
The Genotoxic and Pro-Apoptotic Activities of Advanced Glycation End-Products (MAGE) Measured with Micronuclei Assay Are Inhibited by Their Low Molecular Mass Counterparts


 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
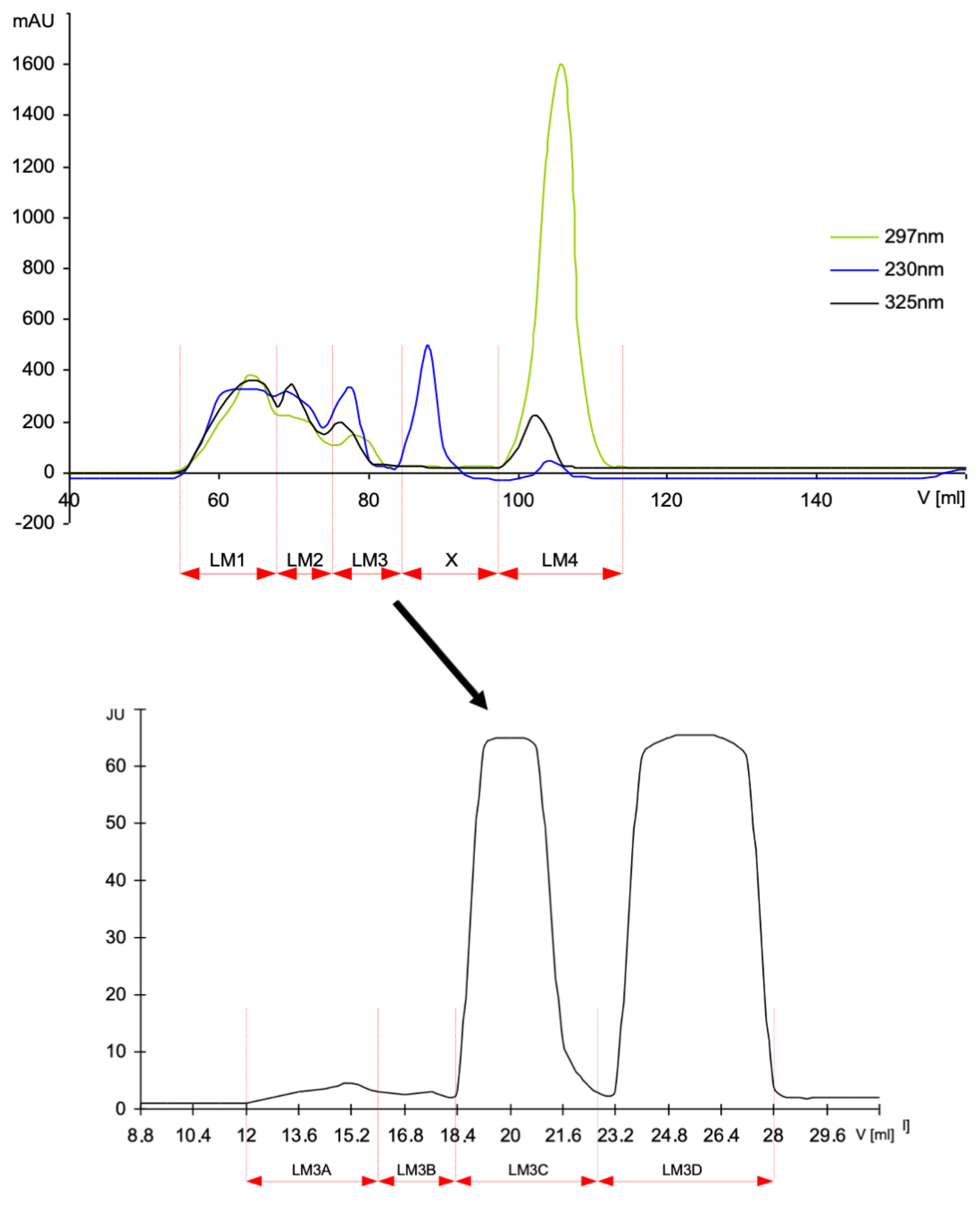
2.2. Preparation and Fractionation of HMW–MAGE and LMW–MAGE
2.3. Cell Culture and Treatment with MAGE
2.4. Micronucleus Frequency Assay
2.5. Irradiation Experiments
2.6. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Melpomeni, P.; Vlassara, H. Advanced glycation end products and diabetic complications: A general overview. Hormones 2005, 4, 28–37. [Google Scholar]
- Vlassara, H.; Uribarri, J.; Ferrucci, L.; Cai, W.; Torreggiani, M.; Post, J.B.; Zheng, F.; Striker, G.E. Identifying advanced glycation end products as a major source of oxidants in aging: Implications for the management and/or prevention of reduced renal function in elderly persons. Semin. Nephrol. 2009, 29, 594–603. [Google Scholar] [CrossRef] [Green Version]
- Urlich, P.; Cerami, A. Protein glycation, diabetes and aging. Recent Prog. Horm. Res. 2001, 56, 1–21. [Google Scholar]
- Zhang, Q.; Ames, J.M.; Richard, D.; Smith, R.D.; Baynes, J.W.; Metz, T.O. A perspective on the Maillard reaction and the analysis of protein glycation by mass spectrometry: Probing the pathogenesis of chronic disease. J. Proteome Res. 2009, 8, 754–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, M.; Wells-Knecht, K.J.; Blackledge, J.A.; Lyons, T.J.; Thorpe, S.R.; Baynes, J.W. Glycation, glycoxidation, and cross-linking of collagen by glucose. Kinetics, mechanisms, and inhibition of late stages of the Maillard reaction. Diabetes 1994, 43, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Shipanova, I.N.; Glomb, M.A.; Nagaraj, R.H. Protein modification by methylglyoxal: Chemical nature and synthetic mechanism of a major fluorescent adduct. Arch. Biochem. Biophys. 1997, 344, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Al-Abed, Y.; Kapurniotu, A.; Bucala, R. Advanced glycation end products: Detection and reversal. Methods Enzymol. 1999, 309, 152–171. [Google Scholar] [PubMed]
