
Chronic Mild Stress and Venlafaxine Treatment Were Associated with Altered Expression Level and Methylation Status of New Candidate Inflammatory Genes in PBMCs and Brain Structures of Wistar Rats

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Chronic Mild Stress Procedure
2.3. Specimen Collection
2.4. RNA and DNA Isolation from Peripheral Blood Mononuclear Cells
2.5. Specimen Collection; RNA and DNA Isolation from Brain Tissues
2.6. Reverse Transcription and Gene Expression
2.7. Methylation and HRM Analysis
2.8. Drugs
2.9. Statistical Analysis
3. Results
3.1. Sucrose Intakes and Body Weights of Animals Exposed to CMS and Venlafaxine Administration
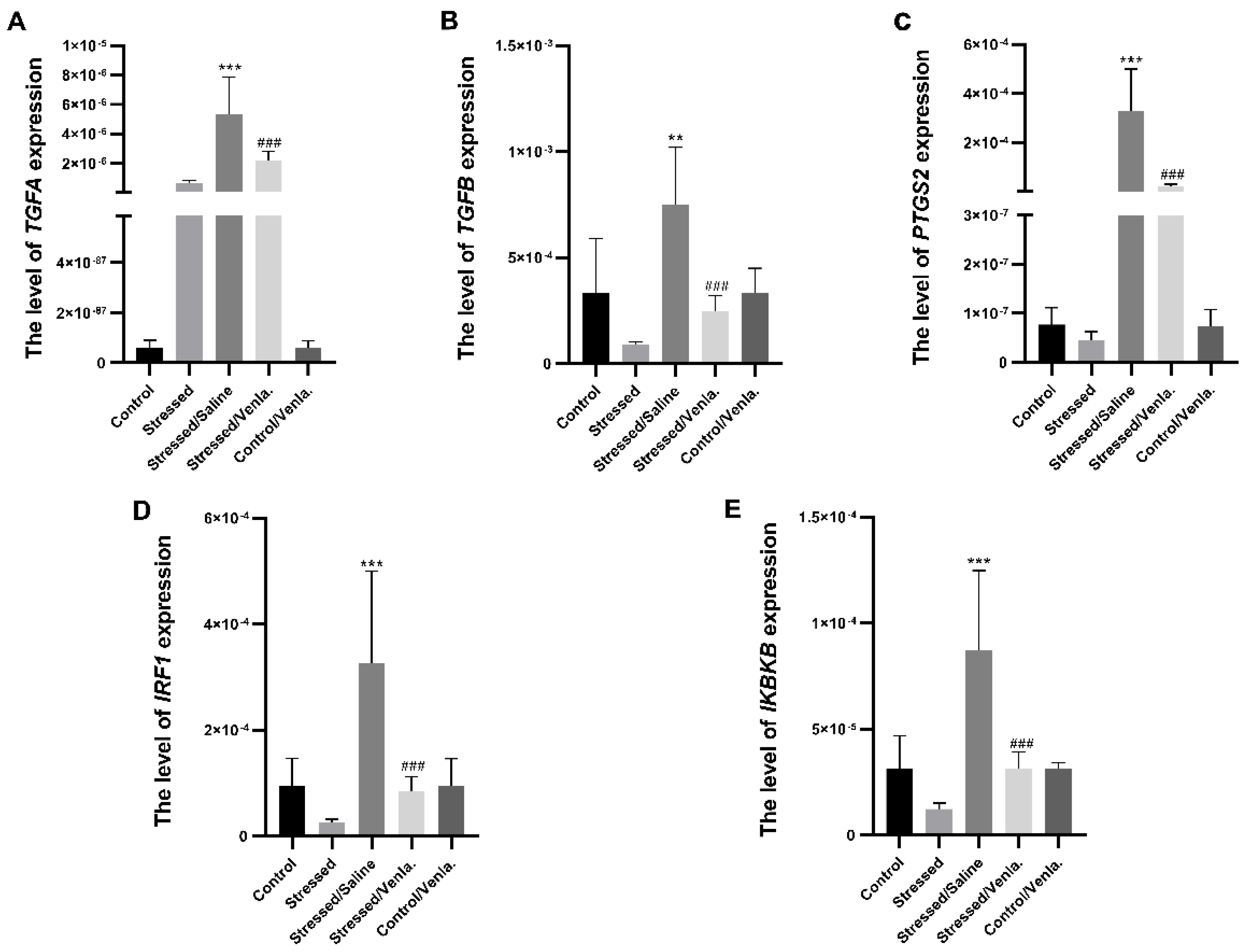
3.2. Gene Expression
3.2.1. Gene Expression in PBMCs after CMS Procedure and Venlafaxine Administration
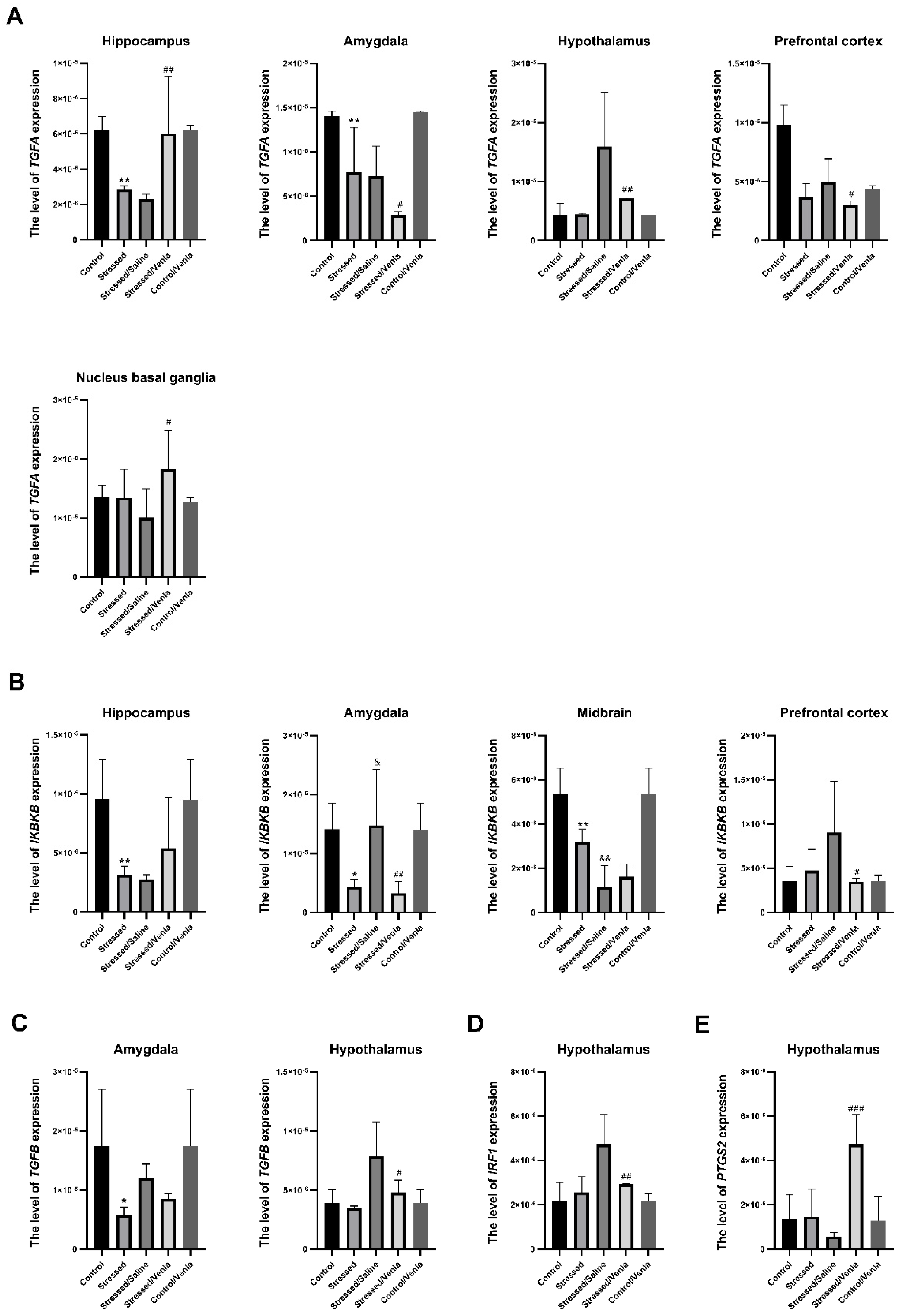
3.2.2. Gene Expression in Brain Structures after CMS Procedure and Venlafaxine Administration
