
Microsatellite DNA Analysis for Diversity Study, Individual Identification and Parentage Control in Pig Breeds in Poland


Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.2. Methods
2.3. Data analysis
- -
- Polymorphic information content—PIC [24];
- -
- Power of discrimination—PD [25];
- -
- Probability of identity—PID [26];
- -
- Probability of parentage exclusion for each locus—when the genotypes of one and both parents are known (PE1 and PE2)—and the cumulative probability of parentage exclusion (CPE), according to Jamieson’s (1994) formula [27].
3. Results
3.1. Diversity Analysis
3.2. Parentage Testing and Individual Identification
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Poland in the European Union. A Statistical Portrait. Statistics Poland, Statistical Products Department. 2019. Available online: http:/stat.gov.pl (accessed on 2 October 2019).
- POLSUS. Pig Breeding. Available online: https://www.polsus.pl/index.php/en/pig-breeding (accessed on 1 January 2020).
- Rychlik, T.; Radko, A.; Duniec, M. Evaluating the usefulness of polymorphism of some genetic markers for parentage control of sheep. Med. Wet. 2003, 59, 1016–1018. [Google Scholar]
- Radko, A.; Rychlik, T.; Slota, E. Genetic characterization of the Wrzosowka sheep breed on the basis of 14 microsatellite DNA markers. Med. Wet. 2006, 62, 1073–1075. [Google Scholar]
- Radko, A. Microsatellite DNA polymorphism and its usefulness for pedigree verification of cattle raised in Poland. Ann. Anim. Sci. 2008, 4, 311–332. [Google Scholar]
- Salamon, D.; Gutierrez-Gil, B.; Arranz, J.J.; Barreta, J.; Batinic, V.; Dzidic, A. Genetic diversity and differentiation of 12 eastern Adriatic and western Dinaric native sheep breeds using microsatellites. Animal 2014, 8, 200–207. [Google Scholar] [CrossRef] [Green Version]
- Zabek, T.; Fornal, A. Evaluation of the 17-plex STR kit for parentage testing of Polish Cold blood and Hucul horses. Ann. Anim. Sci. 2009, 9, 363–372. [Google Scholar]
- Laval, G.; Iannuccelli, N.; Legault, C.; Milan, D.; Groenen, M.A.; Giuffra, E.; Andersson, L.; Nissen, P.H.; Jørgensen, C.B.; Beeckmann, P.; et al. Genetic diversity of eleven European pig breeds. Genet. Sel. Evol. 2000, 32, 187–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabuel, E.; Barragán, C.; Silió, L.; Rodríguez, M.C.; Toro, M.A. Analysis of genetic diversity and conservation priorities in Iberian pigs based on microsatellite markers. Heredity 2004, 93, 104–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicente, A.A.; Carolino, M.I.; Sousa, M.C.O.; Ginja, C.; Silva, F.S.; Martinez, A.M.; Vega-Pla, J.L.; Carolino, N.; Gama, L. Genetic diversity in native and commercial breeds of pigs in Portugal assessed by microsatellites. J. Anim. Sci. 2008, 86, 2496–2507. [Google Scholar] [CrossRef] [Green Version]
- Sollero, B.P.; Paiva, S.R.; Faria, D.A.; Guimarães, S.E.F.; Castro, S.T.R.; Egito, A.A.; Albuquerque, M.S.M.; Piovezan, U.; Bertani, G.R.; Mariante, A.D.S. Genetic diversity of Brazilian pig breeds evidenced by microsatellite markers. Livest. Sci. 2009, 123, 8–15. [Google Scholar] [CrossRef]
- Kramarenko, S.S.; Lugovoy, S.I.; Kharzinova, V.R.; Lykhach, V.Y.; Kramarenko, A.S.; Lykhach, A.V. Genetic diversity of Ukrainian local pig breeds based on microsatellite markers. Regul. Mech. Biosyst. 2018, 9, 177–182. [Google Scholar] [CrossRef]
- Margeta, P.; Škorput, D.; Šalomon, D.; Menčik, S.; Gvozdanović, K.; Karolyi, D.; Luković, Z.; Salajpal, K. 12-plex highly polymorphic microsatellite marker set for parentage analysis in Banija spotted pigs. JCEA 2019, 20, 50–54. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Hsieh, H.-M.; Lee, C.-I.J.; Hsiao, C.-T.; Lin, D.-Y.; Linacre, A.; Tsai, L.-C. Establishing a DNA identification system for pigs (Sus scrofa) using a multiplex STR amplification. For. Sci. Int. Genet. 2014, 9, 12–19. [Google Scholar] [CrossRef]
- ISAG Conference 2012, Cairns, Australia, Pig Molecular Comparison Test Report. Available online: https://www.isag.us/Docs/PigGenGenomics2012.pdf (accessed on 19 July 2014).
