
Identification of Novel Loci and Candidate Genes for Resistance to Powdery Mildew in a Resequenced Cucumber Germplasm

Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resistance Donor | Mapping Population | Gene/QTL | Chromosome | Causal Gene | Reference |
|---|---|---|---|---|---|
| Puerto Rico 37 | - | - | - | - | [14] |
| Indian accession PI197087 | - | - | - | - | [15] |
| Yomaki | - | - | - | - | [13] |
| Natsufushinari | - | - | - | - | [3] |
| Indian accession PI 197088-1 | RILs of PI 197088-1 × Santou | - | 1, 5, 6, 7 | [12] | |
| S94 | RILs (F6:7 family) of Inbred line S94 (R) × S06(S) | pm1.1, pm2.1, pm4.1, pm6.1 | 1, 2, 4, 6 | [9] | |
| WIS2757 | F2 family of WIS2757 × 19032 | - | 5 | [20] | |
| K8 | F2 and F2:3 families of K8 (R) × K18 (S) | pm5.1, pm5.2, pm5.3, pm6.1 | 5, 6 | - | [11] |
| WI 2757 | F2:3 families of WI2757 (R) × True lemon (S) | pm1.1, pm1.2, pm3.1, pm4.1, pm 5.1, pm5.2 | 1, 3, 4, 5 | - | [17] |
| CS-PMR1 | RILs of CS-PMR1 (derived from PI 197088, R) × Santou (intermediate S) | 9 QTLs | All chromosomes except 7 | - | [22] |
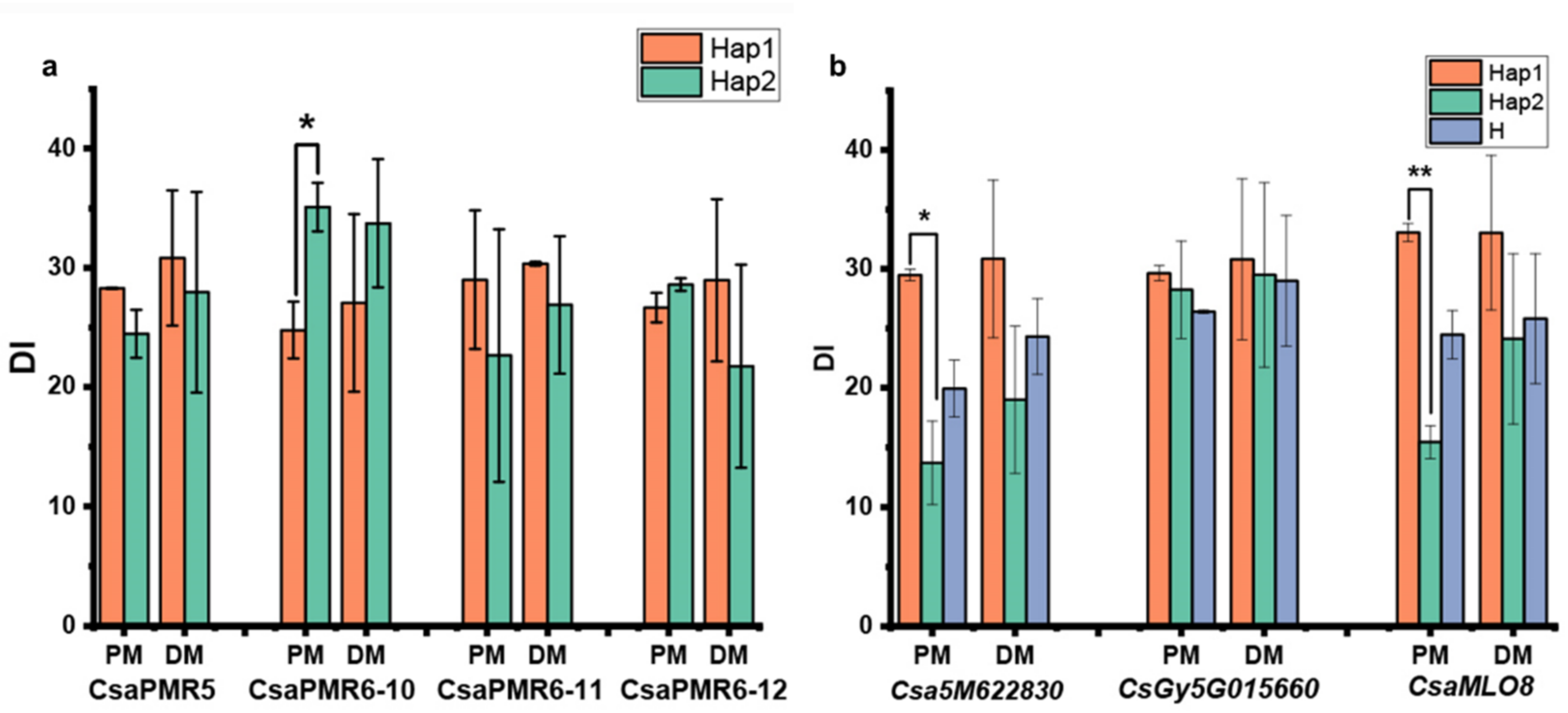
| S1003 | BC3F1 and BC2F2 families of S1003(R) × S1001(S) | pm5.1 | 5 | CsaMLO8 (CsMLO1) | [18,23] |
| Jin5-508 | F2 and CSILs families of Jin5-508(R) × D8(S) | pm1.1 | 1 | [24] | |
| NCG122 | F2 families of NCG122 (R) × NCG121(S) | pm-s | 5 | [10] | |
| PI 197088 | RIL families of PI 19,788 (R) × Coolgreen (S) | pm1.1, pm2.1, pm5.1, pm 6.1 | 1, 2, 5, 6 | [21] | |
| IL52 | RIL families of IL52 (R) × changchunmici (S) | pm | 5 | Csa5M622830 | [25] |