3.3. Methylation of Studied Genes Promoters
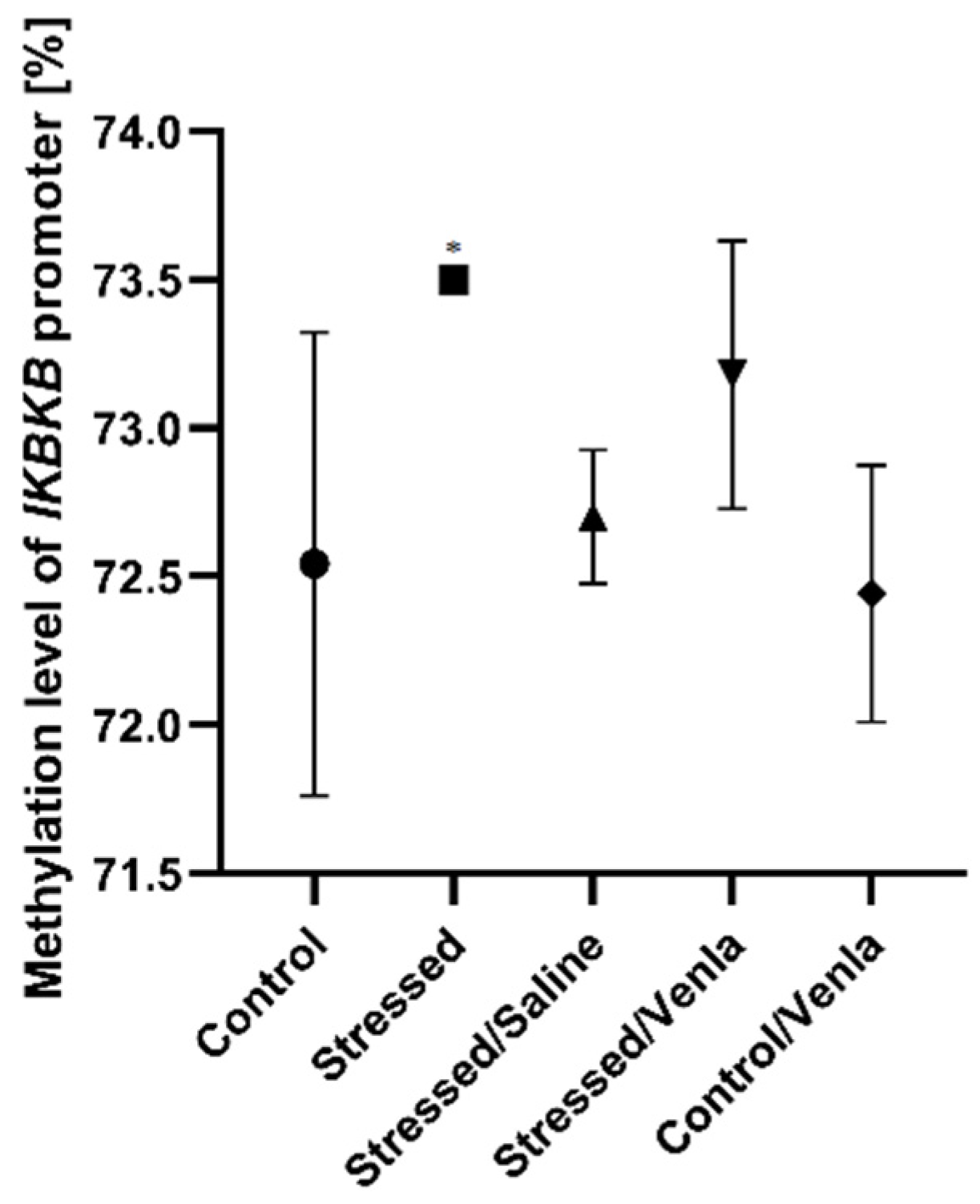
3.3.1. Methylation Status in PBMCs after CMS Procedure and Venlafaxine Administration
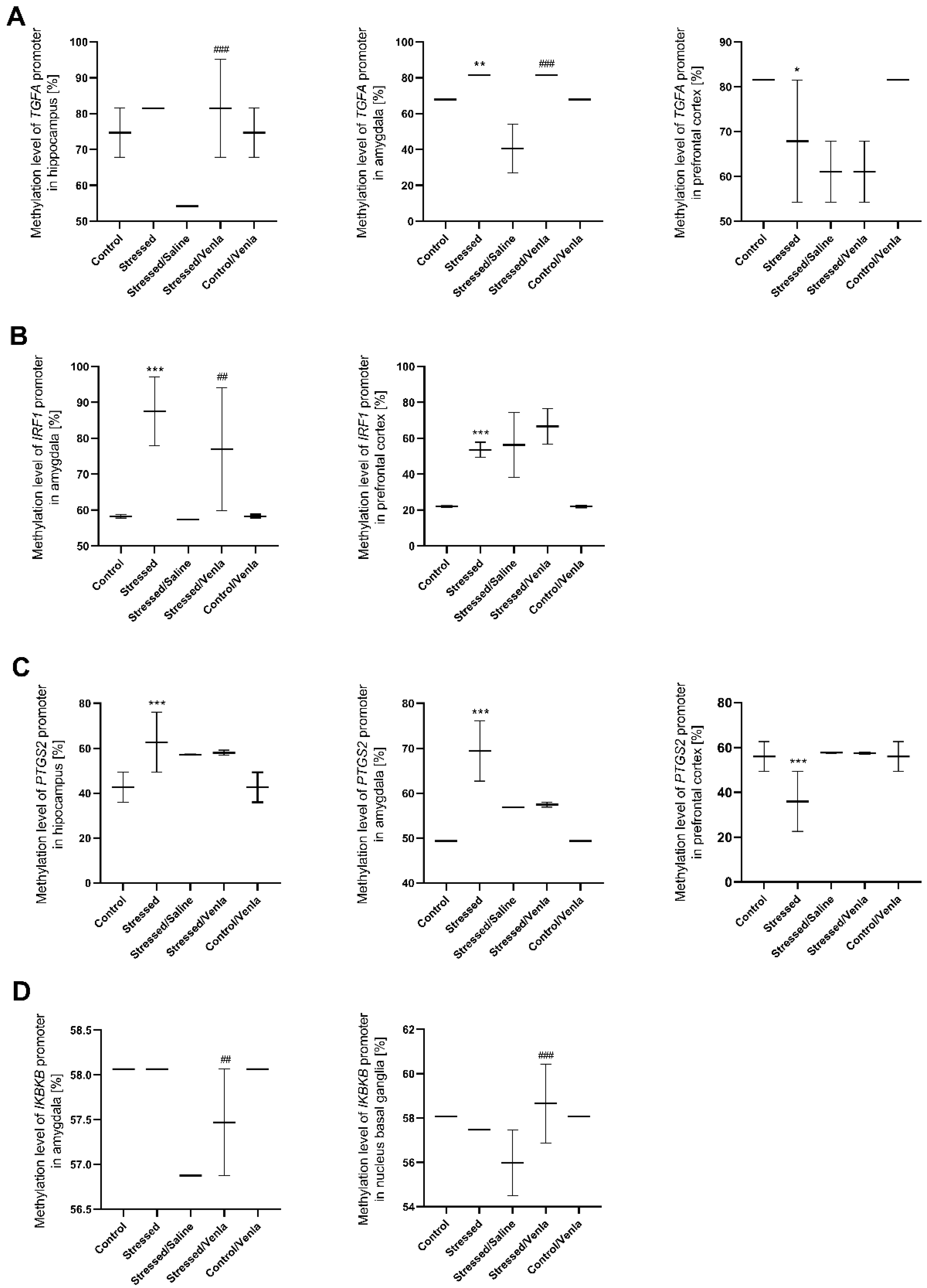
3.3.2. Methylation Status in Brain after CMS Procedure and Venlafaxine Administration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Depression. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/d (accessed on 1 June 2020).