- ISAG Conference 2014, Xi’an, China, Pig Genetics and Genomics. Available online: https://www.isag.us/docs/Pig%20Genetics%20Genomics%20and%20CT_2014.pdf (accessed on 22 August 2014).
- Mucha, A.; Różycki, M. Breeding of the Polish Large White and Polish Landrace breeds from a historical perspective. Wiad. Zoot. 2012, 3, 9–18. [Google Scholar]
- Babicz, M.; Hałabis, M.; Skałecki, P.; Domaradzki, P.; Litwińczuk, A.; Kropiwiec-Domańska, K.; Łukasik, M. Breeding and performance potential of PULawska pigs. Ann. Anim. Sci. 2020, 20, 343–354. [Google Scholar] [CrossRef]
- Sieczkowska, H.; Iwan, R. Utilization of the Duroc breed in commercial production of fatteners. Przeg. Hod. 2011, 9, 26–29. [Google Scholar]
- Nei, M.; Roychoudhury, A.K. Sampling variances of heterozygosity and genetic distance. Genetics 1974, 76, 379–390. [Google Scholar] [CrossRef]
- Wright, S. Evolution and the Genetics of Populations; University of Chicago Press: Chicago, IL, USA, 1978; Volume 4. [Google Scholar]
- Guo, S.; Thompson, E.A. Performing the exact test of Hardy–Weinberg proportion for multiple alleles. Biometrics 1992, 48, 361–372. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. Royal Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Kimberly, A.H. Statistical analysis of STR data. Profiles DNA Promega Corp. 1998, 1, 14–15. [Google Scholar]
- Paetkau, D.; Strobeck, C. Microsatellite analysis of genetic variation in black bear population. Mol. Ecol. 1994, 3, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, A.; Taylor, S.C.S. Comparisons of three probability formulae for parentage exclusion. Anim. Genet. 1997, 28, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.D.; Song, K.D.; Seo, J.H.; Kim, D.K.; Kim, S.H.; Seo, K.S.; Lim, H.T.; Lee, J.B.; Park, H.C.; Ryu, Y.C.; et al. Genetic traceability of Black Pig Meats using microsatellite markers. Anim. Biosci. 