- Nagai, R.; Hayashi, C.M.; Xia, L.; Takeya, M.; Horiuchi, S. Identification in human atherosclerotic lesions of GA-pyridine, a novel structure derived from glycolaldehyde-modified proteins. J. Biol. Chem. 2002, 727, 48905–48912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietkiewicz, J.; Gamian, A.; Staniszewska, M.; Danielewicz, R. Inhibition of human muscle-specific enolase by methylglyoxal and irreversible formation of advanced glycation end-products. J. Enz. Inhib. Med. Chem. 2009, 24, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Pietkiewicz, J.; Seweryn, E.; Bartyś, A.; Gamian, A. Receptors for advanced glycation end products and their physiological and clinical significance. Post. Hig. Med. Dosw. 2008, 62, 511–523. [Google Scholar]
- Kaji, Y.; Amano, S.; Usui, T.; Oshika, T.; Yamashiro, K.; Ishida, S.; Suzuki, K.; Tanaka, S.; Adamis, A.P.; Nagai, R.; et al. Expression and function of receptors for advanced glycation end products in bovine corneal endothelial cells. Invest. Ophthalmol. Vis. Sci. 2003, 44, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Riuzzi, F.; Sorci, G.; Donato, R. The amphoterin (HMGB1)/receptor for advanced glycation end products (RAGE) pair modulates myoblast proliferation, apoptosis, adhesiveness, migration, and invasiveness. Functional inactivation of RAGE in L6 myoblast results in tumor formation in vivo. J. Biol. Chem. 2006, 28, 8242–8253. [Google Scholar] [CrossRef] [Green Version]
- Šebeková, K.; Wagner, Z.; Schupp, N.; Boor, P. Genomic damage and malignancy in end-stage renal failure: Do advanced glycation end products contribute? Kidney Blood Press. Res. 2007, 30, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Kuniyasu, H.; Chihara, Y.; Kondo, H. Differential effects between amphoterin and advanced glycation end products on colon cancer cells. Int. J. Cancer 2003, 104, 722–727. [Google Scholar] [CrossRef] [PubMed]
- Stopper, H.; Schinzel, R.; Sebekova, K.; Heidland, A. Genotoxicity of advanced glycation end products in mammalian cells. Cancer Lett. 2003, 190, 151–156. [Google Scholar] [CrossRef]
- Staniszewska, M.; Bronowicka-Szydełko, A.; Gostomska-Pampuch, K.; Szkudlarek, J.; Bartyś, A.; Bieg, T.; Gamian, E.; Kochman, A.; Picur, B.; Pietkiewicz, J.; et al. The melibiose-derived glycation product mimics a unique epitope present in human and animal tissues. Sci. Rep. 2021. [Google Scholar] [CrossRef]