- Al-Harbi, K.S. Treatment-resistant depression: Therapeutic trends, challenges, and future directions. Patient Prefer. Adherence 2012, 6, 369–388. [Google Scholar] [CrossRef] [Green Version]
- Ionescu, D.F.; Rosenbaum, J.F.; Alpert, J.E. Pharmacological approaches to the challenge of treatment-resistant depression. Dialog. Clin. Neurosci. 2015, 17, 111–126. [Google Scholar]
- Capuron, L.; Miller, A.H. Immune system to brain signaling: Neuropsychopharmacological implications. Pharmacol. Ther. 2011, 130, 226–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiepers, O.J.; Wichers, M.C.; Maes, M. Cytokines and major depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2005, 29, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.L.; Charney, D.S.; Lewis, L.; Golden, R.N.; Gorman, J.M.; Krishnan, K.R.R.; Nemeroff, C.B.; Bremner, J.D.; Carney, R.M.; Coyne, J.C.; et al. Mood Disorders in the Medically Ill: Scientific Review and Recommendations. Biol. Psychiatry 2005, 58, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Zorrilla, E.P.; Luborsky, L.; McKay, J.R.; Rosenthal, R.; Houldin, A.; Tax, A.; McCorkle, R.; Seligman, D.A.; Schmidt, K. The relationship of depression and stressors to im-munological assays: A meta-analytic review. Brain Behav. Immun. 2001, 15, 199–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howren, M.B.; Lamkin, D.M.; Suls, J. Associations of Depression with C-Reactive Protein, IL-1, and IL-6: A Meta-Analysis. Psychosom. Med. 2009, 71, 171–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, A.H.; Maletic, V.; Raison, C.L. Inflammation and Its Discontents: The Role of Cytokines in the Pathophysiology of Major Depression. Biol. Psychiatry 2009, 65, 732–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctôt, K.L. A Meta-Analysis of Cytokines in Major Depression. Biol. Psychiatry 2010, 67, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Steiner, J.; Bielau, H.; Brisch, R.; Danos, P.; Ullrich, O.; Mawrin, C.; Bernstein, H.G.; Bogerts, B. Immunological aspects in the neurobiology of suicide: Ele-vated microglial density in schizophrenia and depression is associated with suicide. J. Psychiatr. Res. 2008, 42, 151–157. [Google Scholar] [CrossRef]
- Michelucci, A.; Heurtaux, T.; Grandbarbe, L.; Morga, E.; Heuschling, P. Characterization of the microglial phenotype under specific pro-inflammatory and anti-inflammatory conditions: Effects of oligomeric and fibrillar amyloid-β. J. Neuroimmunol. 2009, 210, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Min, J.S.; Kim, B.; Chae, U.B.; Yun, J.W.; Choi, M.S.; Kong, I.K.; Chang, K.T.; Lee, D.S. Mitochondrial ROS govern the LPS-induced pro-inflammatory response in microglia cells by regulating MAPK and NF-κB pathways. Neurosci. Lett. 2015, 584, 191–196. [Google Scholar] [CrossRef]
- Kissin, E.Y.; Lemaire, R.; Korn, J.H.; Lafyatis, R. Transforming growth factor β induces fibroblast fibrillin-1 matrix formation. Arthritis Rheum. 2002, 46, 3000–3009. [Google Scholar] [CrossRef] [PubMed]
