
Host-Diet Effect on the Metabolism of Bifidobacterium

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genomic Data and Annotations
2.2. Orthologous Gene Clustering
2.3. Identification of Carbohydrate-Active Enzymes
2.4. Selection of the GH Families for Clustering Bifidobacterium Strains
2.5. Phylogenetic Analysis
2.6. Statistical Analysis
3. Results and Discussion
3.1. Host Diet and the Genome Size of Type Strains
3.2. Distribution of Carbohydrate-Active Enzymes
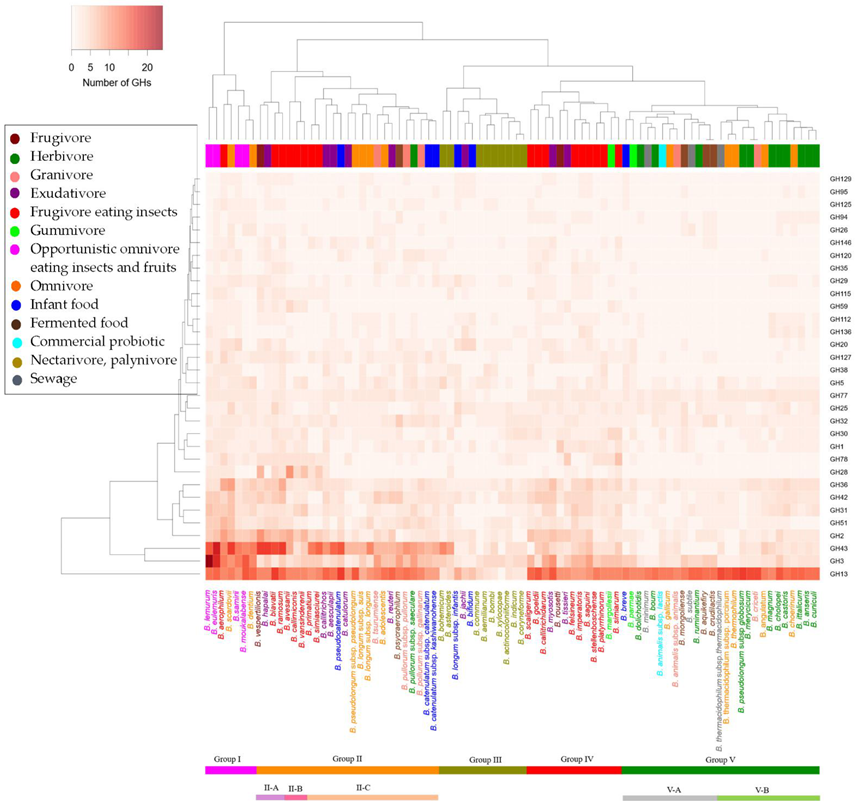
3.3. Clustering of Bifidobacterium Species Based on GH Families
- Group I included strains with the largest number of GH genes. This group reflected species from opportunistic omnivore eating insects and fruits. The group had high numbers of GH43 and GH3 genes associated with degradation of complex plant polysaccharides like xylan, arabinan or arabinoxylan. This suggested that these GH genes were adapted to the hosts of mixed diets (omnivore and frugivore).
- Group II included strains with a high number of GH43 but low GH3. The group included 25 species and was further divided into three: subgroup II-A, -B and -C. The subgroup II-C possessed low numbers of GH2, GH28, GH59 and GH115. The dietary pattern of the hosts varied: omnivore, herbivore, frugivore, insectivore and exudativore.
- Group III included bee isolates and two infant isolates. This group possessed a very low number of GH13. This result was supported by previous studies where the GHs from the insects clustered separately [26]. GH13 enzymes are involved in degradation of starches and malto-oligosaccharides, and such sugars are usually scarce in diets of bees and infants.
- Group IV included strains from hosts of insect and fruit diet. This group had the second highest gene counts for GHs after Group I, which suggested that the species from frugivorous hosts possessed more GH genes.
- Group V included the largest number of strains. This group had the lowest GH gene counts, where many of the GH families were mostly absent (e.g., no GH28, GH38 and GH115). The group was further divided into two subgroups (subgroup V-A and V-B). Subgroup V-B was strains from herbivorous hosts while subgroup V-A included strains from hosts of mixed dietary habits.