- Staniszewska, M.; Jarosz, S.; Jon, M.; Gamian, A. Advanced glycation end-products prepared in solution under high pressure contain epitopes distinct from those formed in the dry reaction at high temperature. Arch. Immunol. Ther. Exp. 2005, 53, 71–78. [Google Scholar]
- Boratyński, J.; Roy, R. High temperature conjugation of proteins with carbohydrates. Glycoconj. J. 1998, 15, 131–138. [Google Scholar] [CrossRef]
- Shamsi, F.A.; Partal, A.; Sady, C.; Glomb, M.A.; Nagaraj, R.H. Immunological evidence for methylglyoxal-derived modifications in vivo. Determination of antigenic epitopes. J. Biol. Chem. 1998, 273, 6928–6936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenech, M.; Morley, A.A. Measurement of micronuclei in lymphocytes. Mutation Res. 1985, 147, 29–36. [Google Scholar] [CrossRef]
- Price, C.L.; Knight, S.C. Methylglyoxal: Possible link between hyperglycaemia and immune suppression? Trends Endocrinol. Metab. 2009, 20, 312–317. [Google Scholar] [CrossRef]
- Otton, R.; Soriano, F.G.; Verlengia, R.; Curi, R. Diabetes induces apoptosis in lymphocytes. J. Endocrinol. 2004, 182, 145–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakowicz-Burkiewicz, M.; Kocbuch, K.; Grden, M.; Szutowicz, A.; Pawelczyk, T. Diabetes-induced decrease of adenosine kinase expression impairs the proliferation potential of diabetic rat T lymphocytes. Immunology 2006, 118, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Schupp, N.; Dette, E.M.; Schmid, U.; Bahner, U.; Winkler, M.; Heidland, A.; Stopper, H. Benfotiamine reduces genomic damage in peripheral lymphocytes of hemodialysis patients. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2008, 378, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Haque, E.; Kamil, M.; Hasan, A.; Irfan, S.; Sheikh, S.; Khatoon, A.; Nazir, A.; Mir, S.S. Advanced glycation end products (AGEs), protein aggregation and their cross talk: New insight in tumorigenesis. Glycobiology 2019, 30, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Hagmar, L.; Stromberg, U.; Tinnerberg, H.; Mikoczy, Z. Epidemiological evaluation of cytogenetic biomarkers as potential surrogate end-points for cancer. IARC Sci. Publ. 2004, 157, 207–215. [Google Scholar]



{kind=link}
{kind=link}