2014, 27, 926–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charoensook, R.; Gatphayak, K.; Brenig, B.; Knorr, C. Genetic diversity analysis of Thai indigenous pig population using microsatellite markers. Asian-Australas J. Anim. Sci. 2019, 32, 1491–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, H.T.; Seo, B.Y.; Jung, E.J.; Yoo, C.K.; Zhong, T.; Cho, I.C.; Yoon, D.H.; Lee, J.G.; Jeon, J.T. Establishment of a microsatellite marker set for individual, pork brand and product origin identification in pigs. J. Anim. Sci. Technol. 2009, 51, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Li, T.; Zhu, C.; Jiang, X.; Zhao, Y.; Xu, Z.; Yang, S.; Chen, A. Selection and use of microsatellite markers for individual identification and meat traceability of six swine breeds in the Chinese market. Food. Sci. Technol. Int. 2018, 4, 292–300. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. Power Marker: Integrated analysis environment for genetic marker data. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [Green Version]
- Vrtková, I.; Filistowicz, A.; Buczyński, J.; Vrtek, Š.; Falková, L. Parameters of variability and diversity of tetrameric STRs for practical use in Zlotnicka White and Zlotnicka Spotted pigs. Anim. Sci. Pap. Rep. 2017, 3, 253–263. [Google Scholar]
- Nechtelberger, D.; Kaltwasser, C.; Stur, I.; Meyer, J.N.; Brem, G.; Mueller, M.; Mueller, S. DNA microsatellite analysis for parentage control in Austrian pigs. Anim. Biotechnol. 2001, 12, 141–144. [Google Scholar] [CrossRef]



{kind=link}
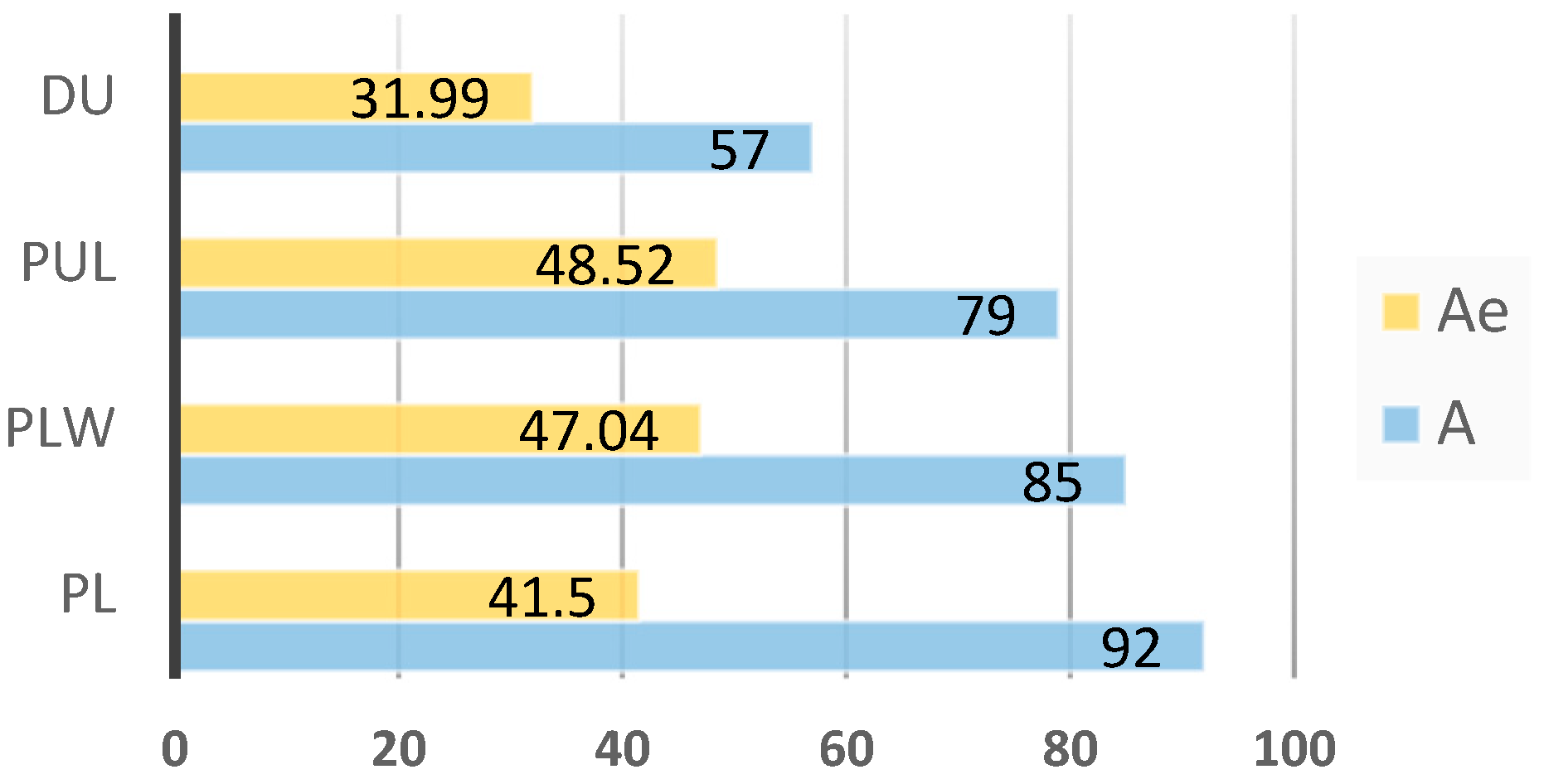{kind=link}
| locus | Size Range | Dye | Primer (5′-3′): Forward Primer (5′-3′): Reverse |
|---|---|---|---|
| S0090 | 244–251 | FAM | CAAGACTGCCTTGTAGGTGAATA GCTATCAAGTATTGTACCATTAGG |
| S0101 | 197–216 | FAM | GAATGCAAAGAGTTCAGTGTAGG GTCTCCCTCACACTTACCGCAG |
| S0155 | 150–166 | PET | TGTTCTCTGTTTCTCCTCTGTTTG AAAGTGGAAAGAGTCAATGGCTAT |
| S0227 | 231–256 | NED | GATCCATTTATAATTTTAGCACAAAGT GCATGGTGTGATGCTATGTCAAGC |
| S0228 | 222–249 | VIC | GGCATAGGCTGGCAGCAACA AGCCCACCTCATCTTATCTACACT |
| S0355 | 243–277 | PET | TCTGGCTCCTACACTCCTTCTTGATG TTGGGTGGGTGCTGAAAAATAGGA |
| S0386 | 156–174 | NED | GAACTCCTGGGTCTTATTTTCTA GTCAAAAATCTTTTTATCTCCAACAGTAT |
| SW24 | 96–121 | PET | CTTTGGGTGGAGTGTGTGC ATCCAAATGCTGCAAGCG |
| SW72 | 100–116 | VIC | ATCAGAACAGTGCGCCGT TTTGAAAATGGGGTGTTTCC |
