] [CrossRef]
- Wang, Y.; Gogol-Doring, A.; Hu, H.; Frohler, S.; Ma, Y.; Jens, M.; Maaskola, J.; Murakawa, Y.; Quedenau, C.; Landthaler, M.; et al. Integrative analysis revealed the molecular mechanism underlying RBM10-mediated splicing regulation. EMBO Mol. Med. 2013, 5, 1431–1442. [Google Scholar] [CrossRef]
- Inoue, A.; Yamamoto, N.; Kimura, M.; Nishio, K.; Yamane, H.; Nakajima, K. RBM10 regulates alternative splicing. FEBS Lett. 2014, 588, 942–947. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, J.; Bechara, E.; Schlesinger, D.; Delgado, J.; Serrano, L.; Valcarcel, J. Tumor suppressor properties of the splicing regulatory factor RBM10. RNA Biol. 2016, 13, 466–472. [Google Scholar] [CrossRef]
- Van Nostrand, E.L.; Freese, P.; Pratt, G.A.; Wang, X.; Wei, X.; Xiao, R.; Blue, S.M.; Chen, J.-Y.; Cody, N.A.L.; Dominguez, D.; et al. A large-scale binding and functional map of human RNA-binding proteins. Nature 2020, 583, 711–719. [Google Scholar] [CrossRef]
- Shukla, S.; Oberdoerffer, S. Co-transcriptional regulation of alternative pre-mRNA splicing. Biochim. Biophys. Acta 2012, 1819, 673–683. [Google Scholar] [CrossRef] [Green Version]
- Davis-Turak, J.; Johnson, T.L.; Hoffmann, A. Mathematical modeling identifies potential gene structure determinants of co-transcriptional control of alternative pre-mRNA splicing. Nucleic Acids Res. 2018, 46, 10598–10607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, S.; Kavak, E.; Gregory, M.; Imashimizu, M.; Shutinoski, B.; Kashlev, M.; Oberdoerffer, P.; Sandberg, R.; Oberdoerffer, S. CTCF-promoted RNA polymerase II pausing links DNA methylation to splicing. Nature 2011, 479, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Jimeno-Gonzalez, S.; Payan-Bravo, L.; Munoz-Cabello, A.M.; Guijo, M.; Gutierrez, G.; Prado, F.; Reyes, J.C. Defective histone supply causes changes in RNA polymerase II elongation rate and cotranscriptional pre-mRNA splicing. Proc. Natl. Acad. Sci. USA 2015, 112, 14840–14845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, V.; Ellis, J.D.; Shen, Z.; Song, D.Y.; Pan, Q.; Watt, A.T.; Freier, S.M.; Bennett, C.F.; Sharma, A.; Bubulya, P.A.; et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Mol. Cell 2010, 39, 925–938. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Hamblin, M.H.; Yin, K.J. The long noncoding RNA Malat1: Its physiological and pathophysiological functions. RNA Biol. 2017, 14, 1705–1714. [Google Scholar] [CrossRef]
- Hewitson, J.P.; West, K.A.; James, K.R.; Rani, G.F.; Dey, N.; Romano, A.; Brown, N.; Teichmann, S.A.; Kaye, P.M.; Lagos, D. Malat1 suppresses immunity to infection through promoting expression of maf and IL-10 in Th cells. J. Immunol. 2020, 204, 2949–2960. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Wang, Z.; Liu, L.; Yang, Z.; Liu, S.; Ma, Z.; Liu, Y.; Ma, Y.; Zhang, L.; Zhang, X.; et al. LncRNA Malat1 inhibition of TDP43 cleavage suppresses IRF3-initiated antiviral innate immunity. Proc. Natl. Acad. Sci. USA 2020, 117, 23695–23706. [Google Scholar] [CrossRef]
- Gast, M.; Rauch, B.H.; Nakagawa, S.; Haghikia, A.; Jasina, A.; Haas, J.; Nath, N.; Jensen, L.; Stroux, A.; Böhm, A.; et al. Immune system-mediated atherosclerosis caused by deficiency of long non-coding RNA MALAT1 in ApoE-/- mice. Cardiovasc. Res. 2019, 115, 302–314. [Google Scholar] [CrossRef]
- Liang, Z.; Tang, F. The potency of lncRNA MALAT1/miR-155/CTLA4 axis in altering Th1/Th2 balance of asthma. Biosci. Rep. 2020, 40, 40. [Google Scholar] [CrossRef] [Green Version]
- Werner, A. Biological functions of natural antisense transcripts. BMC Biol. 2013, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Morrissy, A.S.; Griffith, M.; Marra, M.A. Extensive relationship between antisense transcription and alternative splicing in the human genome. Genome Res. 2011, 21, 1203–1212. [Google Scholar] [CrossRef] [Green Version]
- Rong, J.; Yin, J.; Su, Z. Natural antisense RNAs are involved in the regulation of CD45 expression in autoimmune diseases. Lupus 2015, 24, 235–239. [Google Scholar] [CrossRef]