| PM-R | F2 families of PM-R × PM-S | pm5.2, pm6.1 | 5, 6 | CsGy5G015660 | [16] |
2. Materials and Methods
2.1. Plant Materials
2.2. Disease Tests and Disease Index Calculation
2.3. Genetic Diversity of PM Resistance in the CG Population
2.4. Genome-Wide Association Analyses of PM Resistance
2.5. Linkage-Disequilibrium (LD) Analysis
2.6. Candidate Gene Analysis
3. Results
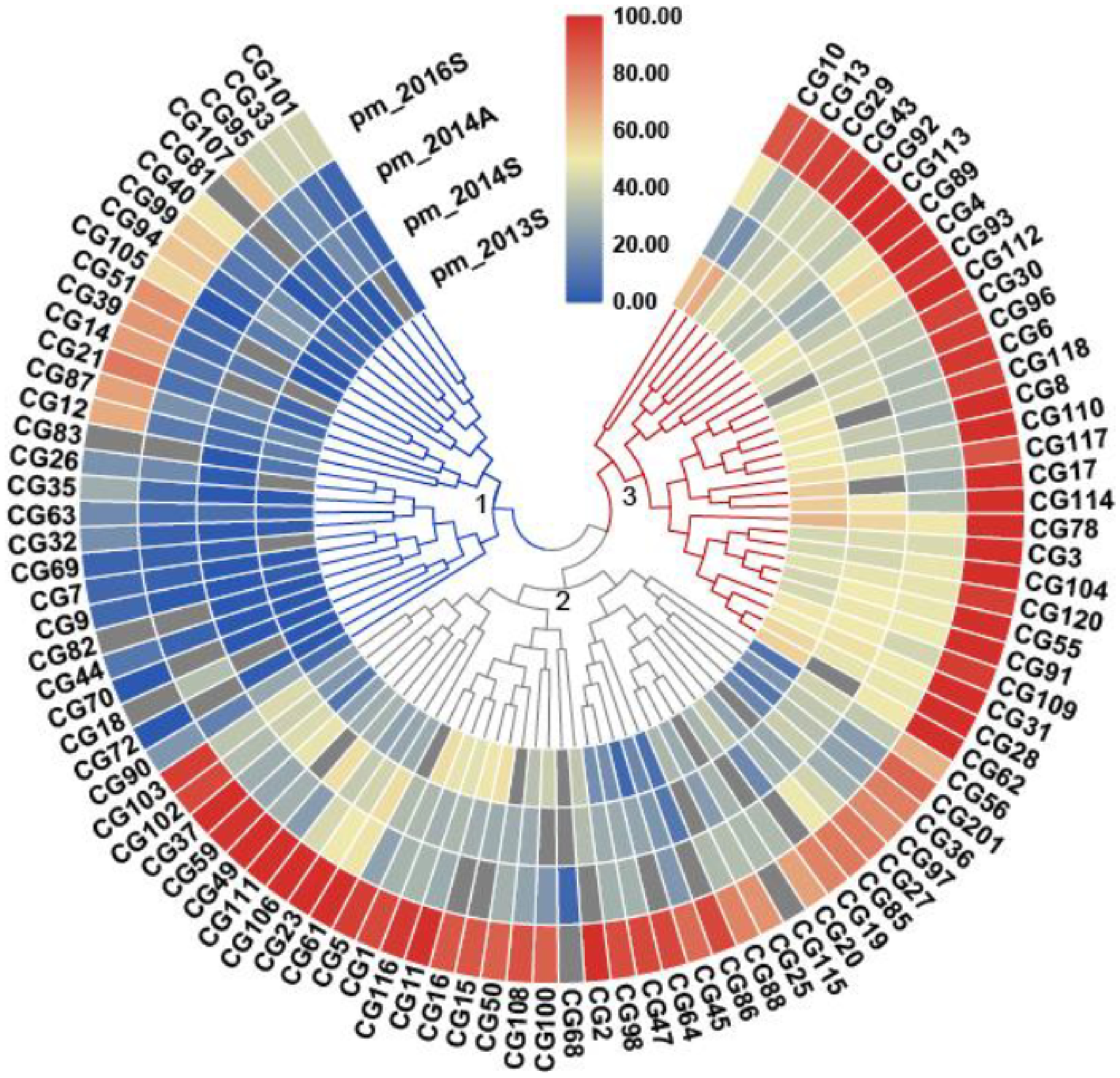
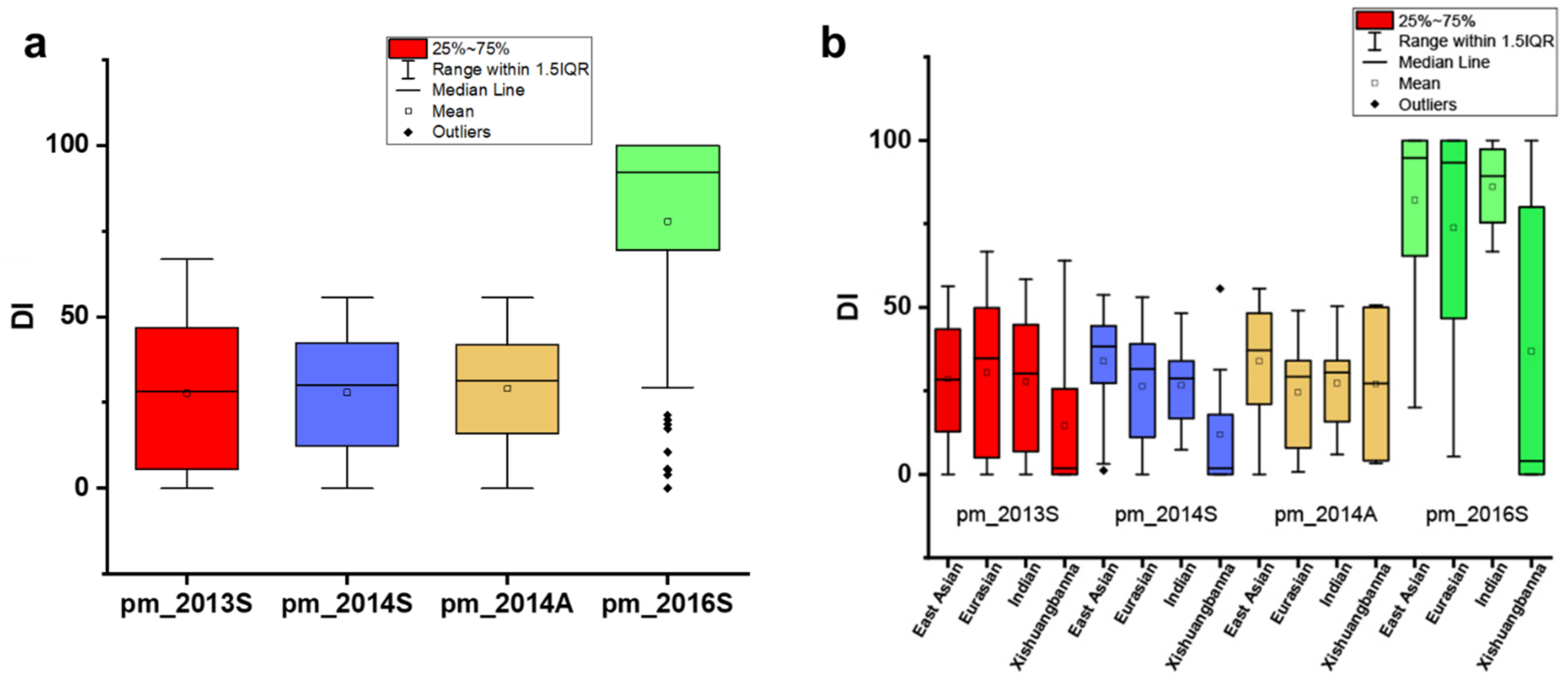
3.1. Genetic Diversity of PM Resistance in the CG Population
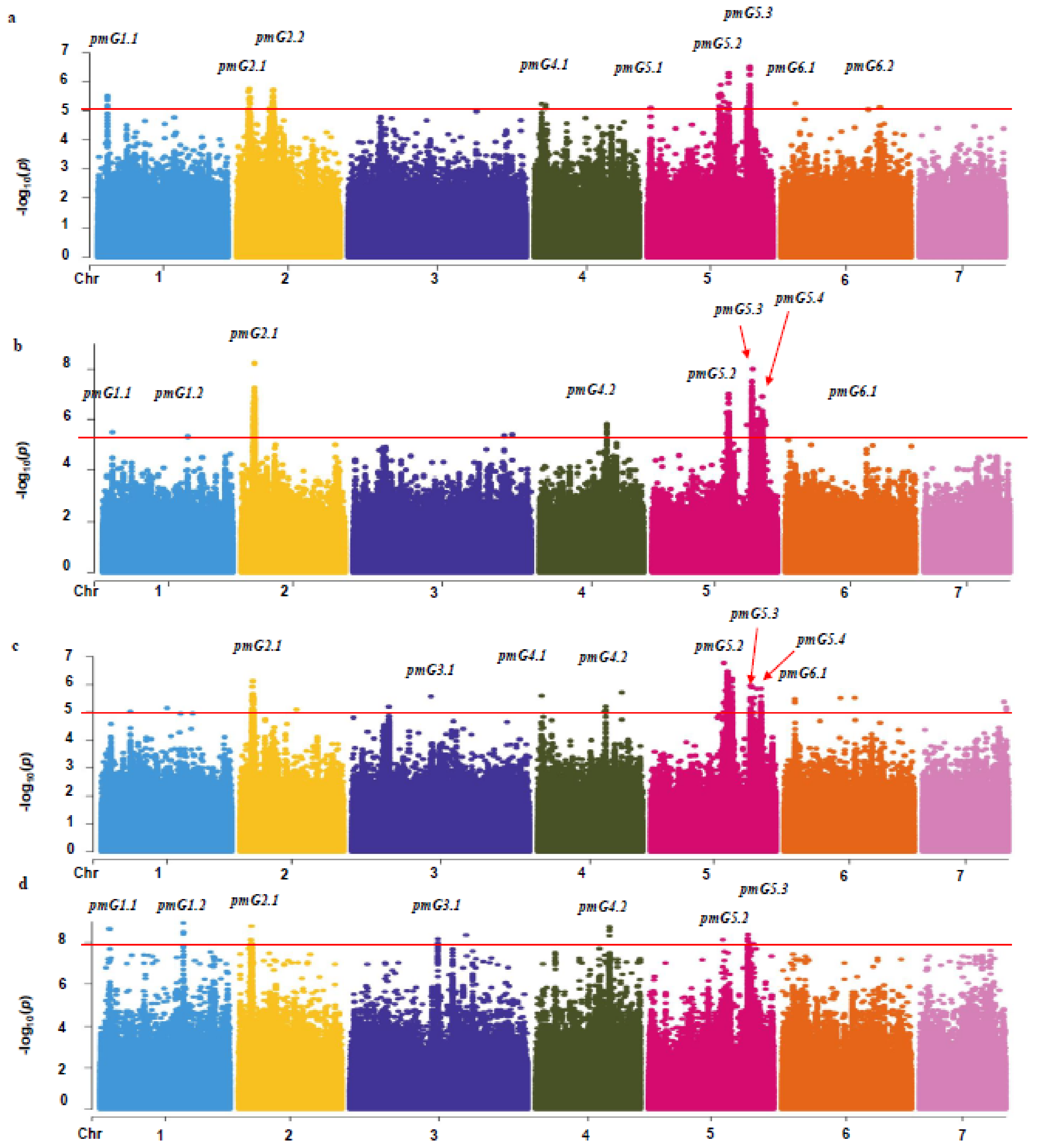
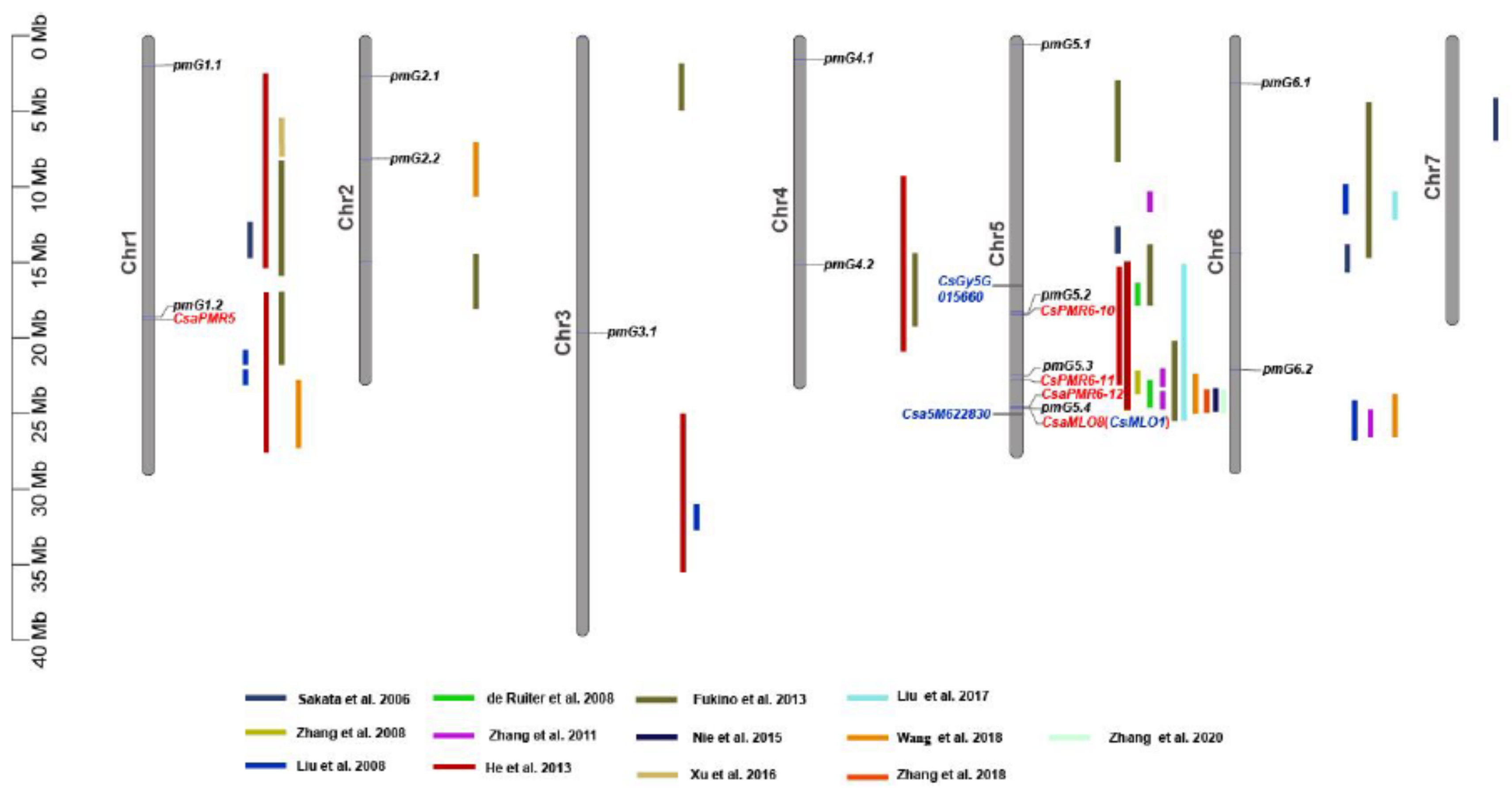
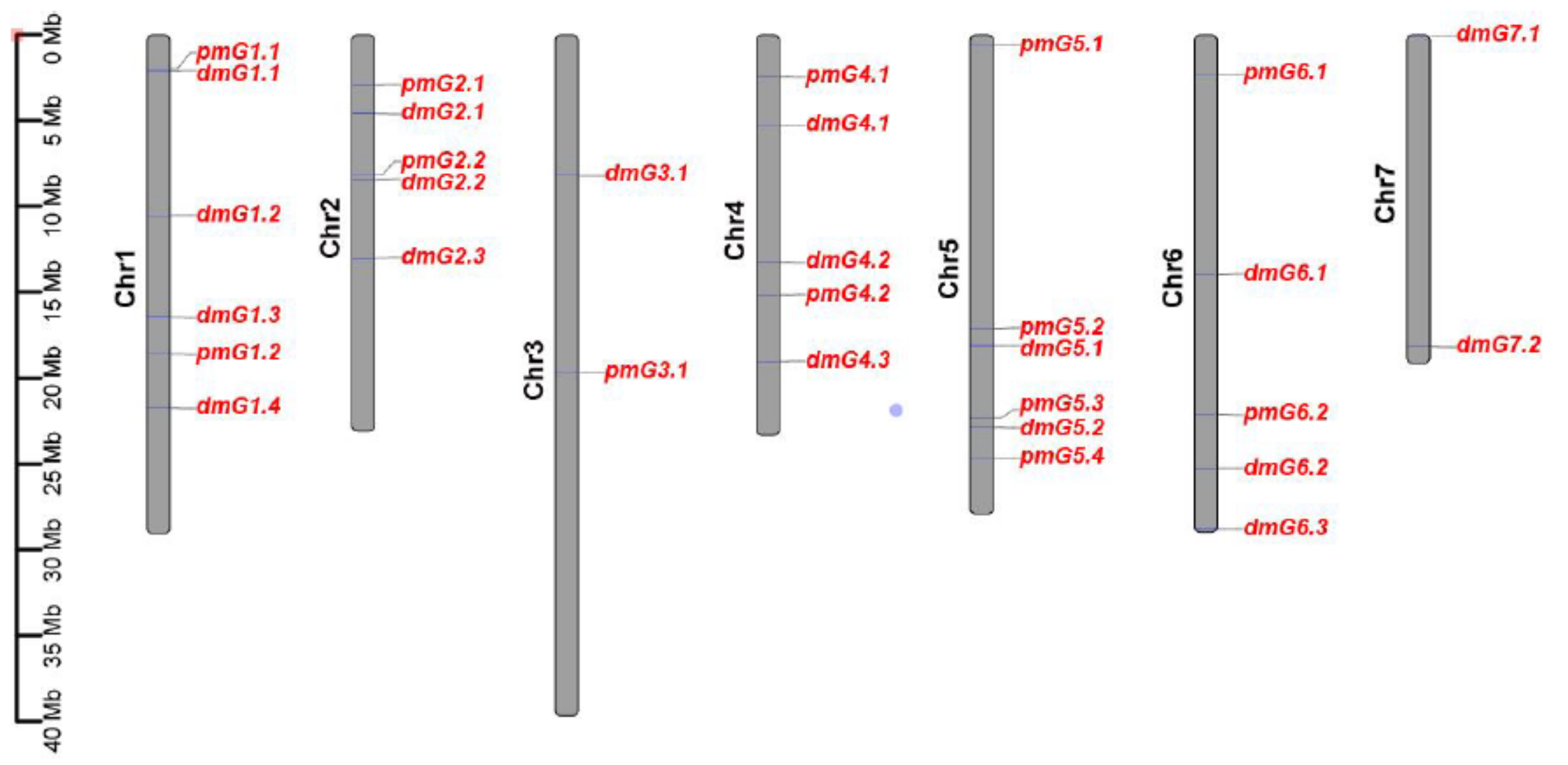
3.2. Genetic Loci Associated with PM Resistance in the CG Population
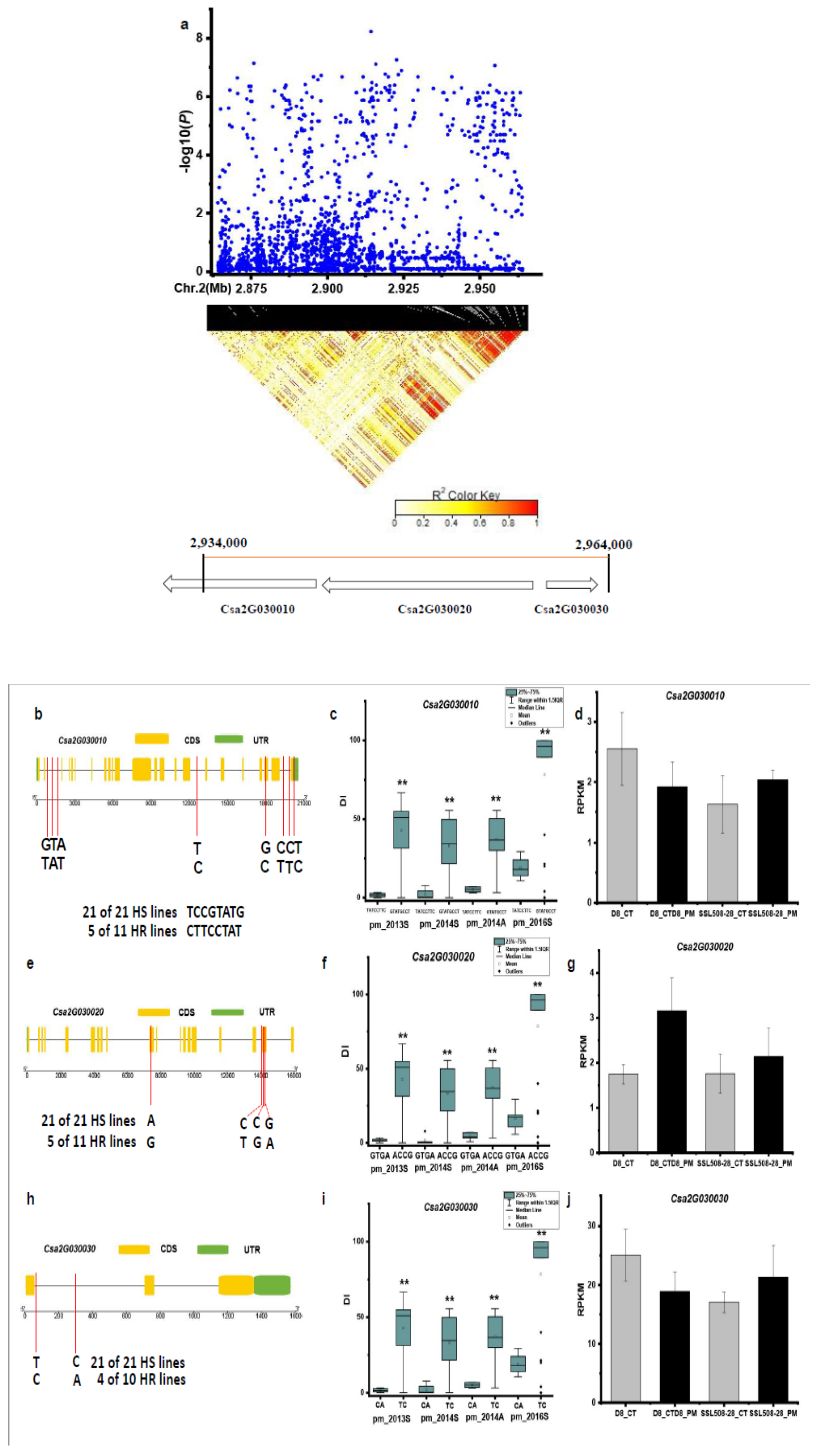
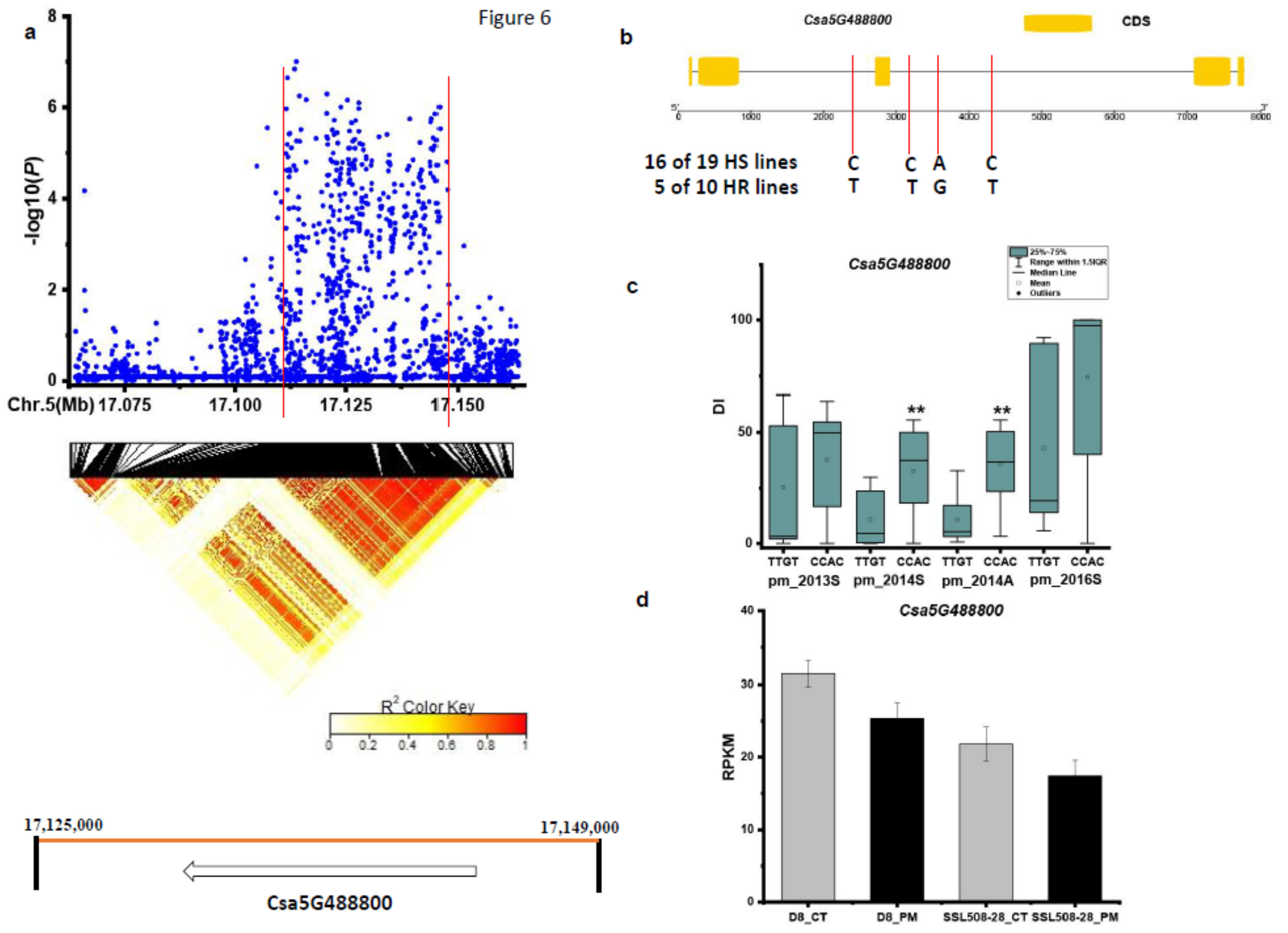
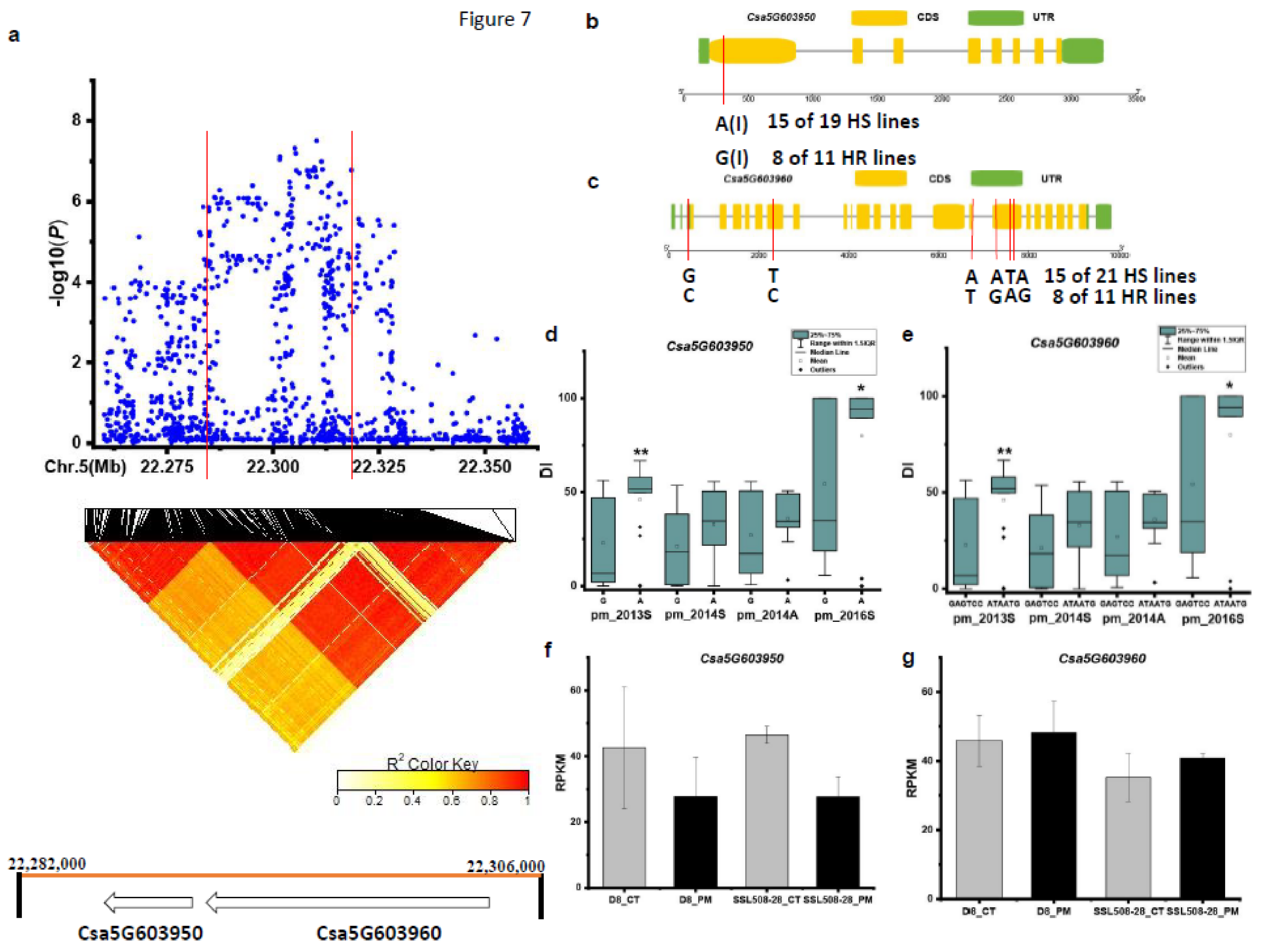
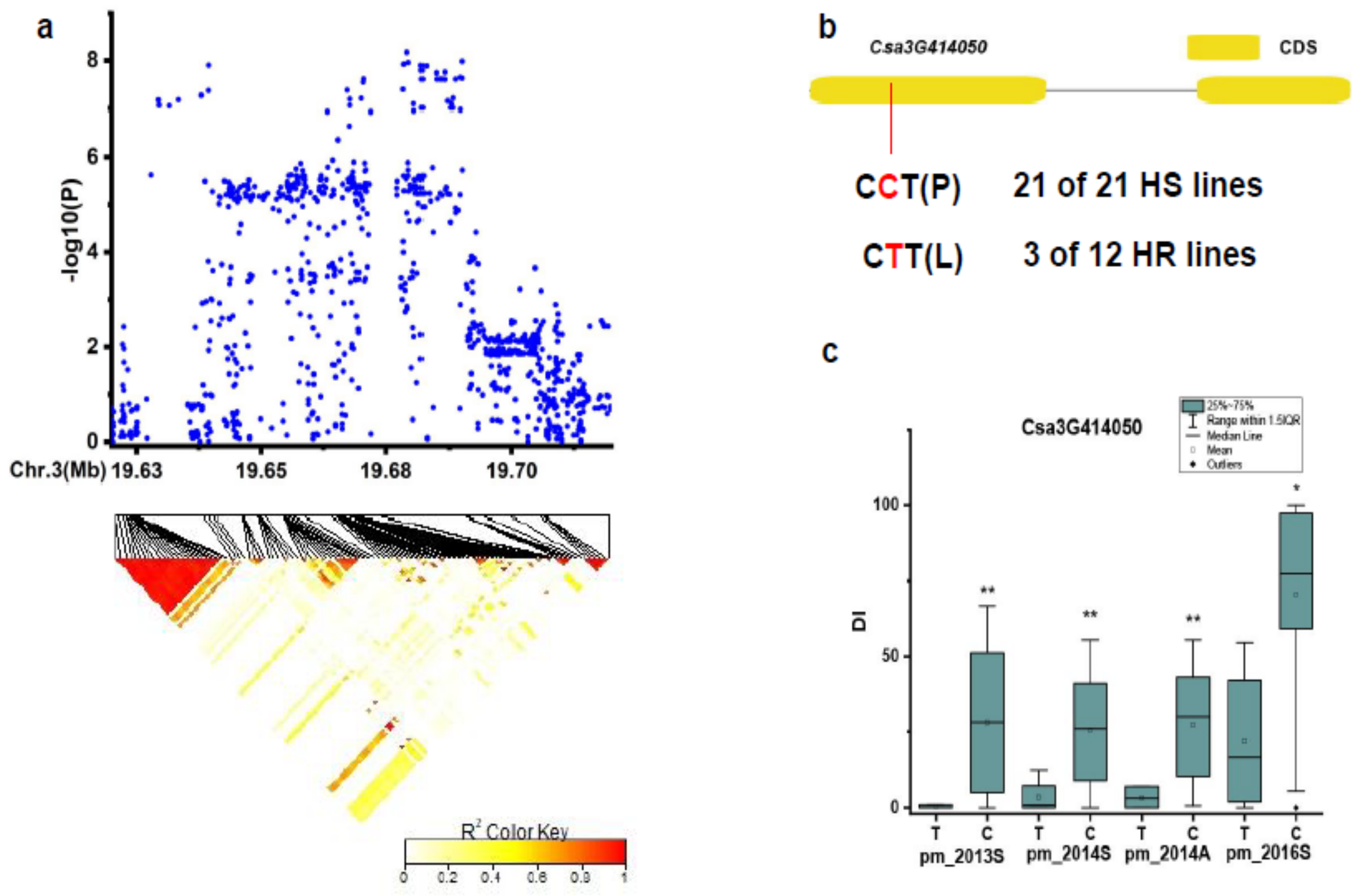
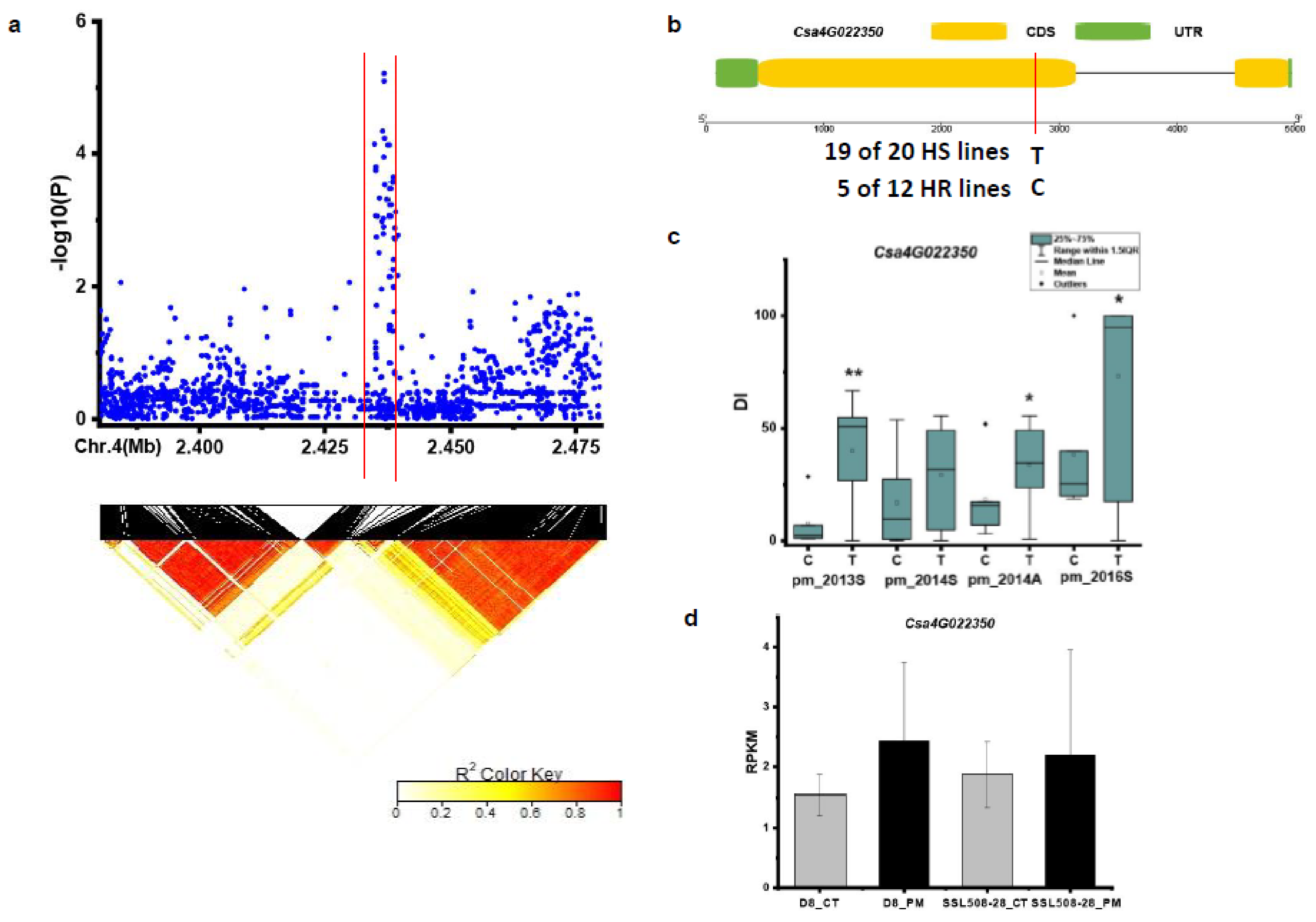
3.3. Candidate Genes for PM Resistance
3.4. Haplotype Analysis of Candidate Genes Reported in Literature for PM Resistance/Susceptibility
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-García, A.; Romero, D.; Fernández-Ortuño, D.; López-Ruiz, F.; De Vicente, A.; Tores, J.A. The powdery mildew fungus Podosphaera fusca (synonym Podosphaera xanthii), a constant threat to cucurbits. Mol. Plant Pathol. 2009, 10, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Ishii, H.; Fraaije, B.A.; Sugiyama, T.; Noguchi, K.; Nishimura, K.; Takeda, T.; Amano, T.; Hollomon, D.W. Occurrence and Molecular Characterization of Strobilurin Resistance in Cucumber Powdery Mildew and Downy Mildew. Phytopathology 2001, 91, 1166–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kooistra, E. Powdery mildew resistance in cucumber. Euphytica 1968, 17, 236–244. [Google Scholar]