| Tested Compounds | Concentration (μg/mL) | Frequencies of MN/1000 CBL | t-Welch Test t | p-Value | Fraction of Damaged Cells (Promille) | t-Welch Test t | p-Value |
|---|---|---|---|---|---|---|---|
| Negative control | - | 20.66 ± 1.39 | - | - | 19.3 ± 1.84 | - | - |
| BSA monomer | 100 | 18.00 ± 1.61 | −27.964 | <0.005 | 16.3 ± 1.52 | −28.107 | <0.005 |
| Melibiose | 100 | 18.61 ± 1.69 | −20.948 | <0.005 | 17.8 ± 1.24 | −15.117 | <0.005 |
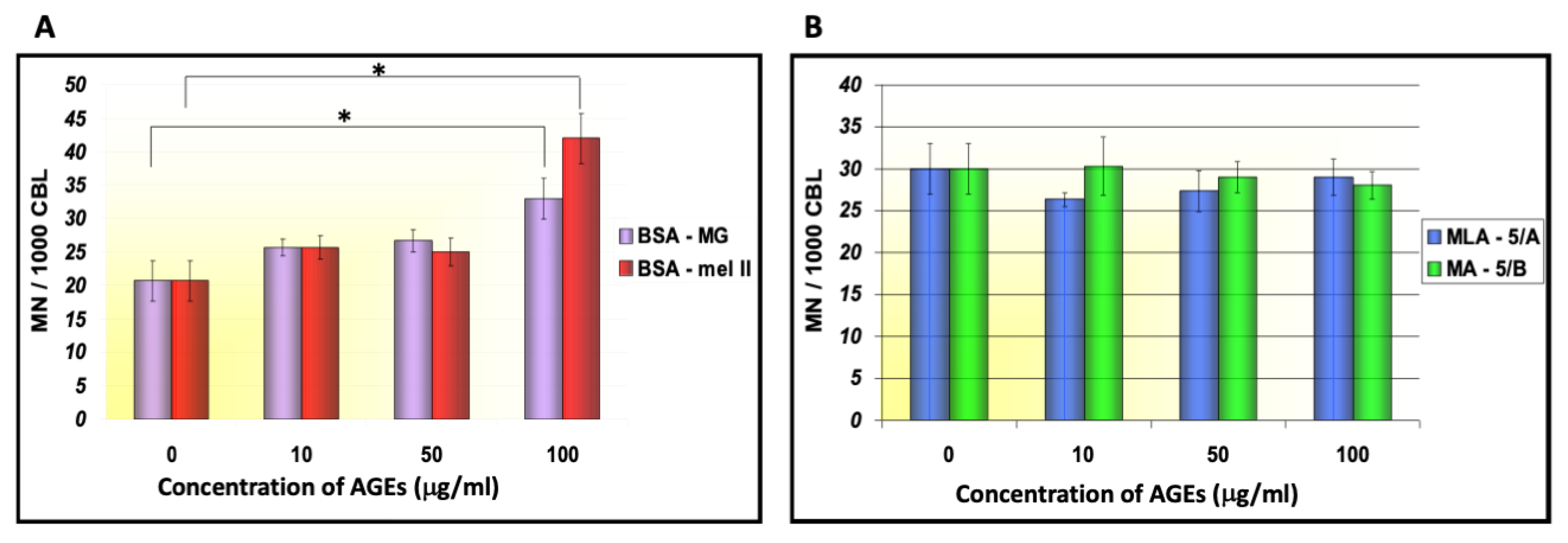
| BSA–mel fr. II HTG | 100 | 40.22 ± 5.34 | 79.264 | <0.005 | 38.0 ± 5.12 | 76.857 | <0.005 |
| BSA–mel fr. II HTG | 50 | 25.70 ± 1.87 | 48.368 | <0.005 | 24.6 ± 2.82 | 35.196 | <0.005 |
| BSA–MG | 100 | 28.80 ± 6.50 | 27.383 | <0.005 | 29.3 ± 4.09 | 49.858 | <0.005 |
| BSA–MG | 50 | 24.89 ± 5.26 | 17.385 | <0.005 | 21.5 ± 2.88 | 14.394 | <0.005 |
| LM 3C | 100 | 17.30 ± 1.25 | −40.191 | <0.005 | 17.0 ± 1.01 | −24.502 | <0.005 |
| LMA 5A | 100 | 19.20 ± 1.63 | −15.240 | <0.005 | 17.4 ± 1.66 | −17.144 | <0.005 |
| Negative control | - | 29.25 ± 2.77 | - | - | 27.00 ± 0.71 | ||
| Bleomycin | 10 | 105.50 ± 7.76 | 206.93 | <0.005 | 82.50 ± 5.22 | 235.57 | <0.005 |
| N-α-acetyl-L-lysine | 100 | 30.20 ± 2.99 | 5.21 | <0.005 | 26.20 ± 1.47 | −10.95 | <0.005 |
| NN-α-acetyl-L-arginine | 100 | 28.46 ± 2.21 | −4.98 | <0.005 | 26.00 ± 1.51 | −13.40 | <0.005 |
| Melibiose | 100 | 27.75 ± 1.92 | −9.95 | <0.005 | 26.40 ± 2.01 | −6.29 | <0.005 |
| LMA 5A | 10 | 26.33 ± 0.94 | −22.32 | <0.005 | 24.67 ± 1.89 | −25.80 | <0.005 |
| 50 | 27.50 ± 2.06 | −11.33 | <0.005 | 25.75 ± 2.86 | −9.48 | <0.005 | |
| 100 | 30.25 ± 1.79 | 6.78 | <0.005 | 28.25 ± 2.05 | 12.88 | <0.005 | |
| LMA 5B | 10 | 30.33 ± 4.03 | 4.93 | <0.005 | 28.30 ± 2.62 | 10.71 | <0.005 |
| 50 | 29.00 ± 2.16 | −1.59 | 0.1118 | 25.33 ± 1.25 | −25.98 | <0.005 | |
| 100 | 27.50 ± 1.66 | −12.11 | <0.005 | 25.00 ± 1.87 | −22.36 | <0.005 |
| Tested Compound | Concentration (μg/mL) | Frequency of MN/1000 CBL | t-Welch Test t | p-Value | Fraction of Damaged Cells (promille) | t-Welch Test t | p-Value |
|---|---|---|---|---|---|---|---|
| Negative control | - | 23.30 ± 1.24 | - | - | 21.60 ± 0.94 | - | - |
| BSA–mel-II | 100 | 41.33 ± 2.86 | 129.33 | <0.005 | 36.00 ± 2.94 | 104.32 | <0.005 |
| BSA–mel-II + LM 3C | 100 100 | 42.66 ± 4.49 | 92.94 | <0.005 | 36.10 ± 3.29 | 94.76 | <0.005 |