- Zhang, T.; Dong, Z.; Cai, H.; Rong, J.; Su, Z. Estradiol regulates the expression of CD45 splicing isoforms in lymphocytes. Mol. Biol. Rep. 2020, 47, 3025–3030. [Google Scholar] [CrossRef]
- Dvinge, H.; Guenthoer, J.; Porter, P.L.; Bradley, R.K. RNA components of the spliceosome regulate tissue- and cancer-specific alternative splicing. Genome Res. 2019, 29, 1591–1604. [Google Scholar] [CrossRef] [Green Version]
- Pu, M.; Chen, J.; Tao, Z.; Miao, L.; Qi, X.; Wang, Y.; Ren, J. Regulatory network of miRNA on its target: Coordination between transcriptional and post-transcriptional regulation of gene expression. Cell. Mol. Life Sci. 2019, 76, 441–451. [Google Scholar] [CrossRef]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Srivastava, R.; Daulatabad, S.V.; Srivastava, M.; Janga, S.C. Role of SARS-CoV-2 in Altering the RNA-Binding protein and miRNA-Directed Post-Transcriptional regulatory networks in humans. Int. J. Mol. Sci. 2020, 21, 7090. [Google Scholar] [CrossRef]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. circRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Wilusz, J.E. A 360 degrees view of circular RNAs: From biogenesis to functions. Wiley Interdiscip. Rev. RNA 2018, 9, e1478. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.L. The expanding regulatory mechanisms and cellular functions of circular RNAs. Nat. Rev. Mol. Cell Biol. 2020, 21, 475–490. [Google Scholar] [CrossRef]
- Zhu, P.; Zhu, X.; Wu, J.; He, L.; Lu, T.; Wang, Y.; Liu, B.; Ye, B.; Sun, L.; Fan, D.; et al. IL-13 secreted by ILC2s promotes the self-renewal of intestinal stem cells through circular RNA circPan3. Nat. Immunol. 2019, 20, 183–194. [Google Scholar] [CrossRef]
- Wang, Y.H.; Yu, X.H.; Luo, S.S.; Han, H. Comprehensive circular RNA profiling reveals that circular RNA100783 is involved in chronic CD28-associated CD8(+)T cell ageing. Immun. Ageing 2015, 12, 17. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, C.X.; Xue, W.; Zhang, Y.; Jiang, S.; Yin, Q.F.; Wei, J.; Yao, R.-W.; Yang, L.; Chen, L.-L. Coordinated circRNA Biogenesis and Function with NF90/NF110 in Viral Infection. Mol. Cell 2017, 67, 214–227.e7. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.X.; Li, X.; Nan, F.; Jiang, S.; Gao, X.; Guo, S.K.; Xue, W.; Cui, Y.; Dong, K.; Ding, H.; et al. Structure and degradation of circular RNAs regulate PKR activation in innate immunity. Cell 2019, 177, 865–880.e21. [Google Scholar] [CrossRef]
- Aartsma-Rus, A. Antisense-mediated modulation of splicing: Therapeutic implications for Duchenne muscular dystrophy. RNA Biol. 2010, 7, 453–461. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, A. Sense in antisense therapy for spinal muscular atrophy. N. Engl. J. Med. 2012, 366, 761–763. [Google Scholar] [CrossRef]
- Arechavala-Gomeza, V.; Anthony, K.; Morgan, J.; Muntoni, F. Antisense oligonucleotide-mediated exon skipping for Duchenne muscular dystrophy: Progress and challenges. Curr. Gene. Ther. 2012, 12, 152–160. [Google Scholar] [CrossRef]
- Slansky, J.E.; Spellman, P.T. Alternative splicing in tumors—A path to immunogenicity? N. Engl. J. Med. 2019, 380, 877–880. [Google Scholar] [CrossRef]
- Smith, C.C.; Selitsky, S.R.; Chai, S.; Armistead, P.M.; Vincent, B.G.; Serody, J.S. Alternative tumour-specific antigens. Nat. Rev. Cancer 2019, 19, 465–478. [Google Scholar] [CrossRef]
- Shen, S.; Park, J.W.; Lu, Z.X.; Lin, L.; Henry, M.D.; Wu, Y.N.; Zhou, Q.; Xing, Y. rMATS: Robust and flexible detection of differential alternative splicing from replicate RNA-Seq data. Proc. Natl. Acad. Sci. USA 2014, 111, E5593–E5601. [Google Scholar] [CrossRef] [Green Version]
- Denti, L.; Rizzi, R.; Beretta, S.; Vedova, G.D.; Previtali, M.; Bonizzoni, P. ASGAL: Aligning RNA-Seq data to a splicing graph to detect novel alternative splicing events. BMC Bioinform. 2018, 19, 444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhou, C.; Tang, L.; Gong, Y.; Wei, Z.; Zhang, G.; Wang, F.; Liu, Q.; Yu, J. ASNEO: Identification of personalized alternative splicing based neoantigens with RNA-seq. Aging 2020, 12, 14633–146348. [Google Scholar] [CrossRef] [PubMed]