| SW240 | 96–115 | FAM | AGAAATTAGTGCCTCAAATTGG AAACCATTAAGTCCCTAGCAAA |
| SW857 | 144–160 | VIC | TGAGAGGTCAGTTACAGAAGACC GATCCTCCTCCAAATCCCAT |
| SW911 | 153–177 | FAM | CTCAGTTCTTTGGGACTGAACC CATCTGTGGAAAAAAAAAGCC |
| SW936 | 80–117 | NED | TCTGGAGCTAGCATAAGTGCC GTGCAAGTACACATGCAGGG |
| SW951 | 125–133 | FAM | TTTCACAACTCTGGCACCAG GATCGTGCCCAAATGGAC |
| Locus | N | Breeds | Size Range | A | Ae |
|---|---|---|---|---|---|
| S0090 | 7 | PL | 234–254 | 7 | 2.90 |
| PLW | 244–254 | 5 | 4.05 | ||
| PUL | 244–254 | 5 | 2.81 | ||
| DU | 246–252 | 4 | 2.62 | ||
| S0101 | 6 | PL | 200–218 | 5 | 2.73 |
| PLW | 200–218 | 5 | 3.33 | ||
| PUL | 200–218 | 6 | 3.40 | ||
| DU | 212–218 | 4 | 1.49 | ||
| S0155 | 7 | PL | 152–168 | 6 | 2.86 |
| PLW | 152–166 | 5 | 2.56 | ||
| PUL | 152–166 | 5 | 2.88 | ||
| DU | 158–166 | 3 | 2.56 | ||
| S0227 | 3 | PL | 232–256 | 3 | 1.52 |
| PLW | 232–256 | 3 | 1.42 | ||
| PUL | 232–256 | 3 | 1.43 | ||
| DU | 232–256 | 2 | 1.54 | ||
| S0228 | 7 | PL | 218–244 | 7 | 2.65 |
| PLW | 218–244 | 6 | 5.21 | ||
| PUL | 218–244 | 6 | 2.61 | ||
| DU | 222–232 | 3 | 1.51 | ||
| S0355 | 7 | PL | 247–275 | 7 | 2.23 |
| PLW | 247–275 | 5 | 3.42 | ||
| PUL | 247–275 | 5 | 1.82 | ||
| DU | 247–275 | 4 | 1.25 | ||
| PL | 157–177 | 8 | 3.58 | ||
| S0386 | 8 | PLW | 157–177 | 7 | 2.94 |
| PUL | 157–177 | 6 | 3.02 | ||
| DU | 167–177 | 3 | 1.93 | ||
| SW24 | 9 | PL | 98–122 | 7 | 2.86 |
| PLW | 98–122 | 8 | 3.16 | ||
| PUL | 98–118 | 6 | 4.44 | ||
| DU | 98–122 | 7 | 3.94 | ||
| SW72 | 6 | PL | 103–119 | 6 | 2.86 |
| PLW | 103–119 | 6 | 4.00 | ||
| PUL | 103–119 | 6 | 3.11 | ||
| DU | 103–119 | 4 | 3.21 | ||
| SW240 | 9 | PL | 98–116 | 9 | 2.82 |
| PLW | 98–118 | 8 | 2.73 | ||
| PUL | 98–116 | 8 | 3.00 | ||
| DU | 100–116 | 5 | 2.65 | ||
| SW857 | 10 | PL | 138–160 | 10 | 6.76 |
| PLW | 138–158 | 9 | 5.03 | ||
| PUL | 138–158 | 9 | 6.25 | ||
| DU | 142–158 | 6 | 2.49 | ||
| SW911 | 7 | PL | 156–168 | 5 | 2.08 |
| PLW | 156–168 | 6 | 2.55 | ||
| PUL | 156–168 | 4 | 2.02 | ||
| DU | 154–168 | 4 | 2.12 | ||
| SW936 | 10 | PL | 94–114 | 9 | 3.97 |
| PLW | 92–114 | 8 | 4.13 | ||
| PUL | 94–114 | 7 | 3.52 | ||
| DU | 96–102 | 4 | 2.34 | ||