- Yamagiwa, S.; Gray, J.D.; Hashimoto, S.; Horwitz, D.A. A Role for TGF-β in the Generation and Expansion of CD4 + CD25 + Reg-ulatory T Cells from Human Peripheral Blood. J. Immunol. 2001, 166, 7282–7289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, J.-S.; Terabe, M.; Kang, M.-J.; Chae, H.; Voong, N.; Yang, Y.-A.; Laurence, A.; Michalowska, A.M.; Mamura, M.; Lonning, S.; et al. Transforming Growth Factor β Subverts the Immune System into Directly Promoting Tumor Growth through Interleukin-17. Cancer Res. 2008, 68, 3915–3923. [Google Scholar] [CrossRef] [Green Version]
- Passos, S.T.; Silver, J.S.; O’Hara, A.C.; Sehy, D.; Stumhofer, J.S.; Hunter, C.A. IL-6 Promotes NK Cell Production of IL-17 during Toxo-plasmosis. J. Immunol. 2010, 184, 1776–1783. [Google Scholar] [CrossRef] [Green Version]
- Vivien, D.; Ali, C. Transforming growth factor-β signalling in brain disorders. Cytokine Growth Factor Rev. 2006, 17, 121–128. [Google Scholar] [CrossRef]
- Hong, M.; Zheng, J.; Ding, Z.-Y.; Chen, J.-H.; Yu, L.; Niu, Y.; Hua, Y.-Q.; Wang, L.-L. Imbalance between Th17 and Treg Cells May Play an Important Role in the Development of Chronic Unpredictable Mild Stress-Induced Depression in Mice. Neuroimmunomodulation 2013, 20, 39–50. [Google Scholar] [CrossRef]
- Musil, R.; Schwarz, M.; Riedel, M.; Dehning, S.; Cerovecki, A.; Spellmann, I.; Arolt, V.; Müller, N. Elevated macrophage migration inhibitory factor and decreased transforming growth factor-beta levels in major depression—No influence of celecoxib treatment. J. Affect. Disord. 2011, 134, 217–225. [Google Scholar] [CrossRef]
- Sutcigil, L.; Oktenli, C.; Musabak, U.; Bozkurt, A.; Cansever, A.; Uzun, O.; Sanisoglu, S.Y.; Yesilova, Z.; Ozmenler, N.; Ozsahin, A.; et al. Pro- and anti-inflammatory cytokine balance in ma-jor depression: Effect of sertraline therapy. Clin. Dev. Immunol. 2007, 2007, 76396. [Google Scholar] [CrossRef] [Green Version]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 25, 639–653. [Google Scholar] [CrossRef]
- Hansson, M.; Olsson, I.; Nauseef, W.M. Biosynthesis, processing, and sorting of human myeloperoxidase. Arch. Biochem. Biophys. 2006, 445, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Minghetti, L. Cyclooxygenase-2 (COX-2) in Inflammatory and Degenerative Brain Diseases. J. Neuropathol. Exp. Neurol. 2004, 63, 901–910. [Google Scholar] [CrossRef] [Green Version]
- Minghetti, L. Role of COX-2 in Inflammatory and Degenerative Brain Diseases. Subcell. Biochem. 2007, 42, 127–141. [Google Scholar] [CrossRef]
- Cassano, P.; Hidalgo, A.; Burgos, V.; Adris, S.; Argibay, P. Hippocampal upregulation of the cyclooxygenase-2 gene following neonatal clomipramine treatment (a model of depression). Pharm. J. 2006, 6, 381–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kröger, A.; Köster, M.; Schroeder, K.; Hauser, H.; Mueller, P.P. Review: Activities of IRF-1. J. Interf. Cytokine Res. 2002, 22, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Yanai, H.; Savitsky, D.; Taniguchi, T. The IRF Family Transcription Factors in Immunity and Oncogenesis. Annu. Rev. Immunol. 2008, 26, 535–584. [Google Scholar] [CrossRef] [PubMed]
- Zahiu, C.D.M.; Rimbas, M. Neuropsychiatric side-effects of interferon-alpha treatment: Pathophysiology and therapeutic op-tions. In Maedica; 2014; 9, pp. 121–126. Available online: https://pubmed.ncbi.nlm.nih.gov/25705266 (accessed on 1 June 2020).