| BSA–mel-II + LMA 5A | 100 100 | 42.00 ± 2.94 | 131.05 | <0.005 | 36.33 ± 2.05 | 146.05 | <0.005 |
| BSA MG | 100 | 32.33 ± 1.69 | 96.33 | <0.005 | 29.66 ± 1.24 | 115.83 | <0.005 |
| BSA–MG + LM 3C | 100 100 | 33.70 ± 1.69 | 110.94 | <0.005 | 30.00 ± 1.41 | 110.84 | <0.005 |
| BSA–MG + LM 2D/2 | 100 100 | 33.86 ± 2.04 | 98.91 | <0.005 | 30.10 ± 0.82 | 152.37 | <0.005 |
| Tested Compounds | Concentration (μg/mL) | Frequency of MN/1000 CBL | t-Welch Test t | p-Value | Fraction of Damaged Cells (promille) | t-Welch Test t | p-Value |
|---|---|---|---|---|---|---|---|
| Negative control | - | 28.35 ± 3.49 | - | - | 25.50 ± 2.04 | - | - |
| X-rays (2 Gy) | - | 322.50 ± 21.61 | 300.47 | <0.005 | 287.25 ± 25.58 | 228.08 | <0.005 |
| LMA 5A + irradiation | 1 | 310.17 ± 12.01 | 503.86 | <0.005 | 268.05 ± 30.26 | 178.83 | <0.005 |
| 10 | 297.72 ± 10.11 | 563.16 | <0.005 | 241.00 ± 15.05 | 317.28 | <0.005 | |
| 100 | 289.25 ± 13.63 | 414.64 | <0.005 | 237.50 ± 12.22 | 382.63 | <0.005 | |
| LMA 5B + irradiation | 1 | 341.00 ± 11.37 | 587.8 | <0.005 | 277.60 ± 12.04 | 461.62 | <0.005 |
| 10 | 298.00 ± 14.9 | 394 | <0.005 | 244.36 ± 8.50 | 559.85 | <0.005 | |
| 100 | 297.75 ± 10.22 | 557.8 | <0.005 | 245.50 ± 8.77 | 546.34 | <0.005 | |
| N-α-acetyl-L-lysine + irradiation | 1 | 313.00 ± 21.33 | 294.49 | <0.005 | 252.10 ± 15.51 | 323.9 | <0.005 |
| 10 | 337.44 ± 20.42 | 333.63 | <0.005 | 266.00 ± 20.61 | 259.66 | <0.005 | |
| 100 | 310.33 ± 7.89 | 730.84 | <0.005 | 251.00 ± 8.48 | 578.12 | <0.005 | |
| Melibiose + irradiation | 1 | 319.00 ± 10.71 | 576.97 | <0.005 | 249.67 ± 8.34 | 583.82 | <0.005 |
| 10 | 340.62 ± 10.66 | 622.51 | <0.005 | 264.50 ± 11.94 | 441.19 | <0.005 | |
| 100 | 298.30 ± 22.66 | 263.28 | <0.005 | 240.67 ± 14.29 | 333.31 | <0.005 | |
| N-α-acetyl-L-arginine + irradiation | 1 | 332.00 ± 16.39 | 405.18 | <0.005 | 261.10 ± 13.34 | 390.38 | <0.005 |
| 10 | 330.00 ± 19.03 | 348.63 | <0.005 | 266.67 ± 15.82 | 338.08 | <0.005 | |
| 100 | 341.65 ± 17.40 | 394.76 | <0.005 | 262.00 ± 21.18 | 248.53 | <0.005 |
| Tested Compounds | Concentration (μg/mL) | Cells with Micronuclei (%) | Number of Micronuclei in 100 Cells | Cell Division Index NDI (%) | Apoptotic Cells (%) |
|---|---|---|---|---|---|
| Negative control | - | 2.9 | 2.9 | 2.1 | 3.3 |