- Panstruga, R.; Schulze-Lefert, P. Live and let live: Insights into powdery mildew disease and resistance. Mol. Plant Pathol. 2002, 3, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Zijlstra, S.; Groot, S.P. Search for novel genes for resistance to powdery mildew (Sphaerotheca fuliginae) in cucumber (Cucumis sativus). Euphytica 1992, 64, 31–37. [Google Scholar]
- Block, C.C.; Reitsma, K.R. Powdery Mildew Resistance in the U.S. National Plant Germplasm System Cucumber Collection. HortScience 2005, 40, 416–420. [Google Scholar] [CrossRef] [Green Version]
- Shanmugasundaram, S.; Williams, P.; Peterson, C.J.P. Inheritance of resistance to powdery mildew in cucumber. Phytopathology 1971, 61, 1218–1221. [Google Scholar] [CrossRef]
- Clair, D.A.S. Quantitative Disease Resistance and Quantitative Resistance Loci in Breeding. Annu. Rev. Phytopathol. 2010, 48, 247–268. [Google Scholar] [CrossRef]
- Liu, L.; Yuan, X.; Cai, R.; Pan, J.; Zhu, L. Quantitative Trait Loci for Resistance to Powdery Mildew in Cucumber under Seedling Spray Inoculation and Leaf Disc Infection. J. Phytopathol. 2008, 156, 691–697. [Google Scholar]
- Liu, P.; Miao, H.; Lu, H.; Cui, J.; Tian, G.; Wehner, T.; Gu, X.; Zhang, S. Molecular mapping and candidate gene analysis for resistance to powdery mildew in Cucumis sativus stem. Genet. Mol. Res. 2017, 16, 3. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, M.; Miao, H.; Zhang, S.; Yang, Y.; Xie, B.; Gu, X. QTL mapping of resistance genes to powdery mildew in cucumber (Cucumis sativus L.). Sci. Agric. Sin. 2011, 44, 3584–3593. [Google Scholar]