- Bierhaus, A.; Wolf, J.; Andrassy, M.; Rohleder, N.; Humpert, P.M.; Petrov, D.; Ferstl, R.; Von Eynatten, M.; Wendt, T.; Rudofsky, G.; et al. A mechanism converting psychosocial stress into mononuclear cell activation. Proc. Natl. Acad. Sci. USA 2003, 100, 1920–1925. [Google Scholar] [CrossRef] [Green Version]
- Pace, T.W.; Mletzko, T.C.; Alagbe, O.; Musselman, D.L.; Nemeroff, C.B.; Miller, A.H.; Heim, C.M. Increased stress-induced inflammatory responses in male patients with major depression and increased early life stress. Am. J. Psychiatry 2006, 163, 1630–1633. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Krakauer, T. Nuclear factor-κB: Fine-tuning a central integrator of diverse biologic stimuli. Int. Rev. Immunol. 2008, 27, 286–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-κB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerondakis, S.; Fulford, T.S.; Messina, N.L.; Grumont, R.J. NF-κB control of T cell development. Nat. Immunol. 2014, 15, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Shen, S.; Verma, I.M. NF-κB, an Active Player in Human Cancers. Cancer Immunol. Res. 2014, 2, 823–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Delft, M.A.M.; Huitema, L.F.A.; Tas, S.W. The contribution of NF-κB signalling to immune regulation and tolerance. Eur. J. Clin. Investig. 2015, 45, 529–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duman, R.S.; Li, N. A neurotrophic hypothesis of depression: Role of synaptogenesis in the actions of NMDA receptor antago-nists. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 2475–2484. [Google Scholar] [CrossRef] [Green Version]
- Lindholm, J.S.O.; Castrén, E. Mice with altered BDNF signaling as models for mood disorders and antidepressant effects. Front. Behav. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [Green Version]
- Caviedes, A.; Lafourcade, C.; Soto, C.; Wyneken, U. BDNF/NF-κB Signaling in the Neurobiology of Depression. Curr. Pharm. Des. 2017, 23, 3154–3163. [Google Scholar] [CrossRef] [PubMed]
- Pizzi, M.; Spano, P.F. Distinct roles of diverse nuclear factor-κB complexes in neuropathological mechanisms. Eur. J. Pharmacol. 2006, 545, 22–28. [Google Scholar] [CrossRef]
- Meffert, M.K.; Baltimore, D. Physiological functions for brain NF-κB. Trends Neurosci. 2005, 28, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Napetschnig, J.; Wu, H. Molecular Basis of NF-κB Signaling. Annu. Rev. Biophys. 2013, 42, 443–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karin, M.; Ben-Neriah, Y. Phosphorylation Meets Ubiquitination: The Control of NF-κB Activity. Annu Rev. Immunol. 2000, 18, 621–663. [Google Scholar] [CrossRef] [PubMed]
- Cardinez, C.; Miraghazadeh, B.; Tanita, K.; Da Silva, E.; Hoshino, A.; Okada, S.; Chand, R.; Asano, T.; Tsumura, M.; Yoshida, K.; et al. Gain-of-function IKBKB mutation causes human combined immune deficiency. J. Exp. Med. 2018, 215, 2715–2724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bialek, K.; Czarny, P.; Watala, C.; Wigner, P.; Talarowska, M.; Galecki, P.; Szemraj, J.; Sliwinski, T. Novel association between TGFA, TGFB1, IRF1, PTGS2 and IKBKB single-nucleotide polymorphisms and occurrence, severity and treatment response of major depressive disorder. PeerJ 2020, 8, e8676. [Google Scholar] [CrossRef]
- Frank, M.G.; Hershman, S.A.; Weber, M.D.; Watkins, L.R.; Maier, S.F. Chronic exposure to exogenous glucocorticoids primes micro-glia to pro-inflammatory stimuli and induces NLRP3 mRNA in the hippocampus. Psychoneuroendocrinology 2014, 40, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Han, A.; Yeo, H.; Park, M.J.; Kim, S.H.; Choi, H.J.; Hong, C.W.; Kwon, M.S. IL-4/10 prevents stress vulnerability following imipramine dis-continuation. J. Neuroinflamm. 2015, 12, 1–16. [Google Scholar] [CrossRef] [Green Version]
- You, Z.; Luo, C.; Zhang, W.; Chen, Y.; He, J.; Zhao, Q.; Zuo, R.; Wu, Y. Pro- and anti-inflammatory cytokines expression in rat’s brain and spleen exposed to chronic mild stress: Involvement in depression. Behav. Brain Res. 2011, 225, 135–141. [Google Scholar] [CrossRef]
- Vogelzangs, N.; Duivis, H.E.; Beekman, A.; Kluft, C.; Neuteboom, J.; Hoogendijk, W.; Smit, J.H.; De Jonge, P.; Penninx, B.W.J.H. Association of depressive disorders, depression characteristics and antidepressant medication with inflammation. Transl. Psychiatry 2012, 2, e79. [Google Scholar] [CrossRef] [Green Version]
- The 100 Most Important Chemical Compounds: A Reference Guide; ABC-CLIO: Santa Barbara, CA, USA, 2008.