| SW951 | 5 | PL | 123–131 | 3 | 1.68 |
| PLW | 123–131 | 4 | 2.51 | ||
| PUL | 123–131 | 3 | 1.59 | ||
| DU | 123–131 | 4 | 2.34 |
| Locus | Breeds | HO | HE | FIS | HWE | PIC | PD | PID | PE1 | PE2 |
|---|---|---|---|---|---|---|---|---|---|---|
| S0090 | PL | 0.659 | 0.661 | 0.003 | 0.0002 *** | 0.617 | 0.839 | 0.158 | 0.253 | 0.427 |
| PLW | 0.737 | 0.753 | 0.022 | 0.0984 | 0.712 | 0.899 | 0.102 | 0.346 | 0.524 | |
| PUL | 0.626 | 0.644 | 0.028 | 0.0000 *** | 0.604 | 0.823 | 0.167 | 0.241 | 0.417 | |
| DU | 0.676 | 0.619 | −0.093 | 0.0000 *** | 0.542 | 0.712 | 0.222 | 0.201 | 0.340 | |
| S0101 | PL | 0.610 | 0.634 | 0.038 | 0.0000 *** | 0.585 | 0.818 | 0.183 | 0.227 | 0.393 |
| PLW | 0.622 | 0.690 | 0.091 | 0.1991 | 0.652 | 0.866 | 0.138 | 0.285 | 0.458 | |
| PUL | 0.682 | 0.706 | 0.033 | 0.0001 *** | 0.658 | 0.856 | 0.135 | 0.293 | 0.465 | |
| DU | 0.370 | 0.328 | −0.129 | 0.1360 | 0.298 | 0.533 | 0.482 | 0.055 | 0.165 | |
| S0155 | PL | 0.684 | 0.650 | −0.053 | 0.0196 | 0.583 | 0.803 | 0.189 | 0.227 | 0.379 |
| PLW | 0.544 | 0.610 | 0.108 | 0.0098 ** | 0.534 | 0.787 | 0.228 | 0.187 | 0.326 | |
| PUL | 0.668 | 0.653 | −0.023 | 1.0 | 0.591 | 0.808 | 0.182 | 0.231 | 0.388 | |
| DU | 0.630 | 0.610 | −0.032 | 1.0 | 0.535 | 0.763 | 0.227 | 0.190 | 0.330 | |
| S0227 | PL | 0.356 | 0.341 | −0.044 | 1.0 | 0.309 | 0.534 | 0.466 | 0.058 | 0.171 |
| PLW | 0.278 | 0.296 | 0.061 | 1.0 | 0.275 | 0.471 | 0.516 | 0.044 | 0.152 | |
| PUL | 0.275 | 0.290 | 0.080 | 1.0 | 0.282 | 0.464 | 0.508 | 0.046 | 0.159 | |
| DU | 0.352 | 0.350 | −0.006 | 1.0 | 0.295 | 0.514 | 0.478 | 0.061 | 0.151 | |
| S0228 | PL | 0.638 | 0.623 | −0.023 | 1.0 | 0.586 | 0.819 | 0.180 | 0.225 | 0.401 |
| PLW | 0.732 | 0.761 | 0.038 | 1.0 | 0.781 | 0.936 | 0.064 | 0.442 | 0.620 | |
| PUL | 0.607 | 0.617 | 0.017 | 1.0 | 0.569 | 0.808 | 0.194 | 0.215 | 0.380 | |
| DU | 0.324 | 0.337 | 0.038 | 1.0 | 0.291 | 0.507 | 0.485 | 0.057 | 0.153 | |
| S0355 | PL | 0.513 | 0.552 | 0.068 | 0.0044 ** | 0.495 | 0.746 | 0.257 | 0.164 | 0.310 |
| PLW | 0.720 | 0.708 | −0.017 | 0.1680 | 0.657 | 0.861 | 0.136 | 0.287 | 0.458 | |
