
MTHFR C677T and A1298C Polymorphisms in Breast Cancer, Gliomas and Gastric Cancer: A Review


Abstract
:1. Introduction
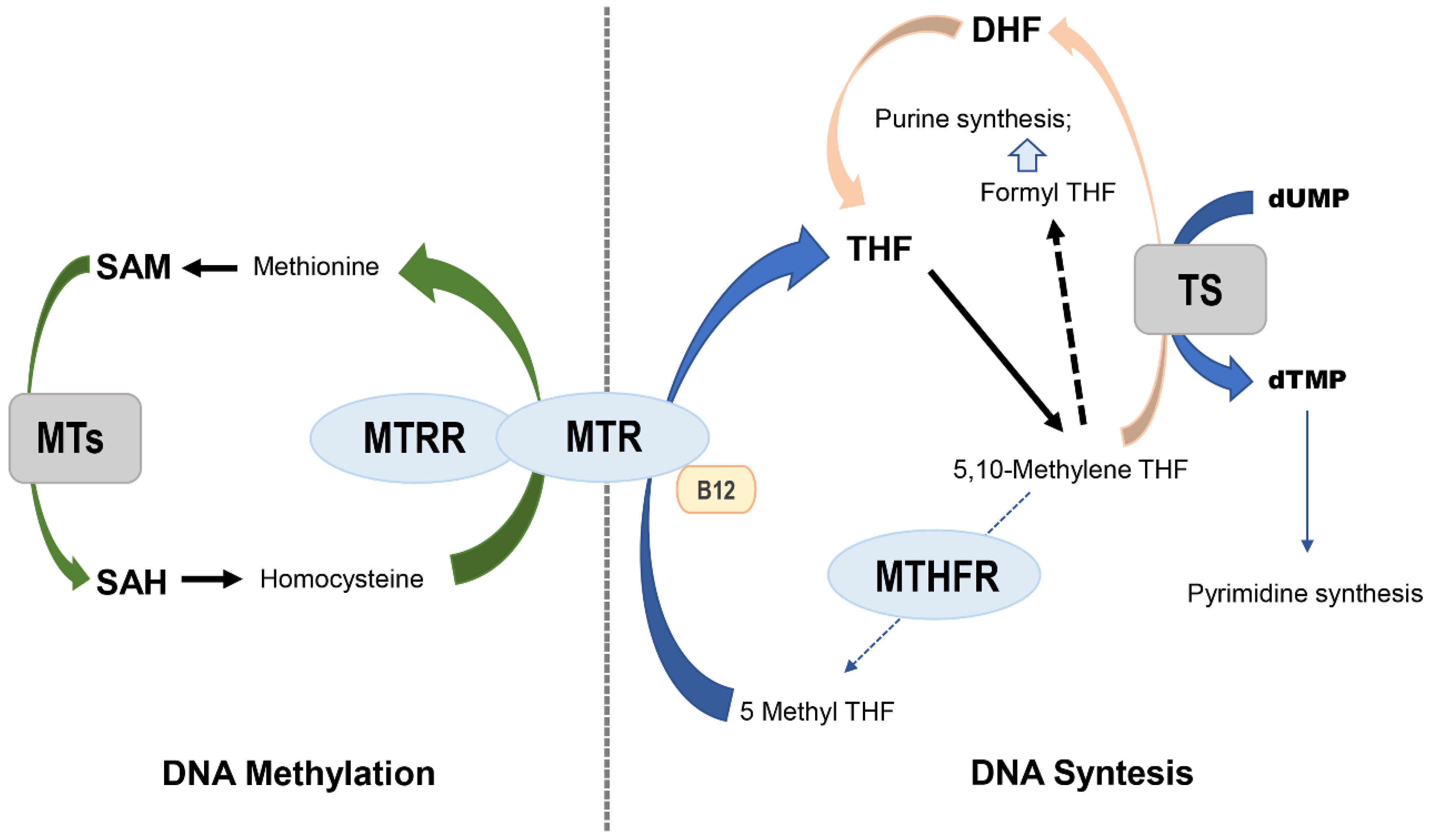
2. MTHFR
3. MTHFR C677T and A1298C Polymorphisms and Cancer
4. MTHFR C677T and A1298C Polymorphisms and Breast Cancer

{kind=link}
| Study Type | Number of Participants | Ethnicity | SNP | Risk Association | References |
|---|---|---|---|---|---|
| Case-control | 560 BC cases and 560 controls | Chinese | C677T | Significant association of the TT genotype with the risk of BC. | [66] |
| Case-control | 150 BC cases and 150 controls | Jordanian | C677T A1298C | Influence of the T allele and TT genotype on the risk of BC. No significant differences for A1298C in BC risk. | [67] |
| Case-control | 100 BC cases and 142 controls | Iranian | C677T | Correlation of the T allele and TT genotype with the risk of BC and menopause status. | [68] |
| Case-control | 669 BC cases and 682 controls | Chinese | C677T A1298C | TT genotype associated with BC risk. Low folate intake associated with the susceptibility to BC at 1289A/A, 677C/C and 677C/T. | [69] |
| Case-control | 1459 BC cases and 1556 controls | Chinese | C677T | Highest prevalence of BC was observed in women with the T allele and low folate intake. | [71] |
| Meta-analysis | 19,260 BC cases and 23,364 controls | Asian and Caucasian | C677T | Strong trend toward the risk for BC in the TT and CT genotypes. Increased risk in Asian and Caucasian populations. | [72] |
| Case-control and Meta-analysis pooled | 28,619 BC cases and 32,388 controls | Indian | C677T | No association between the exchange of T for C at position 677 of MTHFR. | [73] |
| Meta-analysis | 25,877 BC cases and 29,781 controls | Asian, Caucasian and Mixed population | C677T A1298C | Positive association between C677T and the risk of BC, particularly in Asian populations. Non-significant association of A1298C with BC. The A allele affects BC risk in Caucasian populations and a reverse effect in Asian and mixed populations. | [74] |
| Meta-analysis | 3362 BC cases and 4175 controls | Latin American | C677T A1298C | High risk of BC for C677T. No evidence of a relationship between the A1298C polymorphism and BC. | [75] |
| Case-control | 100 BC cases and 144 controls | Brazilian | C677T | Significant association of TT with an increased risk of BC. Relationship between alcohol consumption and the incidence of BC. | [76] |
| Case-control | 315 BC cases and 604 controls | Kazakhstan | C677T A1298C | CT and TT genotypes associated with an increased risk of BC. AC and CC genotypes associated with a decreased risk of BC. | [79] |
| Case-control | 96 BC cases and 117 controls | Moroccan | C677T | T allele and TT genotype associated with increased risk for BC. Correlation between polymorphism and PR expression. | [80] |
| Case-control | 253 BC cases and 257 controls | Brazilian | C677T A1298C | C677T associated with metastasis and ER expression. AA genotype associated with higher degrees of BC and ER expression. | [81] |
| Meta-analysis | 19,527 BC cases and 23,123 controls | Asian, Caucasian and Mixed population | A1298C | Association between the C allele and CC genotype and increased risk for BC. | [82] |
| Case-control | 61 BC cases and 63 controls | Brazilian | C677T A1298C TYMS | No association between C677T and A1298C polymorphisms and BC. Association between TYMS polymorphism with risk of more aggressive BC subtypes | [83] |
| Case-control | 58 BC cases and 58 controls | Italian | C677T A1298C | Polymorphisms related to BC progression. | [84] |
| Case-control | 610 BC cases and 1207 controls | North American | C677T A1298C | No association between polymorphisms and BC. Relationship between folate levels and BC. | [85] |
| Case-control | 100 BC cases and 60 controls | Egyptian | C677T A1298C | Association between TC mutant haplotype and BC aggressiveness. | [86] |
| Case-control | 124 BC cases and 63 controls | Asian | C677T A1298C | TT genotype associated with BC risk in advanced age patients | [87] |
5. MTHFR C677T and A1298C Polymorphisms and Glioma
| Study Type | Number of Participants | Ethnicity | SNP | Histological Stratification | Risk Association | Reference |
|---|---|---|---|---|---|---|
| Case-control Pediatric tumors | 284 cases and 464 controls | Caucasian, East Slav and Russian (grouped together) | C677T | Glial tumors and embryonic brain tumors | No risk associated with brain tumor development | [100] |
| Case-control Pediatric tumors | 73 cases and 205 controls | Thai | C677T | Brain tumors No stratification | Statistically non-significant 5.2 times increased risk of glial tumors for the homozygous TT allele | [99] |
| A1298C | Increased risk of embryonic tumors No risk associated with glial and germ cell tumors | |||||
| Case-control Hospital based | 108 gliomas, 76 meningiomas and 104 controls | Indian | C677T | No WHO malignancy grade stratification | No risk associated with gliomas or meningioma development | [107] |
| A1298C | Increased risk of glioma 38% reduced risk of meningioma for CC and ‘C’ allele containing genotypes | |||||
| Case-control | 112 glioma cases and 141 controls | Indian | C677T | Astrocytoma, glioblastoma, oligodendroglioma and other types of glioma | No risk associated with any glioma type or overall survival | [112] |
| Case-control Hospital based | 39 HGGs, 35 meningiomas and 98 controls | Turkish | C677T | High-grade gliomas (HGG) and meningiomas | No risk associated with meningioma Non-significant 2.15 times increased risk of HGG for the homozygous TT allele | [115] |
| Case-control | 93 cases and 93 controls | mixed Brazilian | C677T | Astrocytic tumors subdivided in WHO grade I (17), grade II (19), grade III (14), and grade IV (43) | Potential protective effect for the homozygous TT genotype No risk associated with histological subtypes | [114] |
| Case-control | 6oo cases and 600 controls | Chinese Han population | C677T | Meningiomas subdivided in WHO grade I (391), grade II (167) and grade III (42) | Reduced risk associated with TT and ‘T’ allele-containing genotypes No risk associated with subtypes | [119] |
| A1298C | No association with meningioma general risk or subtypes | |||||
| Case-control | 317 cases and 320 controls | Northern Chinese Han population | C677T | Meningioma without WHO grade stratification | Increased risk of meningioma for the TT genotype | [120] |
| A1298C | No risk association | |||||
| Case-control | 631 meningioma, 1005 glioma and 1098 controls | Caucasian | C677T | GBM, oligodendrocytes, other astrocytomas and other gliomas’ subtypes | Increased risk of meningioma, but not glioma | [10] |