- Westenberg, H.G.M.; Sandner, C. Tolerability and safety of fluvoxamine and other antidepressants. Int. J. Clin. Pract. 2006, 60, 482–491. [Google Scholar] [CrossRef] [Green Version]
- Papp, M. Models of Affective Illness: Chronic Mild Stress in the Rat. Curr. Protoc. Pharmacol. 2012, 57, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Wojdacz, T.K.; Dobrovic, A.; Hansen, L.L. Methylation-sensitive high-resolution melting. Nat. Protoc. 2008, 3, 1903. [Google Scholar] [CrossRef] [PubMed]
- Wojdacz, T.K.; Dobrovic, A. Methylation-sensitive high resolution melting (MS-HRM): A new approach for sensitive and high-throughput assessment of methylation. Nucleic Acids Res. 2007, 35, e41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreos, R.; Ambrosini, G.; Groux, R.; Périer, R.C.; Bucher, P. The eukaryotic promoter database in its 30th year: Focus on non-vertebrate organisms. Nucleic Acids Res. 2017, 45, D51–D55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojdacz, T.K.; Borgbo, T.; Hansen, L.L. Primer design versus PCR bias in methylation independent PCR amplifications. Epigenetics 2009, 4, 231–234. [Google Scholar] [CrossRef] [Green Version]
- Papp, M.; Gruca, P.; Lason, M.; Niemczyk, M.; Willner, P. The role of prefrontal cortex dopamine D2 and D3 receptors in the mech-anism of action of venlafaxine and deep brain stimulation in animal models of treatment-responsive and treatment-resistant depression. J. Psychopharmacol. 2019, 33, 748–756. [Google Scholar] [CrossRef]
- Papp, M.; Gruca, P.; Lason-Tyburkiewicz, M.; Litwa, E.; Niemczyk, M.; Tota-Glowczyk, K.; Willner, P. Dopaminergic mechanisms in memory consolidation and antidepressant reversal of a chronic mild stress-induced cognitive impairment. Psychopharmacology 2017, 234, 2571–2585. [Google Scholar] [CrossRef] [Green Version]
- Wray, N.R.; Ripke, S.; Mattheisen, M.; Trzaskowski, M.; Byrne, E.M.; Abdellaoui, A.; Adams, M.J.; Agerbo, E.; Air, T.M.; Andlauer, T.M.F.; et al. Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nat. Genet. 2018, 50, 668–681. [Google Scholar] [CrossRef] [Green Version]
- Myint, A.-M.; Leonard, B.E.; Steinbusch, H.W.; Kim, Y.-K. Th1, Th2, and Th3 cytokine alterations in major depression. J. Affect. Disord. 2005, 88, 167–173. [Google Scholar] [CrossRef]
- Lee, K.-M.; Kim, Y.-K. The role of IL-12 and TGF-β1 in the pathophysiology of major depressive disorder. Int. Immunopharmacol. 2006, 6, 1298–1304. [Google Scholar] [CrossRef]
- Kunzmann, S.; Mantel, P.-Y.; Wohlfahrt, J.G.; Akdis, M.; Blaser, K.; Schmidt-Weber, C.B. Histamine enhances TGF-β1-mediated sup-pression of Th2 responses. FASEB J. 2003, 17, 1089–1095. [Google Scholar] [CrossRef]
- Fallon, J.; Reid, S.; Kinyamu, R.; Opole, I.; Opole, R.; Baratta, J.; Korc, M.; Endo, T.L.; Duong, A.; Nguyen, G.; et al. In vivo induction of massive proliferation, directed migration, and differentiation of neural cells in the adult mammalian brain. Proc. Natl. Acad. Sci. USA 2000, 97, 14686–14691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gałecki, P.; Gałecka, E.; Maes, M.; Chamielec, M.; Orzechowska, A.; Bobińska, K.; Lewiński, A.; Szemraj, J. The expression of genes encoding for COX-2, MPO, iNOS, and sPLA2-IIA in patients with recurrent depressive disorder. J. Affect. Disord. 2012, 138, 360–366. [Google Scholar] [CrossRef]
- Chen, Q.; Luo, Y.; Kuang, S.; Yang, Y.; Tian, X.; Ma, J.; Mai, S.; Xue, L.; Yang, J. Cyclooxygenase-2 Signalling Pathway in the Cortex is Involved in the Pathophysiological Mechanisms in the Rat Model of Depression. Sci. Rep. 2017, 7, 488. [Google Scholar] [CrossRef] [Green Version]
- Myint, A.M.; Steinbusch, H.W.; Goeghegan, L.; Luchtman, D.; Kim, Y.K.; Leonard, B.E. Effect of the COX-2 Inhibitor Celecoxib on Behavioural and Immune Changes in an Olfactory Bulbectomised Rat Model of Depression. Neuroimmunomodulation 2007, 14, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, S.-H.; Hosseini, F.; Modabbernia, A.; Ashrafi, M.; Akhondzadeh, S. Effect of celecoxib add-on treatment on symptoms and serum IL-6 concentrations in patients with major depressive disorder: Randomized double-blind placebo-controlled study. J. Affect. Disord. 2012, 141, 308–314. [Google Scholar] [CrossRef]