- Sakata, Y.; Kubo, N.; Morishita, M.; Kitadani, E.; Sugiyama, M.; Hirai, M. QTL analysis of powdery mildew resistance in cucumber (Cucumis sativus L.). Theor. Appl. Genet. 2006, 112, 243–250. [Google Scholar] [CrossRef]
- Wilson, J.; John, C.; Wohler, H.; Hoover, M. Two foreign cucumbers resistant to bacterial wilt and powdery mildew. Plant Dis. Rep. 1956, 40, 437–438. [Google Scholar]
- Smith, P. Powdery Mildew Resistance in Cucumber; APS: Saint Paul, MI, USA, 1948; Volume 38, pp. 1027–1028. [Google Scholar]
- Barnes, W.; Epps, W. Powdery mildew resistance in South Carolina cucumbers. Plant Dis. Rep. 1956, 40, 1093. [Google Scholar]
- Zhang, C.; Anarjan, M.B.; Win, K.T.; Begum, S.; Lee, S. QTL-seq analysis of powdery mildew resistance in a Korean cucumber inbred line. Theor. Appl. Genet. 2021, 134, 435–451. [Google Scholar] [CrossRef]
- He, X.; Li, Y.; Pandey, S.; Yandell, B.S.; Pathak, M.; Weng, Y. QTL mapping of powdery mildew resistance in WI 2757 cucumber (Cucumis sativus L.). Theor. Appl. Genet. 2013, 126, 2149–2161. [Google Scholar] [CrossRef]
- Nie, J.; He, H.; Peng, J.; Yang, X.; Bie, B.; Zhao, J.; Wang, Y.; Si, L.; Pan, J.-S.; Cai, R. Identification and fine mapping of pm5.1: A recessive gene for powdery mildew resistance in cucumber (Cucumis sativus L.). Mol. Breed. 2015, 35, 1–13. [Google Scholar] [CrossRef]
- De Ruiter, W.; Hofstede, R.; de Vries, J.; Van den Heuvel, H.; Pitrat, M. Combining QTLs for resistance to CYSDV and powdery mildew in a single cucumber line. Cucurbitaceae 2008, 181–188. [Google Scholar]