| MB–mel ACG | 1 | 2.9 | 2.9 | 2.09 | 3.2 |
| 10 | 2.9 | 2.9 | 2.01 | 3.5 | |
| 20 | 2.8 | 2.8 | 2.07 | 3.5 | |
| 50 | 2.8 | 2.9 | 1.99 | 3.6 | |
| 100 | 3.3 | 4.1 | 2.01 | 3.5 | |
| MB–mel HTG | 1 | 3.4 | 3.4 | 2.04 | 3.5 |
| 10 | 4 | 4.2 | 1.95 | 3.8 | |
| 20 | 5.1 | 5.7 | 2.02 | 3.9 | |
| 50 | 5.7 | 6 | 2.09 | 4 | |
| 100 | 5.7 | 6.5 | 1.99 | 3.9 | |
| Melibiose | 100 | 2.8 | 2.9 | 2.1 | 3.4 |
| Myoglobin | 100 | 2.9 | 2.9 | 2.02 | 3 |
| Bleomycin | 10 | 13.5 | 19.5 | 1.33 | 5 |
| Tested Compounds | Concentration (μg/mL) | Cells with Micronuclei (%) | Number of Micronuclei in 100 Cells | Cell Division Index NDI (%) | Apoptotic Cells (%) |
|---|---|---|---|---|---|
| Negative control | - | 2.2 | 2.1 | 2.09 | 1.1 |
| MB–mel ACG | 1 | 2.1 | 2.1 | 2.00 | 1.1 |
| 10 | 2.2 | 1.6 | 1.99 | 1.2 | |
| 20 | 2.1 | 2.2 | 1.98 | 1.2 | |
| 50 | 2.2 | 2.4 | 2.04 | 1.2 | |
| 100 | 2.1 | 2.3 | 2.00 | 1.2 | |
| MB–mel HTG | 1 | 2.3 | 2.7 | 2.00 | 1.4 |
| 10 | 2.8 | 3.3 | 1.96 | 1.7 | |
| 20 | 3.0 | 3.6 | 2.03 | 1.6 | |
| 50 | 3.1 | 3.8 | 1.96 | 1.7 | |
| 100 | 3.3 | 4.3 | 1.95 | 2.4 | |
| Melibiose | 100 | 2.1 | 2.1 | 2.05 | 1.1 |
| Myoglobin | 100 | 2.1 | 2.2 | 2.08 | 1.2 |
| Bleomycin | 10 | 15.9 | 24.4 | 1.56 | 2.7 |
| Tested Compounds | Concentration (μg/mL) | Cells with Micronuclei (%) | Number of Micronuclei in 100 Cells | Cell Division Index NDI (%) | Apoptotic Cells (%) |
|---|---|---|---|---|---|
| Negative control | 0 | 1.5 | 1.2 | 1.94 | 1.3 |
| MB–mel ACG | 1 | 1.6 | 1.6 | 1.99 | 1.4 |
| 10 | 1.5 | 1.6 | 1.99 | 1.3 | |
| 20 | 1.5 | 1.6 | 1.93 | 1.4 | |
| 50 | 1.5 | 1.6 | 2.01 | 1.3 | |
| 100 | 1.7 | 1.8 | 2.01 | 1.5 | |
| MB–mel HTG | 1 | 1.8 | 1.8 | 1.99 | 1.3 |
| 10 | 3.5 | 3.8 | 1.98 | 1.3 | |
| 20 | 4.4 | 5.3 | 1.92 | 1.5 | |
| 50 | 4.9 | 5.2 | 1.95 | 1.4 | |
| 100 | 6.2 | 6.8 | 1.94 | 1.6 | |
| Melibiose | 100 | 1.5 | 1.5 | 2.01 | 1.3 |
| Myoglobin | 100 | 1.4 | 1.1 | 1.98 | 1.3 |
| Bleomycin | 10 | 11.6 | 15.2 | 1.37 | 2.3 |
| Concentration (μg/mL) | Cells with Micronuclei (%) | Cell Division Index NDI (%) | Apoptotic Cells (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| MB–mel HTG | LM 3C | |||||||||
| 0 | 0 | 1 | 2 | 1 | 1.92 | 1.97 | 1.84 | 0.8 | 0.6 | 0.5 |
| 10 | 0 | 2 | 2 | 1 | 2.04 | 1.95 | 1.92 | 0.7 | 0.5 | 0.8 |