- Dong, S.Q.; Zhang, Q.P.; Zhu, J.X.; Chen, M.; Li, C.F.; Liu, Q.; Geng, D.; Yi, L.T. Gypenosides reverses depressive behavior via inhibiting hippo-campal neuroinflammation. Biomed. Pharm. 2018, 106, 1153–1160. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.M.; Niu, L.; Wang, L.L.; Bai, L.; Fang, X.Y.; Li, Y.C.; Yi, L.T. Berberine attenuates depressive-like behaviors by suppressing neu-ro-inflammation in stressed mice. Brain Res. Bull. 2017, 134, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Mehler, M.F. Epigenetic principles and mechanisms underlying nervous system functions in health and disease. Prog. Neurobiol. 2008, 86, 305–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Héberlé, É.; Bardet, A.F. Sensitivity of transcription factors to DNA methylation. Essays Biochem. 2019, 63, 727–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojdacz, T.K.; Hansen, L.L. Reversal of PCR bias for improved sensitivity of the DNA methylation melting curve assay. Biotech. 2006, 41, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Nicolia, V.; Cavallaro, R.A.; López-González, I.; Maccarrone, M.; Scarpa, S.; Ferrer, I.; Fuso, A. DNA Methylation Profiles of Selected Pro-Inflammatory Cytokines in Alzheimer Disease. J. Neuropathol. Exp. Neurol. 2017. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment Start | ||||
|---|---|---|---|---|
| 5 weeks adaptation to 1% sucrose consumption test | ||||
| 2 weeks without stress | 2 weeks of initial stress | |||
| 5 weeks without stress and with venlafaxine administration | 5 weeks of stress with saline administration | 5 weeks of stress with venlafaxine administration | ||
| Stress Procedure | ||||
| Stressor | Duration | Number of periods | ||
| Food and water deprivation | 10–14 h | 2 periods | ||
| 45-degree cage tilt | 10–14 h | 2 periods | ||
| Soiled cage (250 mL water in sawdust bedding) | 10–14 h | 2 periods | ||
| Paired housing | 10–14 h | 1 period | ||
| Low-intensity stroboscopic illumination (150 flashes/min) | 10–14 h | 2 periods | ||
| Intermittent illumination | 10–14 h (light on and off every two hours) | 2 periods | ||
| No stress | 10–14 h | 3 periods | ||
| Final sucrose consumption test and decapitation | ||||
| Gene | Starter Sequence (5′->3′) | Tm (°C) | Product Size (bp) | Number of CpG Islands | Product %CGs | CpGs in Product |
|---|---|---|---|---|---|---|
| IKBKB | F:AGGGTGGTTTTTTATTTTTATTTT R:AACCCCCACTAAAACTAACTTAA | 55 | 117 | 1 | 36.75 | 5 |
| IRF1 | F:TTGGAGATTTAGGGAGTTAGGT R:CCCCTTACCTATCTTAAAAAACC | 55 | 123 | 1 | 43.90 | 4 |
| PTGS2 | F:GTAATAGTAGGGAGGAAAAATTTTAA R:ATCCTAACAAACCCCAAA | 55 | 111 | 1 | 37.84 | 10 |
| TGFA | F:GTTTTTTTAGGTGGTTGGTTAAG R:CTTCAAACACCTCCCTACAATA | 55 | 188 | 1 | 42.55 | 11 |
| Weeks of CMS | Control | Stressed | Stressed/Saline | Stressed/Venlafaxine | Control/Venlafaxine |
|---|---|---|---|---|---|
| Week 0 | 12.6 ± 1.6 | 11.0 ± 0.7 | 11.7 ± 0.7 | 11.4 ± 0.5 | 11.9 ± 0.7 |
| Week 2 | 15.6 ± 1.9 | 6.8 ± 1.0 ** | 4.9 ± 0.6 **** | 5.8 ± 0.5 * | 13.9 ± 0.9 |
| Week 7 | - | - | 6.1 ± 0.7 | 12.6 ± 1.0 *** | 13.3 ± 1.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bialek, K.; Czarny, P.; Wigner, P.; Synowiec, E.; Barszczewska, G.; Bijak, M.; Szemraj, J.; Niemczyk, M.; Tota-Glowczyk, K.; Papp, M.; et al. Chronic Mild Stress and Venlafaxine Treatment Were Associated with Altered Expression Level and Methylation Status of New Candidate Inflammatory Genes in PBMCs and Brain Structures of Wistar Rats. Genes 2021, 12, 667. https://doi.org/10.3390/genes12050667
Bialek K, Czarny P, Wigner P, Synowiec E, Barszczewska G, Bijak M, Szemraj J, Niemczyk M, Tota-Glowczyk K, Papp M, et al. Chronic Mild Stress and Venlafaxine Treatment Were Associated with Altered Expression Level and Methylation Status of New Candidate Inflammatory Genes in PBMCs and Brain Structures of Wistar Rats. Genes. 2021; 12(5):667. https://doi.org/10.3390/genes12050667
Chicago/Turabian StyleBialek, Katarzyna, Piotr Czarny, Paulina Wigner, Ewelina Synowiec, Gabriela Barszczewska, Michal Bijak, Janusz Szemraj, Monika Niemczyk, Katarzyna Tota-Glowczyk, Mariusz Papp, and et al. 2021. "Chronic Mild Stress and Venlafaxine Treatment Were Associated with Altered Expression Level and Methylation Status of New Candidate Inflammatory Genes in PBMCs and Brain Structures of Wistar Rats" Genes 12, no. 5: 667. https://doi.org/10.3390/genes12050667
APA StyleBialek, K., Czarny, P., Wigner, P., Synowiec, E., Barszczewska, G., Bijak, M., Szemraj, J., Niemczyk, M., Tota-Glowczyk, K., Papp, M., & Sliwinski, T. (2021). Chronic Mild Stress and Venlafaxine Treatment Were Associated with Altered Expression Level and Methylation Status of New Candidate Inflammatory Genes in PBMCs and Brain Structures of Wistar Rats. Genes, 12(5), 667. https://doi.org/10.3390/genes12050667