- Zhang, H.; Wang, Z.; Mao, A.; Zhang, F.; Wang, Y.; Xu, Y. SSR markers linked to the resistant gene of cucumber powdery mildew. Acta Agri Boreali Sin. 2008, 23, 77–80. [Google Scholar]
- Wang, Y.; Vandenlangenberg, K.; Wen, C.; Wehner, T.C.; Weng, Y. QTL mapping of downy and powdery mildew resistances in PI 197088 cucumber with genotyping-by-sequencing in RIL population. Theor. Appl. Genet. 2018, 131, 597–611. [Google Scholar] [CrossRef]
- Fukino, N.; Yoshioka, Y.; Sugiyama, M.; Sakata, Y.; Matsumoto, S. Identification and validation of powdery mildew (Podosphaera xanthii)-resistant loci in recombinant inbred lines of cucumber (Cucumis sativus L.). Mol. Breed. 2013, 32, 267–277. [Google Scholar] [CrossRef]
- Nie, J.; Wang, Y.; He, H.; Guo, C.; Zhu, W.; Pan, J.; Li, D.; Lian, H.; Pan, J.; Cai, R. Loss-of-function mutations in CsMLO1 confer durable powdery mildew resistance in cucumber (Cucumis sativus L.). Front. Plant Sci. 2015, 6, 1155. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yu, T.; Xu, R.; Shi, Y.; Lin, X.; Xu, Q.; Qi, X.; Weng, Y.; Chen, X. Fine mapping of a dominantly inherited powdery mildew resistance major-effect QTL, Pm1.1, in cucumber identifies a 41.1 kb region containing two tandemly arrayed cysteine-rich receptor-like protein kinase genes. Theor. Appl. Genet. 2016, 129, 507–516. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, X.; Zhu, W.; Qin, X.; Xu, J.; Cheng, C.; Lou, Q.; Li, J.; Chen, J. Complete resistance to powdery mildew and partial resistance to downy mildew in a Cucumis hystrix introgression line of cucumber were controlled by a co-localized locus. Theor. Appl. Genet. 2018, 131, 2229–2243. [Google Scholar] [CrossRef] [PubMed]