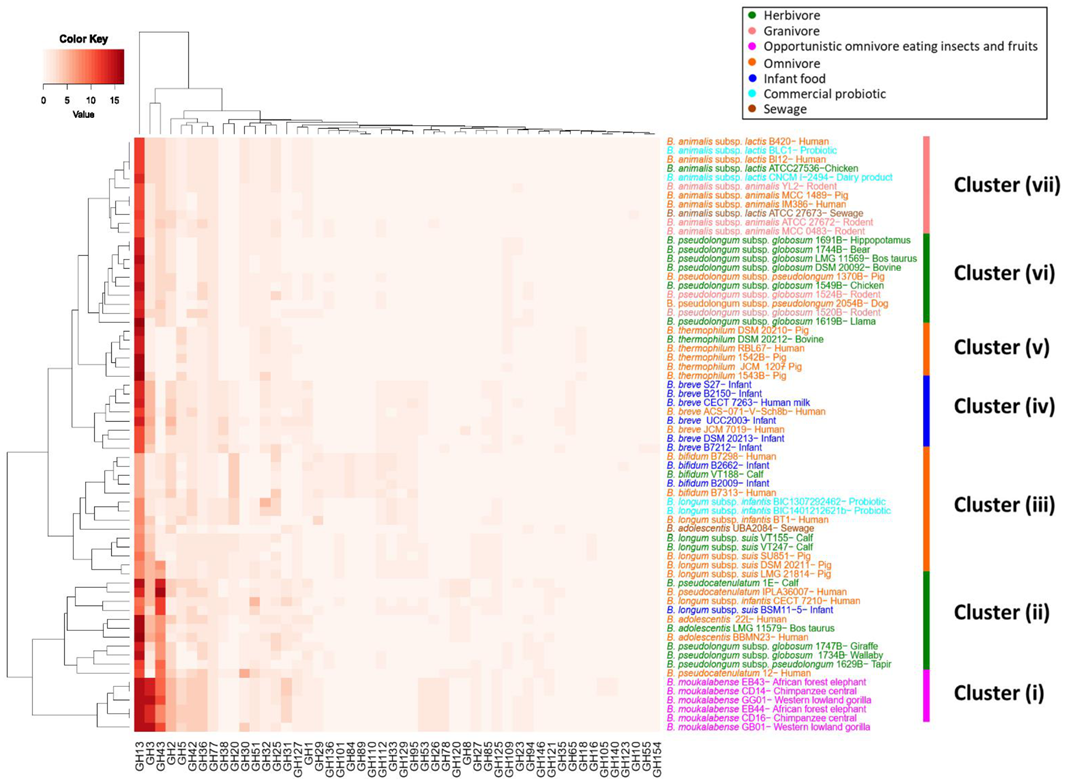
3.4. Comparison of Bifidobacterium Species from Multiple Host Animals
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alberoni, D.; Gaggìa, F.; Baffoni, L.; Modesto, M.M.; Biavati, B.; Di Gioia, D. Bifidobacterium xylocopae sp. nov. and Bifidobacterium aemilianum sp. nov., from the carpenter bee (Xylocopa violacea) digestive tract. Syst. Appl. Microbiol. 2019, 42, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Modesto, M.; Watanabe, K.; Arita, M.; Satti, M.; Oki, K.; Sciavilla, P.; Patavino, C.; Cammà, C.; Michelini, S.; Sgorbati, B.; et al. Bifidobacterium jacchi sp. nov., isolated from the faeces of a baby common marmoset (Callithrix jacchus). Int. J. Syst. Evol. Microbiol. 2019, 69, 2477–2485. [Google Scholar] [CrossRef] [PubMed]
- Modesto, M.; Satti, M.; Watanabe, K.; Puglisi, E.; Morelli, L.; Huang, C.-H.; Liou, J.-S.; Miyashita, M.; Tamura, T.; Saito, S.; et al. Characterization of Bifidobacterium species in feaces of the Egyptian fruit bat: Description of B. vespertilionis sp. nov. and B. rousetti sp. nov. Syst. Appl. Microbiol. 2019, 42, 126017. [Google Scholar] [CrossRef] [PubMed]
- Duranti, S.; Lugli, G.A.; Napoli, S.; Anzalone, R.; Milani, C.; Mancabelli, L.; Alessandri, G.; Turroni, F.; Ossiprandi, M.C.; Van Sinderen, D.; et al. Characterization of the phylogenetic diversity of five novel species belonging to the genus Bifidobacterium: Bifidobacterium castoris sp. nov., Bifidobacterium callimiconis sp. nov., Bifidobacterium goeldii sp. nov., Bifidobacterium samirii sp. nov. and Bifidobacterium dolichotidis sp. nov. Int. J. Syst. Evol. Microbiol. 2019, 69, 1288–1298. [Google Scholar] [CrossRef]
- Lugli, G.A.; Mangifesta, M.; Duranti, S.; Anzalone, R.; Milani, C.; Mancabelli, L.; Alessandri, G.; Turroni, F.; Ossiprandi, M.C.; van Sinderen, D.; et al. Phylogenetic classification of six novel species belonging to the genus Bifidobacterium comprising Bifidobacterium anseris sp. nov., Bifidobacterium criceti sp. nov., Bifidobacterium imperatoris sp. nov., Bifidobacterium italicum sp. nov., Bifidobacterium margollesii sp. nov. and Bifidobacterium parmae sp. nov. Syst. Appl. Microbiol. 2018, 41, 173–183. [Google Scholar] [CrossRef]
- Trovatelli, L.D.; Crociani, F.; Pedinotti, M.; Scardovi, V. Bifidobacterium pullorum sp. nov.: A new species isolated from chicken feces and a related group of bifidobacteria isolated from rabbit feces. Arch. Microbiol. 1974, 98, 187–198. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, W.; Guo, C.; Yang, X.; Liu, W.; Wu, Y.; Song, Y.; Kwok, L.Y.; Cui, Y.; Menghe, B.; et al. Comparative Genomic Analysis of 45 Type Strains of the Genus Bifidobacterium: A Snapshot of Its Genetic Diversity and Evolution. PLoS ONE 2015, 10, e0117912. [Google Scholar] [CrossRef] [Green Version]