| A1298C | Increased risk of meningioma, glioblastoma and oligodendroglioma for the heterozygous genotype | |||||
| Meta-analysis | 1323 cases and 1883 controls from 10 studies | Caucasian, Chinese, Asian | C677T | Meningioma | No risk association | [122] |
| 1855 cases and 3331 controls | A1298C | Increased risk for Caucasian populations in heterozygous (AC) and dominant (CC + AC) models | ||||
| Meta-analysis | 1615 cases and 1909 controls | Asian and Caucasian | C677T | Meningioma without WHO grade stratification | Increased meningioma risk for CT genotype carriers in the total population. No risk associated with Asian populations and increased risk for Caucasian populations with CT and TT genotypes | [121] |
| Meta-analysis | 1786 cases and 2076 controls | Asian, Brazilian and Caucasian | C677T | Glioma without WHO grade stratification | No association was observed for total population or Caucasian populations | [116] |
| Meta-analysis | 3059 cases and 3324 controls | Asian, Brazilian and Caucasian | C677T | Glioma and meningioma | Increased risk for T allele carriers (TC + TT) and 1.38 times increased risk of meningioma for TC carriers Asian populations had an increased brain tumor risk, but no association was observed for Caucasian populations | [117] |
| Meta-analysis | 2236 cases and 2248 controls from five studies | Asian and Caucasian | A1298C | Glioma and meningioma | Increased glioma risk in the total population. In Caucasian populations, increased risk of meningioma and glioma in the heterozygous model (AC) and dominant model (CC + AC) | [118] |
| Case-control | 328 cases and 400 controls | Caucasian | C677T | Glioblastoma | No risk associated with the polymorphism | [111] |
| Retrospective cohort study | 214 patients | Caucasian | C677T | Glioblastoma | Poor overall survival in patients younger than 60 years | [123] |
| A1298C | No association with overall survival |
6. MTHFR C677T and A1298C Polymorphisms and Gastric Cancer
| Study Type | Number of Participants | Ethnicity | SNP | Risk Association | References |
|---|---|---|---|---|---|
| Case-control | 70 GC and 61 controls | Turkish | C677T A1298C | The AC genotype of the A1298C polymorphism is a risk factor for GC. The TT genotype of C677T was not at a higher risk than the CC genotype. | [135] |
| Case-control | 307 GC and 560 controls | Chinese | C677T | TT genotype associated with a decreased risk for GC | [136] |
| Case-control | 107 GC and 220 controls | Chinese | C677T | MTHFR variant genotypes + smoking and drink habits are associated with a higher risk for GC | [137] |
| Meta-analysis | 5757 GC and 8501 controls | Asian and Caucasian | C677T | Significant association was found between GC and the MTHFR C677T polymorphism. Elevated risk of GC in Asian individuals carrying the MTHFR C677T polymorphism, but not in Caucasian populations | [138] |
| Meta-analysis | 1584 GC and 2785 controls | Asian and Caucasian | C677T A1298C | MTHFR C677T and A1298C polymorphisms, respectively, contribute to the susceptibility of GC. In East Asian populations with C677T, the association was significant but not in Caucasian populations. The A1298C polymorphism was associated with GCA in East Asian populations. | [139] |
| Case-control | 790 GC and 202 controls | Italian | C677T A1298C | Increased risk of GC for the C677T variant (homozygous TT), but no effect of the A1298C polymorphism. | [140] |
| Meta-analysis | 2727 | Asian, Caucasian and Mixed | C677T | MTHFR C677T was associated with GC | [141] |
| Meta-analysis | 6572 GC and 9584 controls | Asian, Caucasian and Mixed | C677T A1298C | C677T was related to a significantly increased risk for GC. No correlation was found with A1298C. C677T, but not A1298C, was associated with an increased risk of GC in Asian and Caucasian populations | [142] |
| Meta-analysis | 4070/6462 cases/controls for C677T and 1923/3561 cases/controls for A1298C polymorphism | Eastern and Western | C677T A1298C | No significant association was found in the CC genotype of A1298C. The C677T allele T was associated with an increased risk of GC. Subgroup analyses revealed an increased risk for Asian populations but not for Caucasian populations | [143] |
| Case-control | 76 GC and 91 controls | Turkish | C677T | CT heterozygotes had a lower susceptibility to GC | [144] |
| Meta-analysis | 1718 | Asian, European and Mixed | C677T | TT was related more to hematologic toxicity than the CC or CT genotype | [147] |
| Multicenter, single-arm, phase 2 study | 60 | Dutch | C677T | The TT genotype was related to inferior progression-free survival and OS | [150] |
| Retrospective comparative exploratory study | 218 | German | C677T A1298C | A1298C was an independent prognostic factor associated with a poor prognosis in neoadjuvantly treated GC patients | [151] |
| Single-center, cross sectional observational trial | 128 | Chinese | C677T | The TT genotype showed an increased risk of moderate-to-severe precancerous gastric lesions | [152] |
| Case-control study | 450 GC and 780 controls | Iranian | C677T | MTHFR C677T carriers who were also positive for H. pylori, showed an increased risk for GC | [153] |
| Case-control | 58 GC patients with H. pylori infection and 94 non-infected patients | Chinese | C677T | TT genotype was considered a susceptibility factor of H. pylori infection. | [154] |
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Berry, R.J.; Bailey, L.; Mulinare, J.; Bower, C.; Dary, O. Fortification of Flour with Folic Acid. Food Nutr. Bull. 2010, 31, S22–S35. [Google Scholar] [CrossRef]
- Ebara, S. Nutritional role of folate. Congenit. Anom. 2017, 57, 138–141. [Google Scholar] [CrossRef]
- Salazar, M.D.; Ratnam, M. The folate receptor: What does it promise in tissue-targeted therapeutics? Cancer Metastasis Rev. 2007, 26, 141–152. [Google Scholar] [CrossRef]
- Stover, P.J. One-Carbon Metabolism–Genome Interactions in Folate-Associated Pathologies. J. Nutr. 2009, 139, 2402–2405. [Google Scholar] [CrossRef] [Green Version]
- Bae, S.; Ulrich, C.M.; Bailey, L.B.; Malysheva, O.; Brown, E.C.; Maneval, D.R.; Neuhouser, M.L.; Cheng, T.Y.D.; Miller, J.W.; Caudill, M.A.; et al. Impact of folic acid fortification on global DNA methylation and one-carbon biomarkers in the Women’s Health Initiative Observational Study cohort. Epigenetics 2014, 9, 396–403. [Google Scholar] [CrossRef] [Green Version]
- Bailey, L.B.; Stover, P.J.; McNulty, H.; Fenech, M.F.; Gregory, J.F.; Mills, J.L.; Pfeiffer, C.M.; Fazili, Z.; Zhang, M.; Ueland, P.M.; et al. Biomarkers of Nutrition for Development—Folate Review. J. Nutr. 2015, 145, 1636S–1680S. [Google Scholar] [CrossRef] [Green Version]
- Duthie, S.J.; Narayanan, S.; Brand, G.M.; Pirie, L.; Grant, G. Impact of Folate Deficiency on DNA Stability. J. Nutr. 2002, 132, 2444S–2449S. [Google Scholar] [CrossRef]