- Pavan, S.; Jacobsen, E.; Visser, R.G.F.; Bai, Y. Loss of susceptibility as a novel breeding strategy for durable and broad-spectrum resistance. Mol. Breed. 2009, 25, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Schouten, H.J.; Krauskopf, J.; Visser, R.G.F.; Bai, Y. Identification of candidate genes required for susceptibility to powdery or downy mildew in cucumber. Euphytica 2014, 200, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Berg, J.A.; Appiano, M.; Martínez, M.S.; Hermans, F.W.K.; Vriezen, W.H.; Visser, R.G.F.; Bai, Y.; Schouten, H.J. A transposable element insertion in the susceptibility gene CsaMLO8 results in hypocotyl resistance to powdery mildew in cucumber. BMC Plant Biol. 2015, 15, 243. [Google Scholar] [CrossRef] [Green Version]
- Van Schie, C.C.; Takken, F.L. Susceptibility genes 101: How to be a good host. Annu. Rev. Phytopathol. 2014, 52, 551–581. [Google Scholar] [CrossRef]
- Qi, J.; Liu, X.; Shen, D.; Miao, H.; Xie, B.; Li, X.; Zeng, P.; Wang, S.; Shang, Y.; Gu, X.; et al. A genomic variation map provides insights into the genetic basis of cucumber domestication and diversity. Nat. Genet. 2013, 45, 1510–1515. [Google Scholar] [CrossRef]
- Zhang, S.P.; Liu, M.M.; Miao, H.; Yang, Y.H.; Xie, B.Y.; Wehner, T.C.; Gu, X.F. Chromosomal Mapping and QTL Analysis of Resistance to Downy Mildew in Cucumis sativus. Plant Dis. 2013, 97, 245–251. [Google Scholar] [CrossRef] [Green Version]