| 20 | 0 | 4 | 4 | 5 | 1.92 | 1.94 | 1.86 | 0.9 | 1.2 | 0.9 |
| 50 | 0 | 5 | 4 | 6 | 2 | 1.92 | 1.94 | 1.5 | 1.7 | 1.4 |
| 0 | 10 | 1 | 1 | 2 | 1.96 | 2.08 | 2.05 | 0.5 | 0.5 | 0.7 |
| 0 | 20 | 2 | 1 | 1 | 2.08 | 1.9 | 1.92 | 0.4 | 0.6 | 0.6 |
| 0 | 50 | 1 | 2 | 2 | 1.95 | 1.92 | 1.98 | 0.6 | 0.5 | 0.8 |
| 10 | 10 | 2 | 1 | 1 | 1.87 | 1.95 | 1.94 | 0.5 | 0.7 | 0.7 |
| 20 | 20 | 3 | 2 | 2 | 1.98 | 2.1 | 1.95 | 0.6 | 0.6 | 0.7 |
| 50 | 50 | 3 | 3 | 2 | 2.04 | 1.96 | 1.94 | 0.5 | 0.7 | 0.5 |
| Bleomycin 10 μg/ml | 18 | 21 | 17 | 1.47 | 1.68 | 1.52 | 2.8 | 3 | 2.4 | |
| Concentration (μg/mL) | Cells with Micronuclei (%) | Number of Micronuclei in 100 Cells | Apoptotic Cells (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Negative control | - | 3.1 | 2.6 | 2.8 | 3.1 | 2.6 | 2.8 | 3.0 | 3.2 | 3.2 |
| MB–mel HTG | 20 | 6.0 | 5.2 | 5.7 | 6.4 | 5.5 | 6.1 | 3.8 | 4.1 | 3.6 |
| MB–mel HTG + LM 3A | 20 10 | 5.0 | 6.1 | 5.5 | 5.6 | 6.3 | 5.7 | 3.5 | 3.9 | 3.3 |
| MB–mel HTG + LM 3B | 20 10 | 4.0 | 4.8 | 4.4 | 4.2 | 4.8 | 4.8 | 3.1 | 3.3 | 3.3 |
| MB–mel HTG + LM 3C | 20 10 | 4.2 | 4.6 | 4.7 | 4.2 | 4.8 | 4.9 | 3.5 | 3.2 | 3.4 |
| Melibiose | 100 | 2.7 | 3.1 | 3.0 | 2.7 | 3.1 | 3.0 | 3.2 | 3.0 | 3.0 |
| Myoglobin | 100 | 2.6 | 2.8 | 3.1 | 2.6 | 2.8 | 3.1 | 3.0 | 3.2 | 2.9 |
| LM 3A | 10 | 3.0 | 3.0 | 2.9 | ||||||
| LM 3B | 10 | 2.8 | 2.8 | 3.1 | ||||||
| LM 3C | 10 | 3.0 | 3.0 | 2.9 | ||||||
| Concentration (μg/ ml) | Cells with Micronuclei (%) | Number of Micronuclei in 100 Cells | Apoptotic Cells (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 0 | 2.0 | 1.9 | 2.2 | 2.0 | 1.9 | 2.2 | 1.0 | 1.2 | 1.0 |
| MB–mel HTG | 20 | 3.4 | 3.8 | 3.3 | 3.6 | 4.1 | 3.5 | 1.4 | 1.8 | 2.0 |
| MB–mel HTG + LM 3A | 20 10 | 3.6 | 3.4 | 3.2 | 3.6 | 3.6 | 3.4 | 1.5 | 1.7 | 2.9 |
| MB–mel HTG + LM 3B | 20 10 | 2.6 | 2.8 | 3.0 | 2.6 | 3.0 | 3.4 | 1.1 | 1.3 | 1.1 |
| MB–mel HTG + LM 3C | 20 10 | 2.7 | 3.1 | 3.0 | 2.7 | 3.3 | 3.0 | 1.2 | 1.0 | 1.3 |
| Melibiose | 100 | 1.8 | 2.0 | 2.0 | 1.8 | 2.0 | 2.0 | 1.1 | 0.9 | 1.0 |
| Myoglobin | 100 | 2.0 | 2.2 | 2.0 | 2.0 | 2.2 | 2.0 | 1.0 | 1.0 | 1.2 |
| LM 3A | 10 | 1.9 | 1.9 | 0.9 | ||||||
| LM 3B | 10 | 2.2 | 2.2 | 1.1 | ||||||
| LM 3C | 10 | 2.0 | 2.0 | 1.0 | ||||||