- Donnelly, J.G. Folic Acid. Crit. Rev. Clin. Lab. Sci. 2001, 38, 183–223. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.H.; Pittman, D.L.; Wyatt, M.D. Uracil in DNA: Consequences for carcinogenesis and chemotherapy. Biochem. Pharm. 2008, 76, 697–706. [Google Scholar] [CrossRef] [Green Version]
- Bethke, L.; Webb, E.; Murray, A.; Schoemaker, M.; Feychting, M.; Lönn, S.; Ahlbom, A.; Malmer, B.; Henriksson, R.; Auvinen, A.; et al. Functional Polymorphisms in Folate Metabolism Genes Influence the Risk of Meningioma and Glioma. Cancer Epidemiol. Biomark. Prev. 2008, 17, 1195–1202. [Google Scholar] [CrossRef] [Green Version]
- Hasan, T.; Arora, R.; Bansal, A.K.; Bhattacharya, R.; Sharma, G.S.; Singh, L.R. Disturbed homocysteine metabolism is associated with cancer. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef]
- Bravatà, V. Controversial roles of methylenetetrahydrofolate reductase polymorphisms and folate in breast cancer disease. Int. J. Food Sci. Nutr. 2014, 66, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Ito, H.; Wakai, K.; Hirose, K.; Saito, T.; Suzuki, T.; Kato, T.; Hirai, T.; Kanemitsu, Y.; Hamajima, H.; et al. One-carbon metabolism related gene polymorphisms interact with alcohol drinking to influence the risk of colorectal cancer in Japan. Carcinogenesis 2005, 26, 2164–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singal, R.; Ferdinand, L.; Das, P.M.; Reis, I.M.; Schlesselman, J.J. Polymorphisms in the methylenetetrahydrofolate reductase gene and prostate cancer risk. Int. J. Oncol. 2004, 25, 1465–1471. [Google Scholar] [CrossRef]
- Robien, K.; Ulrich, C.M. 5,10-Methylenetetrahydrofolate reductase polymorphisms and leukemia risk: A HuGE minireview. Am. J. Epidemiol. 2003, 157, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Hickey, S.E.; Curry, C.J.; Toriello, H.V. ACMG Practice Guideline: Lack of evidence for MTHFR polymorphism testing. Genet. Med. 2013, 15, 153–156. [Google Scholar] [CrossRef] [Green Version]
- Tsang, B.L.; Devine, O.J.; Cordero, A.M.; Marchetta, C.M.; Mulinare, J.; Mersereau, P.; Guo, J.; Qi, Y.P.; Berry, R.J.; Rosenthal, J.; et al. Assessing the association between the methylenetetrahydrofolate reductase (MTHFR) 677C>T polymorphism and blood folate concentrations: A systematic review and meta-analysis of trials and observational studies. Am. J. Clin. Nutr. 2015, 101, 1286–1294. [Google Scholar] [CrossRef]
- De Mattia, E.; Toffoli, G. C677T and A1298C MTHFR polymorphisms, a challenge for antifolate and fluoropyrimidine-based therapy personalisation. Eur. J. Cancer 2009, 45, 1333–1351. [Google Scholar] [CrossRef] [PubMed]
- Farkas, S.A.; Böttiger, A.K.; Isaksson, H.S.; Finnell, R.H.; Ren, A.; Nilsson, T.K.; Nilsson, T.K. Epigenetic alterations in folate transport genes in placental tissue from fetuses with neural tube defects and in leu-kocytes from subjects with hyperhomocysteinemia. Epigenetics 2013, 8, 303–316. [Google Scholar] [CrossRef] [Green Version]
- Toffoli, G.; De Mattia, E. Pharmacogenetic relevance of MTHFR polymorphisms. Pharmacogenomics 2008, 9, 1195–1206. [Google Scholar] [CrossRef]
- Esteller, M. Epigenetics in cancer. N. Engl. J. Med. 2008, 358, 1148–1159. [Google Scholar] [CrossRef]
- Semmler, A.; Simon, M.; Moskau, S.; Linnebank, M. Polymorphisms of methionine metabolism and susceptibility to meningioma formation: Laboratory investigation. J. Neurosurg. 2008, 108, 999–1004. [Google Scholar] [CrossRef] [Green Version]
- Ulrich, C.M. Nutrigenetics in Cancer Research—Folate Metabolism and Colorectal Cancer. J. Nutr. 2005, 135, 2698–2702. [Google Scholar] [CrossRef]
- Goyette, P.; Pai, A.; Milos, R.; Frosst, P.; Tran, P.; Chen, Z.; Chan, M.; Rozen, R. Gene structure of human and mouse methylenetetrahydrofolate reductase (MTHFR). Mamm. Genome 1998, 9, 652–656. [Google Scholar] [CrossRef]
- Goyette, P.; Rozen, R. The thermolabile variant 677C?T can further reduce activity when expressed in CIS with severe mutations for human methylenetetrahydrofolate reductase. Hum. Mutat. 2000, 16, 132–138. [Google Scholar] [CrossRef]
- Levin, B.L.; Varga, E. MTHFR: Addressing Genetic Counseling Dilemmas Using Evidence-Based Literature. J. Genet. Couns. 2016, 25, 901–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.-H.; Botto, L.D.; Gallagher, M.; Friedman, J.M.; Sanders, C.L.; Koontz, D.; Nikolova, S.; Erickson, J.D.; Steinberg, K. Prevalence and effects of gene-gene and gene-nutrient interactions on serum folate and serum total homocysteine concentrations in the United States: Findings from the third National Health and Nutrition Examination Survey DNA Bank. Am. J. Clin. Nutr. 2008, 88, 232–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, N.; Murata, M.; Ikeda, Y.; Opare-Sem, O.; Zivelin, A.; Geffen, E.; Seligsohn, U. The frequent 5,10-methylenetetrahydrofolate reductase C677T polymorphism is associated with a common haplo-type in whites, Japanese, and Africans. Am. J. Hum. Genet 2002, 70, 758–762. [Google Scholar] [CrossRef] [Green Version]
- Kauwell, G.P.; Wilsky, C.E.; Cerda, J.J.; Herrlinger-Garcia, K.; Hutson, A.D.; Theriaque, D.W.; Boddie, A.; Rampersaud, G.C.; Bailey, L.B. Methylenetetrahydrofolate reductase mutation (677C → T) negatively influences plasma homocysteine response to marginal folate intake in elderly women. Metabolism 2000, 49, 1440–1443. [Google Scholar] [CrossRef]
- Rozen, R. Genetic Predisposition to Hyperhomocysteinemia: Deficiency of Methylenetetrahydrofolate Reductase (MTHFR). Thromb. Haemost. 1997, 78, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Frosst, P.; Blom, H.; Milos, R.; Goyette, P.; Sheppard, C.; Matthews, R.; Boers, G.; Heijer, M.D.; Kluijtmans, L.; Heuve, L.V.D.; et al. A candidate genetic risk factor for vascular disease: A common mutation in methylenetetrahydrofolate reductase. Nat. Genet. 1995, 10, 111–113. [Google Scholar] [CrossRef]
- Ulrich, C.M.; Robien, K.; Sparks, R. Pharmacogenetics and folate metabolism –a promising direction. Pharmacogenomics 2002, 3, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Lievers, K.J.; Boers, G.H.; Verhoef, P.; Heijer, M.; Kluijtmans, L.A.; Put, N.M.; Trijbels, F.J.; Blom, H.J. A second common variant in the methylenetetrahydrofolate reductase (MTHFR) gene and its relationship to MTHFR enzyme activity, homocysteine, and cardiovascular disease risk. J. Mol. Med. 2001, 79, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Friedman, G.; Goldschmidt, N.; Friedlander, Y.; Ben-Yehuda, A.; Selhub, J.; Babaey, S.; Mendel, M.; Kidron, M.; Bar-On, H. A Common Mutation A1298C in Human Methylenetetrahydrofolate Reductase Gene: Association with Plasma Total Homocysteine and Folate Concentrations. J. Nutr. 1999, 129, 1656–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Xu, J.; Zhang, Z.; Huang, X.; Zhang, A.; Wang, J.; Zheng, Q.; Fu, L.; Du, J. Association study between methylenetetrahydrofolate reductase polymorphisms and unexplained recurrent pregnancy loss: A meta-analysis. Gene 2013, 514, 105–111. [Google Scholar] [CrossRef]