- Milani, C.; Lugli, G.A.; Duranti, S.; Turroni, F.; Bottacini, F.; Mangifesta, M.; Sanchez, B.; Viappiani, A.; Mancabelli, L.; Taminiau, B.; et al. Genomic Encyclopedia of Type Strains of the Genus Bifidobacterium. Appl. Environ. Microbiol. 2014, 80, 6290–6302. [Google Scholar] [CrossRef] [Green Version]
- Bottacini, F.; Medini, D.; Pavesi, A.; Turroni, F.; Foroni, E.; Riley, D.; Giubellini, V.; Tettelin, H.; Van Sinderen, D.; Ventura, M. Comparative genomics of the genus Bifidobacterium. Microbiology 2010, 156, 3243–3254. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, C.I.; Martiny, J.B.H. Evolutionary relationships among bifidobacteria and their hosts and environments. BMC Genom. 2020, 21, 26. [Google Scholar] [CrossRef] [Green Version]
- Bottacini, F.; Milani, C.; Turroni, F.; Sánchez, B.; Foroni, E.; Duranti, S.; Serafini, F.; Viappiani, A.; Strati, F.; Ferrarini, A.; et al. Bifidobacterium asteroides PRL2011 Genome Analysis Reveals Clues for Colonization of the Insect Gut. PLoS ONE 2012, 7, e44229. [Google Scholar] [CrossRef] [Green Version]
- Tanizawa, Y.; Fujisawa, T.; Nakamura, Y. DFAST: A flexible prokaryotic genome annotation pipeline for faster genome publication. Bioinformatics 2018, 34, 1037–1039. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Moreira, B.; Vinuesa, P. GET_HOMOLOGUES, a Versatile Software Package for Scalable and Robust Microbial Pangenome Analysis. Appl. Environ. Microbiol. 2013, 79, 7696–7701. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Stoeckert, C.J.; Roos, D.S. OrthoMCL: Identification of Ortholog Groups for Eukaryotic Genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [Green Version]
- Satti, M.; Tanizawa, Y.; Endo, A.; Arita, M. Comparative analysis of probiotic bacteria based on a new definition of core genome. J. Bioinform. Comput. Biol. 2018, 16. [Google Scholar] [CrossRef]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. dbCAN2: A meta server for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Keck, F.; Rimet, F.; Bouchez, A.; François, K. Phylosignal: An R package to measure, test, and explore the phylogenetic signal. Ecol. Evol. 2016, 6, 2774–2780. [Google Scholar] [CrossRef]
- Blomberg, S.P.; Garland, T., Jr.; Ives, A.R. Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Evolution 2003, 57, 717–745. [Google Scholar] [CrossRef]
- Lugli, G.A.; Milani, C.; Duranti, S.; Alessandri, G.; Turroni, F.; Mancabelli, L.; Tatoni, D.; Ossiprandi, M.C.; Van Sinderen, D.; Ventura, M. Isolation of novel gut bifidobacteria using a combination of metagenomic and cultivation approaches. Genome Biol. 2019, 20, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Stevens, J.R.; Hallinan, E.V.; Hauser, M.D. The ecology and evolution of patience in two New World monkeys. Biol. Lett. 2005, 1, 223–226. [Google Scholar] [CrossRef]