| PUL | 0.327 | 0.452 | 0.276 | 0.0000 *** | 0.421 | 0.635 | 0.332 | 0.109 | 0.258 | |
| DU | 0.213 | 0.197 | −0.078 | 0.0970 | 0.191 | 0.364 | 0.651 | 0.020 | 0.105 | |
| S0386 | PL | 0.753 | 0.721 | −0.044 | 0.0032 ** | 0.688 | 0.884 | 0.111 | 0.326 | 0.510 |
| PLW | 0.479 | 0.660 | 0.274 | 0.0000 *** | 0.603 | 0.827 | 0.173 | 0.242 | 0.404 | |
| PUL | 0.645 | 0.669 | 0.037 | 0.0990 | 0.638 | 0.855 | 0.141 | 0.273 | 0.457 | |
| DU | 0.474 | 0.482 | 0.022 | 0.3570 | 0.437 | 0.675 | 0.314 | 0.118 | 0.260 | |
| SW24 | PL | 0.626 | 0.650 | 0.038 | 0.0046 ** | 0.598 | 0.834 | 0.174 | 0.241 | 0.406 |
| PLW | 0.612 | 0.684 | 0.105 | 0.0000 *** | 0.635 | 0.857 | 0.149 | 0.275 | 0.445 | |
| PUL | 0.730 | 0.775 | 0.058 | 0.0055 ** | 0.740 | 0.912 | 0.086 | 0.387 | 0.564 | |
| DU | 0.778 | 0.746 | −0.043 | 0.0000 *** | 0.715 | 0.892 | 0.096 | 0.358 | 0.541 | |
| SW72 | PL | 0.626 | 0.650 | 0.038 | 0.0074 ** | 0.598 | 0.834 | 0.174 | 0.241 | 0.406 |
| PLW | 0.751 | 0.750 | −0.002 | 1.0 | 0.710 | 0.895 | 0.102 | 0.348 | 0.526 | |
| PUL | 0.682 | 0.678 | −0.006 | 1.0 | 0.621 | 0.827 | 0.161 | 0.261 | 0.424 | |
| DU | 0.769 | 0.698 | −0.117 | 0.0012 ** | 0.624 | 0.809 | 0.162 | 0.251 | 0.410 | |
| SW240 | bz | 0.652 | 0.645 | −0.011 | 1.00 | 0.604 | 0.840 | 0.167 | 0.245 | 0.420 |
| PLW | 0.618 | 0.634 | 0.025 | 1.00 | 0.610 | 0.846 | 0.158 | 0.249 | 0.437 | |
| PUL | 0.635 | 0.667 | 0.048 | 0.0035 ** | 0.634 | 0.846 | 0.144 | 0.272 | 0.455 | |
| DU | 0.574 | 0.599 | 0.077 | 0.1820 | 0.565 | 0.789 | 0.200 | 0.209 | 0.368 | |
| SW857 | PL | 0.849 | 0.852 | 0.004 | 1.0 | 0.835 | 0.958 | 0.039 | 0.539 | 0.703 |
| PLW | 0.830 | 0.811 | −0.027 | 1.0 | 0.776 | 0.934 | 0.064 | 0.444 | 0.621 | |
| PUL | 0.844 | 0.840 | −0.005 | 1.0 | 0.821 | 0.950 | 0.044 | 0.518 | 0.686 | |
| DU | 0.593 | 0.599 | 0.011 | 0.1890 | 0.550 | 0.798 | 0.210 | 0.197 | 0.359 | |
| SW911 | PL | 0.537 | 0.520 | −0.033 | 1.0 | 0.434 | 0.680 | 0.317 | 0.137 | 0.246 |
| PLW | 0.591 | 0.608 | 0.027 | 1.0 | 0.533 | 0.773 | 0.228 | 0.195 | 0.334 | |
| PUL | 0.564 | 0.526 | −0.105 | 0.0978 | 0.436 | 0.668 | 0.314 | 0.131 | 0.254 | |
| DU | 0.593 | 0.528 | −0.122 | 0.1605 | 0.483 | 0.737 | 0.267 | 0.146 | 0.300 | |