- Wu, W.; Shen, O.; Qin, Y.; Lu, J.; Niu, X.; Zhou, Z.; Lu, C.; Xia, Y.; Wang, S.; Wang, X. Methylenetetrahydrofolate reductase C677T polymorphism and the risk of male infertility: A meta-analysis. Int. J. Androl. 2011, 35, 18–24. [Google Scholar] [CrossRef]
- Kang, S.; Zhao, X.; Liu, L.; Wu, W.; Zhang, N. Association of the C677T Polymorphism in the MTHFR Gene with Hemorrhagic Stroke: A Meta-Analysis. Genet. Test. Mol. Biomark. 2013, 17, 412–417. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Wu, X.; Zhi, X.; Zheng, Q.; Sun, G. Methylenetetrahydrofolate reductase C677T polymorphism and type 2 diabetes mellitus in Chinese population: A me-ta-analysis of 29 case-control studies. PLoS ONE 2014, 9, e102443. [Google Scholar] [CrossRef]
- Chang, W.-W.; Zhang, L.; Yao, Y.-S.; Su, H.; Jin, Y.-L.; Chen, Y. Methylenetetrahydrofolate reductase (MTHFR) C677T polymorphism and susceptibility to diabetic nephropathy in Chinese type 2 diabetic patients: A meta-analysis. Ren. Fail. 2013, 35, 1038–1043. [Google Scholar] [CrossRef] [Green Version]
- Karabacak, E.; Aydin, E.; Ozcan, O.; Dogan, B.; Gultepe, M.; Cosar, A.; Muftuoglu, T. Methylenetetrahydrofolate reductase (MTHFR) 677C>T gene polymorphism as a possible factor for reducing clini-cal severity of psoriasis. Int. J. Clin. Exp. Med. 2014, 7, 697–702. [Google Scholar]
- Peng, Q.; Lao, X.; Huang, X.; Qin, X.; Li, S.; Zeng, Z. The MTHFR C677T polymorphism contributes to increased risk of Alzheimer’s Disease: Evidence based on 40 case-control studies. Neurosci. Lett. 2015, 586, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.; Shmorgun, D.; Chan, W. Common C677T Polymorphism of the Methylenetetrahydrofolate Reductase Gene and the Risk of Venous Thromboembolism: Meta-Analysis of 31 Studies. Pathophysiol. Haemost. Thromb. 2002, 32, 51–58. [Google Scholar] [CrossRef] [PubMed]
- McColgan, P.; Sharma, P. The Genetics of Carotid Dissection: Meta-Analysis of a MTHFR/C677T Common Molecular Variant. Cereb. Dis. 2008, 25, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.-Y.; Qian, Z.-Z.; Gong, F.-F.; Lu, S.-S.; Feng, F.; Wu, Y.-L.; Yang, H.-Y.; Sun, Y.-H. Methylenetetrahydrofolate reductase (MTHFR) polymorphism susceptibility to schizophrenia and bipolar disorder: An updated meta-analysis. J. Neural. Transm. 2014, 122, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Liew, S.C.; Gupta, E.D. Methylenetetrahydrofolate reductase (MTHFR) C677T polymorphism: Epidemiology, metabolism and the associated diseases. Eur. J. Med. Genet. 2015, 58, 1–10. [Google Scholar] [CrossRef]
- Bolufer, P.; Barragán, E.; Collado, M.; Cervera, J.; Lopez, J.-A.; Sanz, M.A. Influence of genetic polymorphisms on the risk of developing leukemia and on disease progression. Leuk. Res. 2006, 30, 1471–1491. [Google Scholar] [CrossRef]
- Zanrosso, C.W.; Hatagima, A.; Emerenciano, M.; Ramos, F.; Figueiredo, A.; Félix, T.M.; Segal, S.L.; Giugliani, R.; Muniz, M.T.C.; Pombo-De-Oliveira, M.S. The role of methylenetetrahydrofolate reductase in acute lymphoblastic leukemia in a Brazilian mixed population. Leuk. Res. 2006, 30, 477–481. [Google Scholar] [CrossRef]
- Yan, J.; Yin, M.; Dreyer, Z.E.; Scheurer, M.E.; Kamdar, K.; Wei, Q.; Okcu, M.F. A meta-analysis of MTHFR C677T and A1298C polymorphisms and risk of acute lymphoblastic leukemia in children. Pe-Diatr Blood Cancer 2012, 58, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Kurzwelly, D.; Knop, S.; Guenther, M.; Loeffler, J.; Korfel, A.; Thiel, E.; Hebart, H.; Simon, M.; Weller, M.; Linnebank, M.; et al. Genetic variants of folate and methionine metabolism and PCNSL incidence in a German patient population. J. Neuro-Oncol. 2010, 100, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.N.; Lee, I.; Kim, Y.; Tran, H.T.T.; Yang, D.; Lee, J.; Shin, M.; Park, K.; Shin, M.; Choi, J.; et al. Association between folate-metabolizing pathway polymorphism and non-Hodgkin lymphoma. Br. J. Haematol. 2008, 140, 287–294. [Google Scholar] [CrossRef]
- Tang, M.; Wang, S.-Q.; Liu, B.-J.; Cao, Q.; Li, B.-J.; Li, P.-C.; Li, Y.-F.; Qin, C.; Zhang, W. The methylenetetrahydrofolate reductase (MTHFR) C677T polymorphism and tumor risk: Evidence from 134 case–control studies. Mol. Biol. Rep. 2014, 41, 4659–4673. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.-Y.; Yang, S.-J.; Zhang, J.-X.; Chen, X.-Y. Methylenetetrahydrofolate reductase genetic polymorphisms and esophageal squamous cell carcinoma susceptibility: A meta-analysis of case-control studies. Asian Pac. J. Cancer Prev. 2013, 14, 21–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Xiao, F.; An, Z.; Hao, L. Systematic review on the relationship between genetic polymorphisms of methylenetetrahydrofolate reductase and esophageal squamous cell carcinoma. Asian Pac. J. Cancer Prev. 2011, 12, 1861–1866. [Google Scholar] [PubMed]
- Boccia, S.; Hung, R.; Ricciardi, G.; Gianfagna, F.; Ebert, M.P.A.; Fang, J.-Y.; Gao, C.-M.; Götze, T.; Graziano, F.; Lacasaña-Navarro, M.; et al. Meta- and Pooled Analyses of the Methylenetetrahydrofolate Reductase C677T and A1298C Polymorphisms and Gastric Cancer Risk: A Huge-GSEC Review. Am. J. Epidemiol. 2007, 167, 505–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulvik, A.; Evensen, E.T.; Lien, E.A.; Hoff, G.; Vollset, S.E.; Majak, B.M.; Ueland, P.M. Smoking, folate and methylenetetrahydrofolate reductase status as interactive determinants of adenomatous and hyper-plastic polyps of colorectum. Am. J. Med. Genet. 2001, 101, 246–254. [Google Scholar] [CrossRef]
- Friso, S.; Choi, S.-W.; Girelli, D.; Mason, J.B.; Dolnikowski, G.G.; Bagley, P.J.; Olivieri, O.; Jacques, P.F.; Rosenberg, I.H.; Corrocher, R.; et al. A common mutation in the 5,10-methylenetetrahydrofolate reductase gene affects genomic DNA methylation through an interaction with folate status. Proc. Natl. Acad. Sci. USA 2002, 99, 5606–5611. [Google Scholar] [CrossRef] [Green Version]
- Stern, L.L.; Mason, J.B.; Selhub, J.; Choi, S.W. Genomic DNA hypomethylation, a characteristic of most cancers, is present in peripheral leukocytes of individuals who are homozygous for the C677T polymorphism in the methylenetetrahydrofolate reductase gene. Cancer Epidemiol. Biomark. Prev. 2000, 9, 849–853. [Google Scholar]