- SAS Institute. SAS/OR 9.3 User’s Guide: Mathematical Programming Examples; SAS Institute Inc.: Cary, NC, USA, 2012. [Google Scholar]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Lu, H.; Liu, P.; Miao, H.; Bai, Y.; Gu, X.; Zhang, S. Identification of Novel Loci and Candidate Genes for Cucumber Downy Mildew Resistance Using GWAS. Plants 2020, 9, 1659. [Google Scholar] [CrossRef]
- Wang, Y.; Bo, K.; Gu, X.; Pan, J.; Li, Y.; Chen, J.; Wen, C.; Ren, Z.; Ren, H.; Chen, X.; et al. Molecularly tagged genes and quantitative trait loci in cucumber with recommendations for QTL nomenclature. Hortic. Res. 2020, 7, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Pang, Z.; Zhao, S.; Zhang, L.; Lv, Q.; Yin, D.; Li, D.; Liu, X.; Zhao, X.; Li, X.; et al. Importance of OsRac1 and RAI1 in signalling of nucleotide-binding site leucine-rich repeat protein-mediated resistance to rice blast disease. New Phytol. 2019, 223, 828–838. [Google Scholar] [CrossRef]
- Wan, H.; Yuan, W.; Bo, K.; Shen, J.; Pang, X.; Chen, J. Genome-wide analysis of NBS-encoding disease resistance genes in Cucumis sativus and phylogenetic study of NBS-encoding genes in Cucurbitaceae crops. BMC Genom. 2013, 14, 109. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef] [Green Version]
- Berg, J.A.; Hermans, F.W.K.; Beenders, F.; Lou, L.; Vriezen, W.H.; Visser, R.G.F.; Bai, Y.; Schouten, H.J. Analysis of QTL DM4.1 for Downy Mildew Resistance in Cucumber Reveals Multiple subQTL: A Novel RLK as Candidate Gene for the Most Important subQTL. Front. Plant Sci. 2020, 11, 569876. [Google Scholar] [CrossRef]