- Kwiecinski, G.G.; Griffiths, T.A. Rousettus egyptiacus. Mamm. Species 1999, 611, 1–9. [Google Scholar] [CrossRef]
- Milani, C.; Lugli, G.A.; Duranti, S.; Turroni, F.; Mancabelli, L.; Ferrario, C.; Mangifesta, M.; Hevia, A.; Viappiani, A.; Scholz, M.; et al. Bifidobacteria exhibit social behavior through carbohydrate resource sharing in the gut. Sci. Rep. 2015, 5, 15782. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and their role as members of the human gut microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef] [Green Version]
- Ward, R.E.; Niñonuevo, M.; Mills, D.A.; Lebrilla, C.B.; German, J.B. In vitro fermentability of human milk oligosaccharides by several strains of bifidobacteria. Mol. Nutr. Food Res. 2007, 51, 1398–1405. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dietary Groups | GH Families | Related Activities in Bifidobacteria [27] |
|---|---|---|
| Opportunistic omnivore eating insects and fruits and Frugivore eating insects (Group I, Group II-B and Group IV) | GH13 | α-1,4-glucosidase, amylopullulanase, sucrose Phosphorylase, α-amylase |
| GH3 | β-glucosidase, β-hexosaminidase | |
| GH43 | Endo-1,5-α-l-arabinosidase, α-l-arabinofuranosidase, Endo-1,4-β-xylanase, β-1,4-xylosidase | |
| GH26 | Endo-1,4-β-mannosidase | |
| GH53 | Endogalactanase | |
| GH31 | α-xylosidase | |
| GH78 | α-l-rhamnosidase | |
| CBM67 | l-rhamnose binding activity | |
| Frugivore eating insects (Group II-B and Group IV) | GH115 | xylan α-1,2-glucuronidase, α-(4-O-methyl)-glucuronidase |
| GH28 | Galacturan1,4-α-galacturonidase, pectinesterase | |
| Herbivore (Group V-B) | GH94 | Cellobiose-phosphorylase |
| GH36 | α-galactosidase, raffinose synthase | |
| Infant food (Group II-C) | GH33 | Sialidase |
| GH20 | β-hexosaminidase | |
| GH29 | α-l-fucosidase | |
| GH95 | α-l-fucosidase | |
| GH112 | Lacto-N-biosephosphorylase | |
| GH29 | α-l-fucosidase | |
| GH95 | α-l-fucosidase | |
| Nectarivore and Palynivore (Group III) | GH65 | α,α-trehalase |
| GH13 * | α-1,4-glucosidase, amylopullulanase, sucrose Phosphorylase, α-amylase | |
| GT20 | α,α-trehalose-phosphate synthase | |
| GT35 * | glycogen or starch phosphorylase | |
| CBM48 * | appended to GH13 modules | |
| CE10 * | arylesterase |
| Family | Related Subfamilies | Significantly High | Significantly Low |
|---|---|---|---|
| GH1 | β-glucosidase, β-galactosidase | B. bifidum | B. longum subsp. suis |
| GH2 | β-galactosidase | all others | B. thermophilum |
| GH3 | β-glucosidase, β-hexosaminidase, β-glucosideglucohydrolase | B. thermophilum, B. bifidum | B. moukalabense |
| GH5 | β-mannosidase, β-glucosidase, β-exoglucanase | B. moukalabense | B. pseudolongum subsp. globosum |