- Quinlivan, E.P.; Davis, S.R.; Shelnutt, K.P.; Henderson, G.N.; Ghandour, H.; Shane, B.; Selhub, J.; Bailey, L.B.; Stacpoole, P.W.; Gregory, J.F. Methylenetetrahydrofolate Reductase 677C→T Polymorphism and Folate Status Affect One-Carbon Incorporation into Human DNA Deoxynucleosides. J. Nutr. 2005, 135, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Cecilio, A.P.; Takakura, E.T.; Jumes, J.J.; Dos Santos, J.W.; Herrera, A.C.; Victorino, V.J.; Panis, C. Breast cancer in Brazil: Epidemiology and treatment challenges. Breast Cancer Dove Med. Press 2015, 7, 43–49. [Google Scholar]
- Ciriello, G.; Sinha, R.; Hoadley, K.A.; Jacobsen, A.S.; Reva, B.; Perou, C.M.; Sander, C.; Schultz, N. The molecular diversity of Luminal A breast tumors. Breast Cancer Res. Treat. 2013, 141, 409–420. [Google Scholar] [CrossRef] [Green Version]
- Network, C.G.A. Comprehensive molecular portraits of human breast tumours. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Goldhirsch, A.; Winer, E.P.; Coates, A.S.; Gelber, R.D.; Piccart-Gebhart, M.; Thürlimann, B.; Senn, H.-J.; Albain, K.S.; André, F.; Bergh, J.; et al. Personalizing the treatment of women with early breast cancer: Highlights of the St Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer. Ann. Oncol. 2013, 24, 2206–2223. [Google Scholar] [CrossRef]
- Petrone, I.; Rodrigues, F.R.; Fernandes, P.V.; Abdelhay, E. Immunohistochemical Biomarkers in Ductal Carcinoma in Situ. Open J. Pathol. 2020, 10, 129–146. [Google Scholar] [CrossRef]
- Kalemi, T.G.; Lambropoulos, A.F.; Gueorguiev, M.; Chrisafi, S.; Papazisis, K.T.; Kotsis, A. The association of p53 mutations and p53 codon 72, Her 2 codon 655 and MTHFR C677T polymorphisms with breast cancer in Northern Greece. Cancer Lett. 2005, 222, 57–65. [Google Scholar] [CrossRef]
- Larsson, S.C.; Giovannucci, E.; Wolk, A. Folate and Risk of Breast Cancer: A Meta-analysis. J. Natl. Cancer Inst. 2007, 99, 64–76. [Google Scholar] [CrossRef] [Green Version]
- Lu, Q.; Jiang, K.; Li, Q.; Ji, Y.-J.; Chen, W.-L.; Xue, X.-H. Polymorphisms in the MTHFR gene are associated with breast cancer risk and prognosis in a Chinese population. Tumor Biol. 2015, 36, 3757–3762. [Google Scholar] [CrossRef]
- Awwad, N.; Yousef, A.-M.; Abuhaliema, A.; Abdalla, I.; Yousef, M. Relationship between Genetic Polymorphisms in MTHFR (C677T, A1298C and their Haplotypes) and the Incidence Of Breast Cancer among Jordanian Females—Case-Control Study. Asian Pac. J. Cancer Prev. 2015, 16, 5007–5011. [Google Scholar] [CrossRef] [Green Version]
- Hesari, A.; MalekSabet, A.; Tirkani, A.N.; Ghazizadeh, H.; Iranifar, E.; Mohagheg, F.; Anoshrvani, A.A.; Ghasemi, F. Evaluation of the two polymorphisms rs1801133 in MTHFR and rs10811661 in CDKN2A/B in breast cancer. J. Cell. Biochem. 2019, 120, 2090–2097. [Google Scholar] [CrossRef]
- Gao, C.-M.; Tang, J.-H.; Cao, H.-X.; Ding, J.-H.; Wu, J.-Z.; Wang, J.; Liu, Y.-T.; Li, S.-P.; Su, P.; Matsuo, K.; et al. MTHFR polymorphisms, dietary folate intake and breast cancer risk in Chinese women. J. Hum. Genet. 2009, 54, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Sohn, K.-J.; Croxford, R.; Yates, Z.; Lucock, M.; Kim, Y.-I. Effect of the methylenetetrahydrofolate reductase C677T polymorphism on chemosensitivity of colon and breast cancer cells to 5-fluorouracil and methotrexate. J. Natl. Cancer Inst. 2004, 96, 134–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrubsole, M.J.; Jin, F.; Dai, Q.; Shu, X.O.; Potter, J.D.; Hebert, J.R.; Gao, Y.T.; Zheng, W. Dietary folate intake and breast cancer risk: Results from the Shanghai Breast Cancer Study. Cancer Res. 2001, 61, 190–196. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Shen, Y. MTHFR C677T polymorphism and breast, ovarian cancer risk: A meta-analysis of 19,260 patients and 26,364 controls. Onco. Targets Ther. 2017, 10, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Pooja, S.; Carlus, J.; Sekhar, D.; Francis, A.; Gupta, N.; Konwar, R.; Kumar, S.; Kumar, S.; Thangaraj, K.; Rajender, S. MTHFR 677C>T Polymorphism and the Risk of Breast Cancer: Evidence from an Original Study and Pooled Data for 28031 Cases and 31880 Controls. PLoS ONE 2015, 10, e0120654. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, W.; Dong, X. Association of 677 C>T (rs1801133) and 1298 A>C (rs1801131) Polymorphisms in the MTHFR Gene and Breast Cancer Susceptibility: A Meta-Analysis Based on 57 Individual Studies. PLoS ONE 2014, 9, e71290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meneses-Sanchez, P.; Garcia-Hernandez, S.C.; Porchia, L.M.; Pérez-Fuentes, R.; Torres-Rasgado, E.; Soto, A.D.A.; Gonzalez-Mejia, M.E. C677T and A1298C methylenetetrahydrofolate reductase polymorphisms and breast cancer susceptibility among Latinos: A meta-analysis. Breast Cancer 2019, 26, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Zara-Lopes, T.; Gimenez-Martins, A.; Nascimento-Filho, C.; Castanhole-Nunes, M.; Galbiatti-Dias, A.; Padovani-Júnior, J.; Maniglia, J.; Francisco, J.; Pavarino, E.; Goloni-Bertollo, E.; et al. Role of MTHFR C677T and MTR A2756G polymorphisms in thyroid and breast cancer development. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Pu, Y.; Wu, Y.; Qin, R.; Zhang, Q.; Sun, Y.; Zheng, W.; Chen, L. Association between dietary intake of folate and MTHFR and MTR genotype with risk of breast cancer. Genet. Mol. Res. 2014, 13, 8925–8931. [Google Scholar] [CrossRef]
- Jiang-Hua, Q.; De-Chuang, J.; Zhen-Duo, L.; Shu-De, C.; Zhenzhen, L. Association of methylenetetrahydrofolate reductase and methionine synthase polymorphisms with breast cancer risk and interaction with folate, vitamin B6, and vitamin B12 intakes. Tumor Biol. 2014, 35, 11895–11901. [Google Scholar] [CrossRef] [PubMed]
- Akilzhanova, A.; Nurkina, Z.; Momynaliev, K.; Ramanculov, E.; Zhumadilov, Z.; Zhumadilov, Z.; Rakhypbekov, T.; Hayashida, N.; Nakashima, M.; Takamura, N. Genetic profile and determinants of homocysteine levels in Kazakhstan patients with breast cancer. Anticancer. Res. 2013, 33, 4049–4059. [Google Scholar]
- Diakite, B.; Tazzite, A.; Hamzi, K.; Jouhadi, H.; Nadifi, S. Methylenetetrahydrofolate Reductase C677T polymorphism and breast cancer risk in Moroccan women. Afr. Health Sci. 2012, 12, 204–209. [Google Scholar] [CrossRef]
- Rezende, L.M.; Marson, F.A.L.; Lima, C.S.P.; Bertuzzo, C. Can MTHFR C677T and A1298C Polymorphisms Alter the Risk and Severity of Sporadic Breast Cancer in Bra-zilian Women? Clin. Breast Cancer 2017, 17, e199–e208. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, Y.; Li, R.; Han, X.; Ma, Y.; Liu, B.; Kong, X. Association of mthfr A1298C polymorphism with breast cancer and/or ovarian cancer risk: An updated me-ta-analysis. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 72–86. [Google Scholar]