- Oka, M.; Xu, L.; Suzuki, T.; Yoshikawa, T.; Sakamoto, H.; Uemura, H.; Yoshizawa, A.C.; Suzuki, Y.; Nakatsura, T.; Ishihama, Y.; et al. Aberrant splicing isoforms detected by full-length transcriptome sequencing as transcripts of potential neoantigens in non-small cell lung cancer. Genome Biol. 2021, 22, 9. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Moreno, J.F.; Romao, L. Perspective in alternative splicing coupled to nonsense-mediated mRNA decay. Int. J. Mol. Sci. 2020, 21, 9424. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Plank, T.D.; Su, F.; Shi, X.; Liu, C.; Ji, Y.; Li, S.; Huynh, A.; Shi, C.; Zhu, B.; et al. The nonsense-mediated RNA decay pathway is disrupted in inflammatory myofibroblastic tumors. J. Clin. Investig. 2016, 126, 3058–3062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popp, M.W.; Maquat, L.E. Nonsense-mediated mRNA decay and cancer. Curr. Opin. Genet. Dev. 2018, 48, 44–50. [Google Scholar] [CrossRef]
- Perumal, D.; Imai, N.; Lagana, A.; Finnigan, J.; Melnekoff, D.; Leshchenko, V.V.; Solovyov, A.; Madduri, D.; Chari, A.; Cho, H.J.; et al. Mutation-derived neoantigen-specific T-cell responses in multiple myeloma. Clin. Cancer Res. 2020, 26, 450–464. [Google Scholar] [CrossRef] [Green Version]
- Mortezaee, K. Immune escape: A critical hallmark in solid tumors. Life Sci. 2020, 258, 118110. [Google Scholar] [CrossRef]
- Hayashi, H.; Nakagawa, K. Combination therapy with PD-1 or PD-L1 inhibitors for cancer. Int. J. Clin. Oncol. 2020, 25, 818–830. [Google Scholar] [CrossRef]
- De Miguel, M.; Calvo, E. Clinical challenges of immune checkpoint inhibitors. Cancer Cell 2020, 38, 326–333. [Google Scholar] [CrossRef]
- Rotival, M.; Quach, H.; Quintana-Murci, L. Defining the genetic and evolutionary architecture of alternative splicing in response to infection. Nat. Commun. 2019, 10, 1671. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regulator | Acting Way | Target Gene | Associated Function | Ref. |
|---|---|---|---|---|
| DNA mutation—C77G | Cis | CD45 | Mutation in DNA sequence of CD45 exon 4 alters the splicing of the exon. | [78] |
| RNA editing—M6A | Cis | CIRBP | Viral infection modulates alternative splicing of CIRBP through RNA modification. | [81] |
| RBP—hnRNP LL | Trans | CD45 | HnRNP LL protein binds to CD45 transcript to control the expression of CD45RA and RO. | [84] |
| RBP—SRSF1 | Trans | CD46 | SRSF1 protein promotes the exclusion of exon 13 in CD46. | [90] |
| RBP—PTBP1 | Trans | CD46 | PTBP1 protein promotes the inclusion of exon 13 in CD46. | [90] |
| RBP—RBM10 | Trans | SAT1 | RBM10 is responsible for SAT1 exon 4 skipping for limiting viral replication. | [26] |
| lncRNA—MALAT1 | Trans | SAT1 | MALAT1 regulates the splicing of multiple genes including SAT1 by interaction with serine/arginine splicing factors. | [104] |
| sncRNA—U1 RNA | Trans | CD46 | U1 small nuclear RNA defines exons 7 and 8 of CD46. | [90] |
| circRNA—circPan3 | Trans | Pan | circPan3, originated from the back-splicing of Pan3 transcript, increases the expression of IL-13. | [121] |
| DNA modification | co-transcriptional | CD45 | DNA binding protein, CTCF, linking DNA methylation to modulate the splicing of CD45 through controlling the RNA polymerase II elongation rate. | [102] |
| Histone modification | co-transcriptional | CD44 | Histone modification modulates the alternative splicing of CD44 through controlling the RNA polymerase II elongation rate. | [103] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Z.; Huang, D. Alternative Splicing of Pre-mRNA in the Control of Immune Activity. Genes 2021, 12, 574. https://doi.org/10.3390/genes12040574
Su Z, Huang D. Alternative Splicing of Pre-mRNA in the Control of Immune Activity. Genes. 2021; 12(4):574. https://doi.org/10.3390/genes12040574
Chicago/Turabian StyleSu, Zhongjing, and Dongyang Huang. 2021. "Alternative Splicing of Pre-mRNA in the Control of Immune Activity" Genes 12, no. 4: 574. https://doi.org/10.3390/genes12040574
APA StyleSu, Z., & Huang, D. (2021). Alternative Splicing of Pre-mRNA in the Control of Immune Activity. Genes, 12(4), 574. https://doi.org/10.3390/genes12040574