| GH13 | α-1,4-glucosidase, amylopullulanase, sucrose phosphorylase, α-amylase | B. moukalabense | B. bifidum |
| GH20 | β-hexosaminidase | B. bifidum | all others |
| GH26 | Endo-1,4-β-mannosidase | B. moukalabense | all others |
| GH27 | α-galactosidase | B. moukalabense | all others |
| GH29 | α-L-fucosidase | B. bifidum | B. thermophilum |
| GH30 | β-d-xylosidase, endo-1,6-β-glucosidase, Glucosylceramidase | all others | B. thermophilum |
| GH31 | α-xylosidase | B. moukalabense | all others |
| GH32 | β-fructofuranosidase, sucrose-6-phosphatehydrolase | all others | B. bifidum |
| GH33 | Sialidase | B. bifidum | B. pseudolongum subsp. globosum |
| GH36 | α-galactosidase, raffinosesynthase | B. moukalabense | B. thermophilum |
| GH43 | Endo-1,5-α-l-arabinosidase, α-l-arabinofuranosidase, Endo-1,4-β-xylanase, β-1,4-xylosidase | all others | B. thermophilum |
| GH51 | α-L-arabinofuranosidase | B. moukalabense | B. bifidum |
| GH53 | Endogalactanase | B. moukalabense | B. pseudolongum subsp. globosum |
| GH77 | 4-α-glucanotransferase | B. bifidum | all others |
| GH78 | α-l-rhamnosidase | B. moukalabense | all others |
| GH84 | α-l-rhamnosidase | B. bifidum | all others |
| GH85 | Endo-β-N-acetylglucosaminidase D | B. longum subsp. suis | all others |
| GH89 | α-N-acetylglucosaminidase, β-N-hexosaminidase | B. bifidum | all others |
| GH94 | Cellobiose-phosphorylase | B. moukalabense | all others |
| GH95 | α-l-fucosidase | B. bifidum | all others |
| GH101 | endo-α-N-acetylgalactosaminidase | B. bifidum | all others |
| GH109 | α-N-acetylgalactosaminidase | B. pseudolongum subsp. globosum | all others |
| GH110 | Exo-α-galactosidase | B. bifidum | all others |
| GH112 | Lacto-N-biosephosphorylase | B. bifidum | all others |
| GH120 | β-xylosidase | B. pseudocatenulatum | all others |
| GH121 | β-galactosidase | B. pseudocatenulatum | all others |
| GH127 | β-l-arabinofuranosidase | B. moukalabense | all others |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satti, M.; Modesto, M.; Endo, A.; Kawashima, T.; Mattarelli, P.; Arita, M. Host-Diet Effect on the Metabolism of Bifidobacterium. Genes 2021, 12, 609. https://doi.org/10.3390/genes12040609
Satti M, Modesto M, Endo A, Kawashima T, Mattarelli P, Arita M. Host-Diet Effect on the Metabolism of Bifidobacterium. Genes. 2021; 12(4):609. https://doi.org/10.3390/genes12040609
Chicago/Turabian StyleSatti, Maria, Monica Modesto, Akihito Endo, Takeshi Kawashima, Paola Mattarelli, and Masanori Arita. 2021. "Host-Diet Effect on the Metabolism of Bifidobacterium" Genes 12, no. 4: 609. https://doi.org/10.3390/genes12040609
APA StyleSatti, M., Modesto, M., Endo, A., Kawashima, T., Mattarelli, P., & Arita, M. (2021). Host-Diet Effect on the Metabolism of Bifidobacterium. Genes, 12(4), 609. https://doi.org/10.3390/genes12040609