- Durán, M.; Ángel, C.; Araújo, M.D.; Pinheiro, D.D.R.; Burbano, R.M.R.; Borges, B.D.N. Thymidylate synthase and methylenetetrahydrofolate reductase polymorphisms and breast cancer susceptibility in a Brazilian population. Meta Gene. 2021, 28, 100889. [Google Scholar] [CrossRef]
- Castiglia, P.; Sanna, V.; Azara, A.; De Miglio, M.R.; Murgia, L.; Pira, G.; Sanges, F.; Fancellu, A.; Carru, C.; Bisail, M.; et al. Methylenetetrahydrofolate reductase (MTHFR) C677T and A1298C polymorphisms in breast cancer: A Sardinian preliminary case-control study. Int. J. Med. Sci. 2019, 16, 1089–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houghton, S.C.; Eliassen, A.H.; Zhang, S.M.; Selhub, J.; Rosner, B.A.; Willett, W.C.; Hankinson, S.E. Plasma B-vitamins and one-carbon metabolites and the risk of breast cancer in younger women. Breast Cancer Res. Treat. 2019, 176, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Omran, M.H.; E Fotouh, B.; Shousha, W.G.; Ismail, A.; Ibrahim, N.E.; Ramadan, S.S. Strong Correlation of MTHFR Gene Polymorphisms with Breast Cancer and its Prognostic Clinical Factors among Egyptian Females. Asian Pac. J. Cancer Prev. 2021, 22, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Ajaz, S.; Ali, S.M.; Siddiqa, A.; Memon, M.A.; Abid, A.; Khaliq, S. Independent and Combined Associations of 677C/T and 1298A/C Polymorphisms in the MTHFR gene with Breast Cancers in a South-Asian Population. Medrxiv 2021. [Google Scholar] [CrossRef]
- Miranda-Filho, A.; Piñeros, M.; Soerjomataram, I.; Deltour, I.; Bray, F. Cancers of the brain and CNS: Global patterns and trends in incidence. Neuro-oncology 2016, 19, 166–280. [Google Scholar] [CrossRef]
- Philips, A.; Henshaw, D.; Lamburn, G.; O’Carroll, M. Brain Tumours: Rise in Glioblastoma Multiforme Incidence in England 1995–2015 Suggests an Adverse Environ-mental or Lifestyle Factor. J. Environ. Public Health 2018, 2018, 7910754. [Google Scholar] [CrossRef] [Green Version]
- Bell, J.S.; Koffie, R.M.; Rattani, A.; Dewan, M.C.; Baticulon, R.E.; Qureshi, M.M.; Wahjoepramono, E.J.; Rosseau, G.; Park, K.; Nahed, B.V. Global incidence of brain and spinal tumors by geographic region and income level based on cancer registry data. J. Clin. Neurosci. 2019, 66, 121–127. [Google Scholar] [CrossRef]
- Leece, R.; Xu, J.; Ostrom, Q.T.; Chen, Y.; Kruchko, C.; Barnholtz-Sloan, J.S. Global incidence of malignant brain and other central nervous system tumors by histology, 2003–2007. Neuro-Oncology 2017, 19, 1553–1564. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Cioffi, G.; Gittleman, H.; Patil, N.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2012–2016. Neuro-Oncology 2019, 21, v1–v100. [Google Scholar] [CrossRef]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO Classification of Tumours of the Central Nervous System. Acta Neuropathol. 2007, 114, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, D.N.; Perry, A.; Reifenberger, G.; Von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izmirli, M. A literature review of MTHFR (C677T and A1298C polymorphisms) and cancer risk. Mol. Biol. Rep. 2012, 40, 625–637. [Google Scholar] [CrossRef]
- Davis, C.D.; Uthus, E.O. DNA Methylation, Cancer Susceptibility, and Nutrient Interactions. Exp. Biol. Med. 2004, 229, 988–995. [Google Scholar] [CrossRef]
- Kulis, M.; Esteller, M. DNA Methylation and Cancer. Adv. Genet. 2010, 70, 27–56. [Google Scholar] [CrossRef]
- Van Tongelen, A.; Loriot, A.; De Smet, C. Oncogenic roles of DNA hypomethylation through the activation of cancer-germline genes. Cancer Lett. 2017, 396, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Sirachainan, N.; Wongruangsri, S.; Kajanachumpol, S.; Pakakasama, S.; Visudtibhan, A.; Nuchprayoon, I.; Lusawat, A.; Phudhicharoenrat, S.; Shuangshoti, S.; Hongeng, S. Folate pathway genetic polymorphisms and susceptibility of central nervous system tumors in Thai children. Cancer Detect. Prev. 2008, 32, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Salnikova, L.E.; Belopolskaya, O.B.; Zelinskaya, N.I.; Rubanovich, A.V. The potential effect of gender in CYP1A1 and GSTM1 genotype-specific associations with pediatric brain tumor. Tumor Biol. 2013, 34, 2709–2719. [Google Scholar] [CrossRef] [PubMed]
- Greenop, K.R.; Miller, M.; De Klerk, N.H.; Scott, R.J.; Attia, J.; Ashton, L.J.; Dalla-Pozza, L.; Bower, C.; Armstrong, B.K.; Milne, E. Maternal Dietary Intake of Folate and Vitamins B6 and B12 During Pregnancy and Risk of Childhood Brain Tumors. Nutr. Cancer 2014, 66, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Chiavarini, M.; Naldini, G.; Fabiani, R. Maternal Folate Intake and Risk of Childhood Brain and Spinal Cord Tumors: A System-atic Review and Meta-Analysis. Neuroepidemiology 2018, 51, 82–95. [Google Scholar] [CrossRef]
- Bunin, G.R.; Kuijten, R.R.; Buckley, J.D.; Rorke, L.B.; Meadows, A.T. Relation between Maternal Diet and Subsequent Primitive Neuroectodermal Brain Tumors in Young Children. N. Engl. J. Med. 1993, 329, 536–541. [Google Scholar] [CrossRef]
- Sirikulchayanonta, C.; Madjupa, K.; Chongsuwat, R.; Pandii, W. Do Thai women of child bearing age need pre-conceptional supplementation of dietary folate? Asia Pac. J. Clin. Nutr. 2004, 13, 69–73. [Google Scholar]
- Sanghvi, T.G.; Harvey, P.W.; Wainwright, E. Maternal iron-folic acid supplementation programs: Evidence of impact and imple-mentation. Food Nutr. Bull. 2010, 31 (Suppl. S2), S100–S107. [Google Scholar] [CrossRef] [PubMed]
- Lucock, M.; Yates, Z. Folic acid—Vitamin and panacea or genetic time bomb? Nat. Rev. Genet. 2005, 6, 235–240. [Google Scholar] [CrossRef]
- Kumawat, R.; Gowda, S.H.; Debnath, E.; Rashid, S.; Niwas, R.; Gupta, Y.; Upadaya, A.D.; Suri, A.; Chandra, P.S.; Gupta, D.K.; et al. Association of Single Nucleotide Polymorphisms (SNPs) in Genes Encoding for Folate Metabolising Enzymes with Glioma and Meningioma in Indian Population. Asian Pac. J. Cancer Prev. 2018, 19, 3415–3425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cartron, P.-F.; Hervouet, E.; Debien, E.; Olivier, C.; Pouliquen, D.; Menanteau, J.; Loussouarn, D.; Martin, S.A.; Campone, M.; Vallette, F.M. Folate supplementation limits the tumourigenesis in rodent models of gliomagenesis. Eur. J. Cancer 2012, 48, 2431–2441. [Google Scholar] [CrossRef] [PubMed]
- Lubecka-Pietruszewska, K.; Kaufman-Szymczyk, A.; Stefanska, B.; Fabianowska-Majewska, K. Folic acid enforces DNA methylation-mediated transcriptional silencing of PTEN, APC and RARbeta2 tumour suppressor genes in breast cancer. Biochem. Biophys. Res. Commun. 2013, 430, 623–628. [Google Scholar] [CrossRef]
- Zgheib, R.; Battaglia-Hsu, S.-F.; Hergalant, S.; Quéré, M.; Alberto, J.-M.; Chéry, C.; Rouyer, P.; Gauchotte, G.; Guéant, J.-L.; Namour, F. Folate can promote the methionine-dependent reprogramming of glioblastoma cells towards pluripotency. Cell Death Dis. 2019, 10, 596. [Google Scholar] [CrossRef] [Green Version]