| SW936 | PL | 0.738 | 0.748 | 0.013 | 0.0813 | 0.710 | 0.892 | 0.101 | 0.356 | 0.533 |
| PLW | 0.811 | 0.758 | −0.070 | 0.0616 | 0.720 | 0.893 | 0.096 | 0.361 | 0.539 | |
| PUL | 0.720 | 0.716 | −0.006 | 0.6825 | 0.672 | 0.879 | 0.125 | 0.313 | 0.487 | |
| DU | 0.657 | 0.572 | −0.150 | 0.4658 | 0.505 | 0.719 | 0.250 | 0.171 | 0.312 | |
| SW951 | PL | 0.418 | 0.406 | −0.030 | 1.0 | 0.366 | 0.610 | 0.393 | 0.082 | 0.208 |
| PLW | 0.527 | 0.592 | 0.121 | 0.0032 ** | 0.523 | 0.782 | 0.237 | 0.188 | 0.322 | |
| PUL | 0.398 | 0.381 | −0.062 | 0.1891 | 0.337 | 0.580 | 0.429 | 0.069 | 0.190 | |
| DU | 0.647 | 0.592 | −0.093 | 0.5454 | 0.505 | 0.719 | 0.250 | 0.171 | 0.312 | |
| 0.599 | 0.605 | 0.007 | 0.559 | 0.771 | ||||||
| Breeds | HO | HE | FIS | PIC | CPD | CPID | CPE1 | CPE2 |
|---|---|---|---|---|---|---|---|---|
| PL | 0.619 | 0.618 | −0.003 | 0.581 | 1 * | 1.8 × 10−11 | 0.98413 | 0.99952 |
| PLW | 0.632 | 0.665 | 0.054 | 0.623 | 1 * | 2.0 × 10−12 | 0.99096 | 0.99978 |
| PUL | 0.600 | 0.616 | 0.026 | 0.573 | 1 * | 2.8 × 10−11 | 0.98235 | 0.99946 |
| DU | 0.546 | 0.520 | −0.051 | 0.477 | 1 * | 6.9 × 10−9 | 0.92755 | 0.99492 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radko, A.; Smołucha, G.; Koseniuk, A. Microsatellite DNA Analysis for Diversity Study, Individual Identification and Parentage Control in Pig Breeds in Poland. Genes 2021, 12, 595. https://doi.org/10.3390/genes12040595
Radko A, Smołucha G, Koseniuk A. Microsatellite DNA Analysis for Diversity Study, Individual Identification and Parentage Control in Pig Breeds in Poland. Genes. 2021; 12(4):595. https://doi.org/10.3390/genes12040595
Chicago/Turabian StyleRadko, Anna, Grzegorz Smołucha, and Anna Koseniuk. 2021. "Microsatellite DNA Analysis for Diversity Study, Individual Identification and Parentage Control in Pig Breeds in Poland" Genes 12, no. 4: 595. https://doi.org/10.3390/genes12040595
APA StyleRadko, A., Smołucha, G., & Koseniuk, A. (2021). Microsatellite DNA Analysis for Diversity Study, Individual Identification and Parentage Control in Pig Breeds in Poland. Genes, 12(4), 595. https://doi.org/10.3390/genes12040595