| Concentration (μg/mL) | Cells with Micronuclei (%) | Number of Micronuclei in 100 Cells | Apoptotic Cells (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Negative control | 0 | 1.3 | 1.4 | 1.8 | 1.3 | 1.4 | 1.8 | 1.5 | 1.1 | 1.2 |
| MB–mel HTG | 20 | 4.5 | 4.2 | 4.8 | 5.5 | 5.2 | 5.8 | 1.5 | 1.8 | 1.6 |
| MB–mel HTG + LM 3A | 20 10 | 4.6 | 4.0 | 4.4 | 5.2 | 4.4 | 4.6 | 1.5 | 1.3 | 1.6 |
| MB–mel HTG + MB 3B | 20 10 | 3.8 | 3.6 | 3.5 | 4.0 | 4.2 | 3.8 | 1.4 | 1.1 | 1.4 |
| MB–mel HTG + LM 3C | 20 10 | 3.9 | 3.7 | 4.2 | 4.5 | 4.4 | 4.2 | 1.2 | 1.6 | 1.5 |
| Melibiose | 100 | 1.3 | 1.7 | 1.5 | 1.3 | 1.7 | 1.5 | 1.5 | 1.1 | 1.4 |
| Myoglobin | 100 | 1.6 | 1.6 | 1.8 | 1.6 | 1.6 | 1.8 | 1.0 | 1.4 | 1.2 |
| LM 3A | 10 | 1.4 | 1.4 | 1.2 | ||||||
| LM 3B | 10 | 1.7 | 1.7 | 1.0 | ||||||
| LM 3C | 10 | 1.4 | 1.5 | 1.4 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czech, M.; Konopacka, M.; Rogoliński, J.; Maniakowski, Z.; Staniszewska, M.; Łaczmański, Ł.; Witkowska, D.; Gamian, A. The Genotoxic and Pro-Apoptotic Activities of Advanced Glycation End-Products (MAGE) Measured with Micronuclei Assay Are Inhibited by Their Low Molecular Mass Counterparts. Genes 2021, 12, 729. https://doi.org/10.3390/genes12050729
Czech M, Konopacka M, Rogoliński J, Maniakowski Z, Staniszewska M, Łaczmański Ł, Witkowska D, Gamian A. The Genotoxic and Pro-Apoptotic Activities of Advanced Glycation End-Products (MAGE) Measured with Micronuclei Assay Are Inhibited by Their Low Molecular Mass Counterparts. Genes. 2021; 12(5):729. https://doi.org/10.3390/genes12050729
Chicago/Turabian StyleCzech, Monika, Maria Konopacka, Jacek Rogoliński, Zbigniew Maniakowski, Magdalena Staniszewska, Łukasz Łaczmański, Danuta Witkowska, and Andrzej Gamian. 2021. "The Genotoxic and Pro-Apoptotic Activities of Advanced Glycation End-Products (MAGE) Measured with Micronuclei Assay Are Inhibited by Their Low Molecular Mass Counterparts" Genes 12, no. 5: 729. https://doi.org/10.3390/genes12050729
APA StyleCzech, M., Konopacka, M., Rogoliński, J., Maniakowski, Z., Staniszewska, M., Łaczmański, Ł., Witkowska, D., & Gamian, A. (2021). The Genotoxic and Pro-Apoptotic Activities of Advanced Glycation End-Products (MAGE) Measured with Micronuclei Assay Are Inhibited by Their Low Molecular Mass Counterparts. Genes, 12(5), 729. https://doi.org/10.3390/genes12050729