- Semmler, A.; Simon, M.; Moskau, S.; Linnebank, M. The Methionine Synthase Polymorphism c.2756A>G Alters Susceptibility to Glioblastoma Multiforme. Cancer Epidemiol. Biomark. Prev. 2006, 15, 2314–2316. [Google Scholar] [CrossRef] [Green Version]
- Pandith, A.A.; Qasim, I.; Zahoor, W.; Shah, P.; Bhat, A.R. ACE I/D sequence variants but not MTHFR C677T, is strongly linked to malignant glioma risk and its variant DD genotype may act as a promising predictive biomarker for overall survival of glioma patients. Gene 2018, 639, 62–68. [Google Scholar] [CrossRef]
- Bethke, L.; Webb, E.; Murray, A.; Schoemaker, M.; Johansen, C.; Christensen, H.C.; Muir, K.; McKinney, P.; Hepworth, S.; Dimitropoulou, P.; et al. Comprehensive analysis of the role of DNA repair gene polymorphisms on risk of glioma. Hum. Mol. Genet. 2007, 17, 800–805. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, D.M.; de Lima, G.P.V.; Faria, M.H.G.; Rabenhorst, S.H.B. Polymorphisms of folate pathway enzymes (methylenetetrahydrofolate reductase and thymidylate synthase) and their relationship with thymidylate synthase expression in human astrocytic tumors. DNA Cell Biol. 2012, 31, 57–66. [Google Scholar] [CrossRef]
- Kafadar, A.M.; Yilmaz, H.; Kafadar, D.; Ergen, A.; Zeybek, U.; Bozkurt, N.; Kuday, C.; Isbir, T. C677T gene polymorphism of methylenetetrahydrofolate reductase (MTHFR) in meningiomas and high-grade gli-omas. Anticancer Res. 2006, 26, 2445–2449. [Google Scholar] [PubMed]
- Lu, Q.; Dai, D.; Zhao, W.; Wang, L.; Yue, Z.; Chen, X.; Han, G.; Hao, B.; Yang, P.; Deng, A.; et al. Association between MTHFR 677C>T polymorphism and risk of gliomas: Evidence from a meta-analysis. Tumor Biol. 2013, 34, 2801–2807. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Yuan, L.; Tian, H.; Cao, H.; Chen, S. Association of the MTHFR C677T polymorphism with primary brain tumor risk. Tumor Biol. 2013, 34, 3457–3464. [Google Scholar] [CrossRef]
- Chen, D.; Dong, J.; Huang, Y.; Gao, F.; Yang, X.; Gong, X.; Lv, X.; Chu, C.; Wu, Y.; Zheng, Y. Folate metabolism genetic polymorphisms and meningioma and glioma susceptibility in adults. Oncotarget 2017, 8, 57265–57277. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, Y.-W.; Shi, H.-P.; Wang, Y.-Z.; Li, G.-L.; Yu, H.-T.; Xie, X.-Y. 5,10-Methylenetetrahydrofolate reductase (MTHFR), methionine synthase (MTRR), and methionine synthase reductase (MTR) gene polymorphisms and adult meningioma risk. J. Neuro-Oncol. 2013, 115, 233–239. [Google Scholar] [CrossRef]
- Li, R.; Wang, R.; Li, Y.; Li, X.; Feng, Y.; Li, Y.; Jiang, C. Association study on MTHFR polymorphisms and meningioma in northern China. Gene 2013, 516, 291–293. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Liu, W.; Yu, X.; Wang, L.; Shao, L.; Yi, W. Risk association of meningiomas with MTHFR C677T and GSTs polymorphisms: A meta-analysis. Int. J. Clin. Exp. Med. 2014, 7, 3904–3914. [Google Scholar]
- Han, X.-Y.; Wang, W.; Wang, L.-L.; Wang, X.-R.; Li, G. Genetic variants and increased risk of meningioma: An updated meta-analysis. Onco.Targets Ther. 2017, 10, 1875–1888. [Google Scholar] [CrossRef] [Green Version]
- Linnebank, M.; Semmler, A.; Moskau, S.; Smulders, Y.; Blom, H.; Simon, M. The methylenetetrahydrofolate reductase (MTHFR) variant c.677C>T (A222V) influences overall survival of pa-tients with glioblastoma multiforme. Neuro Oncol. 2008, 10, 548–552. [Google Scholar] [CrossRef] [Green Version]
- Cadieux, B.; Ching, T.-T.; Vandenberg, S.R.; Costello, J.F. Genome-wide Hypomethylation in Human Glioblastomas Associated with Specific Copy Number Alteration, Methylenetetrahydrofolate Reductase Allele Status, and Increased Proliferation. Cancer Res. 2006, 66, 8469–8476. [Google Scholar] [CrossRef] [Green Version]
- Faria, G.M.; Soares, I.D.P.; Salazar, M.D.; Amorim, M.R.; Pessoa, B.L.; Da Fonseca, C.O.; Quirico-Santos, T. Intranasal perillyl alcohol therapy improves survival of patients with recurrent glioblastoma harboring mutant variant for MTHFR rs1801133 polymorphism. BMC Cancer 2020, 20, 1–10. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 coun-tries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagini, S. Carcinoma of the stomach: A review of epidemiology, pathogenesis, molecular genetics and chemoprevention. World J. Gastrointest. Oncol. 2012, 4, 156–169. [Google Scholar] [CrossRef]
- Rawla, P.; Barsouk, A. Epidemiology of gastric cancer: Global trends, risk factors and prevention. Gastroenterol. Rev. 2019, 14, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Karimi, P.; Islami, F.; Anandasabapathy, S.; Freedman, N.D.; Kamangar, F. Gastric Cancer: Descriptive Epidemiology, Risk Factors, Screening, and Prevention. Cancer Epidemiol. Biomark. Prev. 2014, 23, 700–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cisło, M.; Filip, A.A.; Offerhaus, G.J.A.; Ciseł, B.; Rawicz-Pruszyński, K.; Skierucha, M.; Polkowski, W.P. Distinct molecular subtypes of gastric cancer: From Laurén to molecular pathology. Oncotarget 2018, 9, 19427–19442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauren, P. The two histological main types of gastric carcinoma: Diffuse and so-called intes-tinal-type carcinoma. An at-tempt at a histo-clinical classification. Acta Pathol. Microbiol. Scand 1965, 64, 31–49. [Google Scholar] [CrossRef]
- Henson, D.E.; Dittus, C.; Younes, M.; Nguyen, H.; Albores-Saavedra, J. Differential trends in the intestinal and diffuse types of gastric carcinoma in the United States, 1973–2000: Increase in the signet ring cell type. Arch. Pathol. Lab. Med. 2004, 128, 765–770. [Google Scholar] [CrossRef]
- Network, C.G.A.R. Comprehensive molecular characterization of gastric adenocarcinoma. Nature 2014, 513, 202–209. [Google Scholar]
- Cristescu, R.; Lee, J.; Nebozhyn, M.; Kim, K.-M.; Ting, J.C.; Wong, S.S.; Liu, J.; Yue, Y.G.; Wang, J.; Yu, K.; et al. Molecular analysis of gastric cancer identifies subtypes associated with distinct clinical outcomes. Nat. Med. 2015, 21, 449–456. [Google Scholar] [CrossRef]
- Öksüz, E.; Görgişen, G.; Oto, G.; Özdemir, H.; Aras, A.; Öksüz, M.; Gülaçar, İ.M.; Demirkol, M.H. Relationship between MTHFR Gene Polymorphisms and Gastrointestinal Tumors Development: Perspective from Eastern Part of Turkey. J. Invest. Surg. 2020, 1–9. [Google Scholar] [CrossRef]
- Han, Z.; Sheng, H.; Gao, K.; Fan, Y.; Xie, X. Associations of the the MTHFR rs1801133 polymorphism with gastric cancer risk in the Chinese Han population. Biomed Rep. 2021, 14, 14. [Google Scholar] [CrossRef]