| Genetic Locus | Gene Number | Predicted Protein |
|---|---|---|
| pmG2.1 | Csa2G030010- | DNA polymerase |
| Csa2G030020 | a Lin-9-like protein | |
| Csa2G030030 | an unknown protein | |
| pmG3.1 | Csa3G414050 | Unknown protein |
| pmG4.1 | homolog of Csa4G022350 | Leucine-rich repeat protein kinase family protein |
| pmG5.2 | Csa5G488800 | RNA recognition motif |
| pmG5.3 | Csa5G603950 | Pyruvate dehydrogenase E1 component α subunit |
| Csa5G603960 | Paired amphipathic helix protein Sin3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Gu, X.; Lu, H.; Liu, P.; Miao, H.; Bai, Y.; Zhang, S. Identification of Novel Loci and Candidate Genes for Resistance to Powdery Mildew in a Resequenced Cucumber Germplasm. Genes 2021, 12, 584. https://doi.org/10.3390/genes12040584
Liu X, Gu X, Lu H, Liu P, Miao H, Bai Y, Zhang S. Identification of Novel Loci and Candidate Genes for Resistance to Powdery Mildew in a Resequenced Cucumber Germplasm. Genes. 2021; 12(4):584. https://doi.org/10.3390/genes12040584
Chicago/Turabian StyleLiu, Xiaoping, Xingfang Gu, Hongwei Lu, Panna Liu, Han Miao, Yuling Bai, and Shengping Zhang. 2021. "Identification of Novel Loci and Candidate Genes for Resistance to Powdery Mildew in a Resequenced Cucumber Germplasm" Genes 12, no. 4: 584. https://doi.org/10.3390/genes12040584
APA StyleLiu, X., Gu, X., Lu, H., Liu, P., Miao, H., Bai, Y., & Zhang, S. (2021). Identification of Novel Loci and Candidate Genes for Resistance to Powdery Mildew in a Resequenced Cucumber Germplasm. Genes, 12(4), 584. https://doi.org/10.3390/genes12040584