- Gao, C.; Wu, J.; Ding, J.; Liu, Y.; Zang, Y.; Li, S.; Su, P.; Hu, X.; Xu, T.; Toshiro, T.; et al. Polymorphisms of methylenetetrahydrofolate reductase C677T and the risk of stomach cancer. Zhonghua Liuxingbingxue Zazhi 2002, 23, 289–292. [Google Scholar] [PubMed]
- Chen, L.; Lu, N.; Zhang, B.-H.; Weng, L.; Lu, J. Association between the MTHFR C677T polymorphism and gastric cancer susceptibility: A meta-analysis of 5757 cases and 8501 controls. Oncol. Lett. 2015, 10, 1159–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zintzaras, E. Association of methylenetetrahydrofolate reductase (MTHFR) polymorphisms with genetic susceptibility to gastric cancer: A meta-analysis. J. Hum. Genet. 2006, 51, 618–624. [Google Scholar] [CrossRef] [Green Version]
- Mazzuca, F.; Borro, M.; Botticelli, A.; Aimati, L.; Gentile, G.; Capalbo, C.; Maddalena, C.; Mazzotti, E.; Simmaco, M.; Marchetti, P. Effect of MTHFR Polymorphisms on Gastrointestinal Cancer Risk in Italy. World J. Oncol. 2015, 6, 394–397. [Google Scholar] [CrossRef]
- Dong, L.M.; Potter, J.D.; White, E.; Ulrich, C.M.; Cardon, L.R.; Peters, U. Genetic susceptibility to cancer: The role of polymorphisms in candidate genes. JAMA 2008, 299, 2423–2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, L.; Wang, P.; Sun, B.; Chen, G. The polymorphism of methylenetetrahydrofolate reductase C677T but not A1298C contributes to gastric cancer. Tumor Biol. 2013, 35, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Wu, J.; Liang, P.; Li, J.; Yuan, L.; Liu, X. Methylenetetrahydrofolate Reductase C677T and A1298C Polymorphisms and Gastric Cancer: A Meta-analysis. Arch. Med. Res. 2010, 41, 125–133. [Google Scholar] [CrossRef]
- Hosseini-Asl, S.S.; Pourfarzi, F.; Barzegar, A.; Mazani, M.; Farahmand, N.; Niasti, E.; Yazdanbod, A.; Didevar, R.; Akhavan, H.; Malekzadeh, R. Decrease in gastric cancer susceptibility by MTHFR C677T polymorphism in Ardabil Province, Iran. Turk. J. Gastroenterol. 2013, 24, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.-H.; Shin, M.-H.; Kweon, S.-S.; Kim, H.N.; Song, H.-R.; Piao, J.-M.; Choi, J.-S.; Shim, H.J.; Hwang, J.E.; Kim, H.-R.; et al. Methylenetetrahydrofolate reductase C677T polymorphism in patients with gastric and colorectal cancer in a Korean population. BMC Cancer 2010, 10, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galván-Portillo, M.V.; Cantoral, A.; Oñate-Ocaña, L.F.; Chen, J.; Herrera-Goepfert, R.; Torres-Sanchez, L.; Hernandez-Ramirez, R.U.; Palma-Coca, O.; López-Carrillo, L. Gastric cancer in relation to the intake of nutrients involved in one-carbon metabolism among MTHFR 677 TT carriers. Eur. J. Nutr. 2009, 48, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Yu, S.; Jiang, H.; Li, W.; Xu, X.; Cheng, X.; Peng, K.; Chen, E.; Cui, Y.; Liu, T. A Meta-Analysis: Methylenetetrahydrofolate Reductase C677T Polymorphism in Gastric Cancer Patients Treated with 5-Fu Based Chemotherapy Predicts Serious Hematologic Toxicity but Not Prognosis. J. Cancer 2018, 9, 1057–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wöhrer, S.S.; Raderer, M.; Hejna, M. Palliative chemotherapy for advanced gastric cancer. Ann. Oncol. 2004, 15, 1585–1595. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, X.; Kong, X. MTHFR C677T Polymorphism is Associated with Tumor Response to Preoperative Chemoradiotherapy: A Result Based on Previous Reports. Med. Sci. Monit. 2015, 21, 3068–3076. [Google Scholar] [CrossRef] [Green Version]
- Meulendijks, D.; De Groot, J.W.B.; Los, M.; Boers, J.E.; Beerepoot, L.V.; Polee, M.B.; Beeker, A.; Portielje, J.E.; Goey, S.H.; De Jong, R.S.; et al. Bevacizumab combined with docetaxel, oxaliplatin, and capecitabine, followed by maintenance with capecitabine and bevacizumab, as first-line treatment of patients with advanced HER2-negative gastric cancer: A multicenter phase 2 study. Cancer 2016, 122, 1434–1443. [Google Scholar] [CrossRef] [Green Version]
- Blank, S.; Rachakonda, S.; Keller, G.; Weichert, W.; Lordick, F.; Langer, R.; Springfeld, C.; Bruckner, T.; Becker, K.; Kumar, R.; et al. A retrospective comparative exploratory study on two Methylentetrahydrofolate Reductase (MTHFR) polymorphisms in esophagogastric cancer: The A1298C MTHFR polymorphism is an independent prognostic factor only in neoadjuvantly treated gastric cancer patients. BMC Cancer 2014, 14, 58. [Google Scholar] [CrossRef] [Green Version]
- Kong, S.; Ye, F.; Dang, Y.; Hua, Y.; Zhang, G. Association of MTHFR C677T polymorphism with severity and localization of chronic atrophic gastritis patients without Helicobacter pylori infection: A case control study. BMC Cancer 2020, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Saberi, S.; Zendehdel, K.; Jahangiri, S.; Talebkhan, Y.; Abdirad, A.; Mohajerani, N.; Bababeik, M.; Karami, N.; Esmaili, M.; Oghalaie, A.; et al. Impact of Methylenetetrahydrofolate Reductase C677T Polymorphism on the Risk of Gastric Cancer and Its Interaction with Helicobacter pylori Infection. Iran. Biomed. J. 2012, 16, 179–184. [Google Scholar]
- Wu, X.; Peng, B.; Qian, K.; Zhang, W.; Min, J.; Zhang, M.; Zeng, F.; Wang, Z. The combination of methylenehydrofolate reductase C677T polymorphism screening and gastrointestinal tumor markers detection may be an early screening method for gastrointestinal cancer related to helicobacter pylori infection. Genes Dis. 2021, 8. [Google Scholar] [CrossRef]
- Kakkoura, M.G.; Sokratous, K.; Demetriou, C.A.; Loizidou, M.A.; Loucaides, G.; Kakouri, E.; Hadjisavvas, A.; Kyriacou, K. Mediterranean diet-gene interactions: A targeted metabolomics study in Greek-Cypriot women. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Shiao, S.P.K.; Grayson, J.; Yu, C.H.; Wasek, B.; Bottiglieri, T. Gene Environment Interactions and Predictors of Colorectal Cancer in Family-Based, Multi-Ethnic Groups. J. Pers. Med. 2018, 8, 10. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrone, I.; Bernardo, P.S.; dos Santos, E.C.; Abdelhay, E. MTHFR C677T and A1298C Polymorphisms in Breast Cancer, Gliomas and Gastric Cancer: A Review. Genes 2021, 12, 587. https://doi.org/10.3390/genes12040587
Petrone I, Bernardo PS, dos Santos EC, Abdelhay E. MTHFR C677T and A1298C Polymorphisms in Breast Cancer, Gliomas and Gastric Cancer: A Review. Genes. 2021; 12(4):587. https://doi.org/10.3390/genes12040587
Chicago/Turabian StylePetrone, Igor, Paula Sabbo Bernardo, Everton Cruz dos Santos, and Eliana Abdelhay. 2021. "MTHFR C677T and A1298C Polymorphisms in Breast Cancer, Gliomas and Gastric Cancer: A Review" Genes 12, no. 4: 587. https://doi.org/10.3390/genes12040587
APA StylePetrone, I., Bernardo, P. S., dos Santos, E. C., & Abdelhay, E. (2021). MTHFR C677T and A1298C Polymorphisms in Breast Cancer, Gliomas and Gastric Cancer: A Review. Genes, 12(4), 587. https://doi.org/10.3390/genes12040587