
Analysis of Skin Pigmentation and Genetic Ancestry in Three Subpopulations from Pakistan: Punjabi, Pashtun, and Baloch

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples, DNA Extraction, and DNA Quantification
2.2. Measurement of Quantitative Skin Pigmentation
2.3. Typing with the Precision ID Ancestry Panel and the Human Pigmentation (HuPi) AmpliSeq™ Custom Panel
2.4. Analysis of Sequencing Data
2.5. Classification of Biogeographic Ancestry
2.6. Correlations between Biogeographic Ancestry, Pigmentary Variants, and Skin Pigmentation
2.7. Prediction of Skin Colour Using the HIrisPlex-S
2.8. Statistical Methods
3. Results
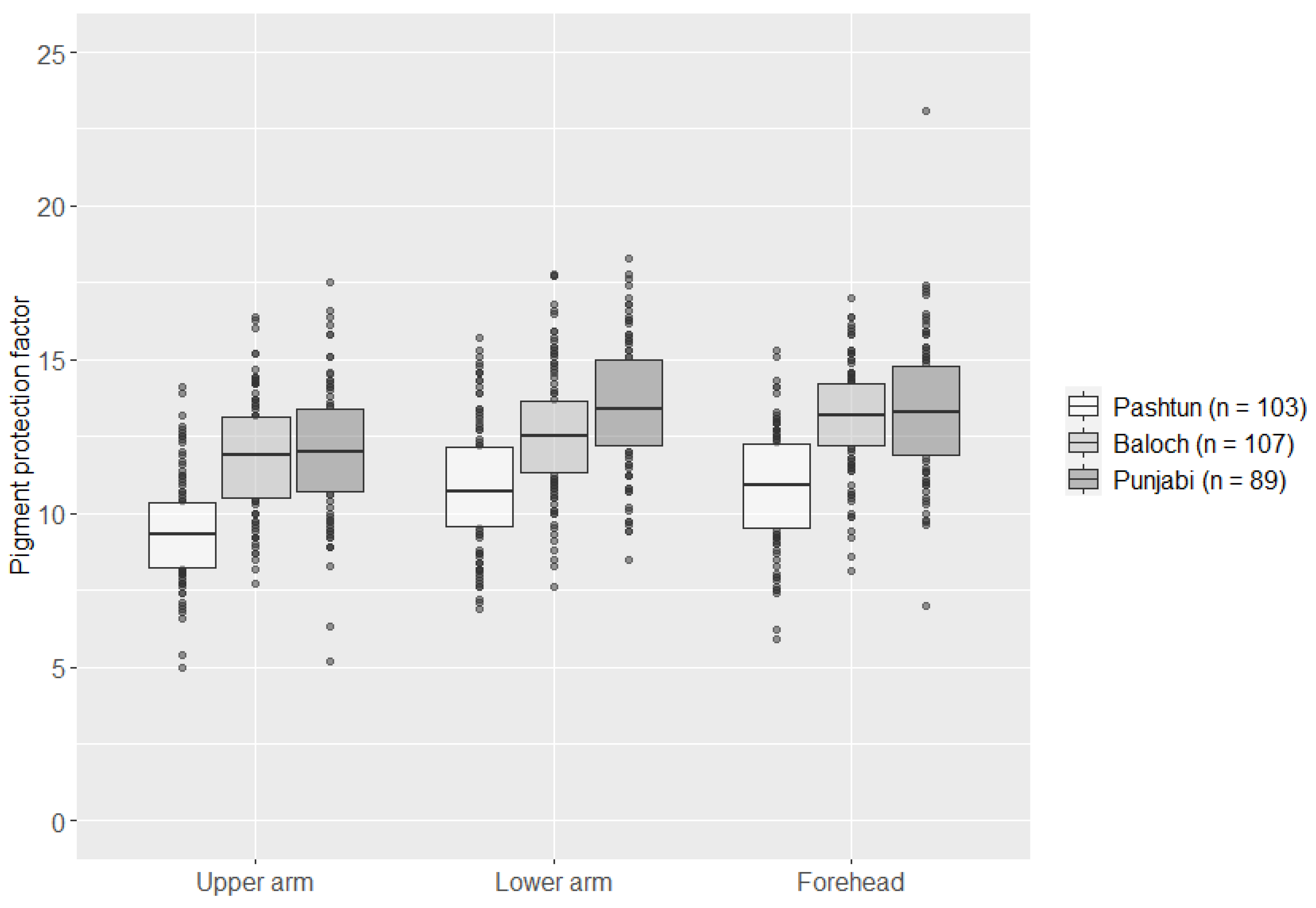
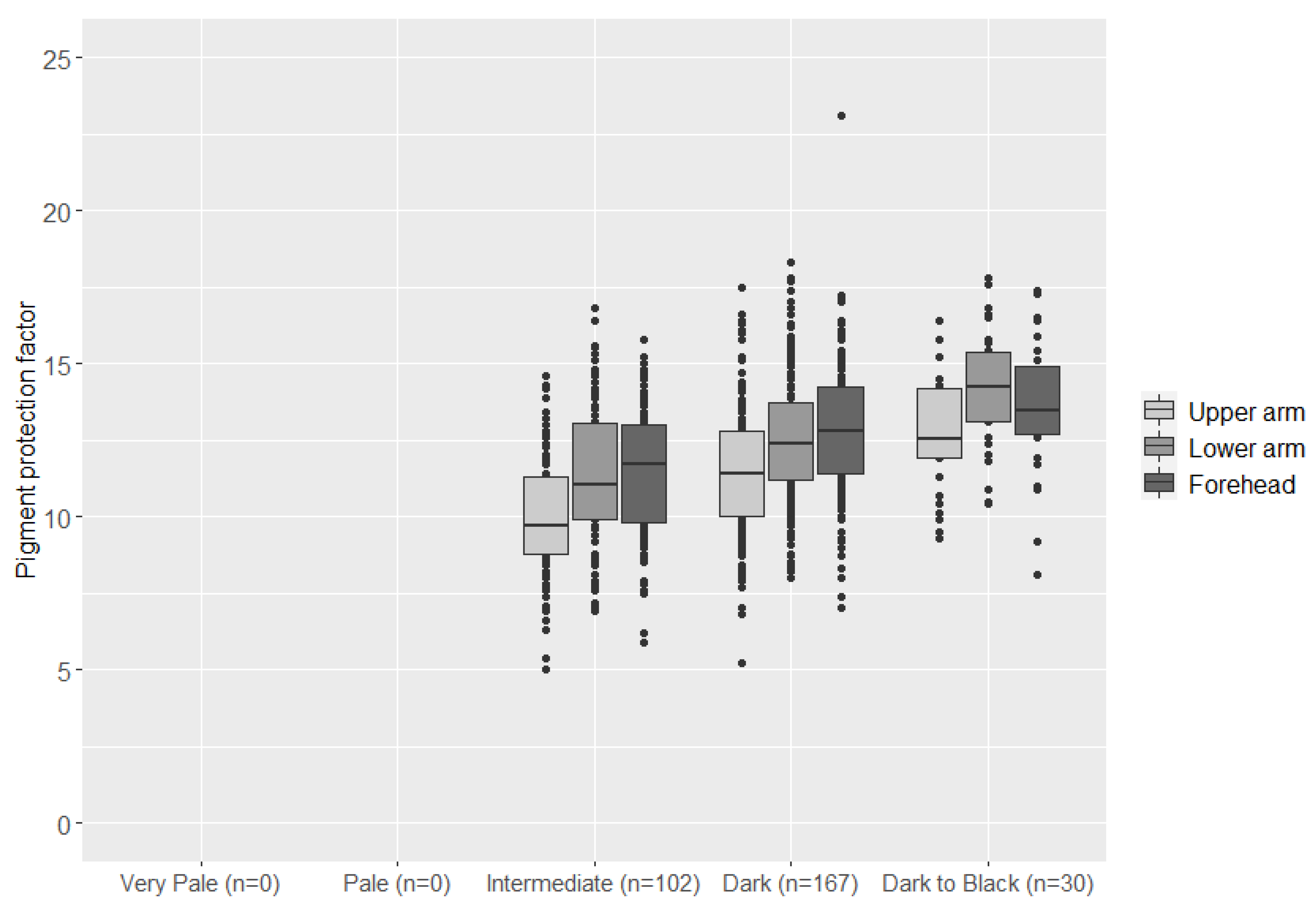
3.1. Skin Pigmentation Measurements
3.2. The Precision ID Ancestry Panel
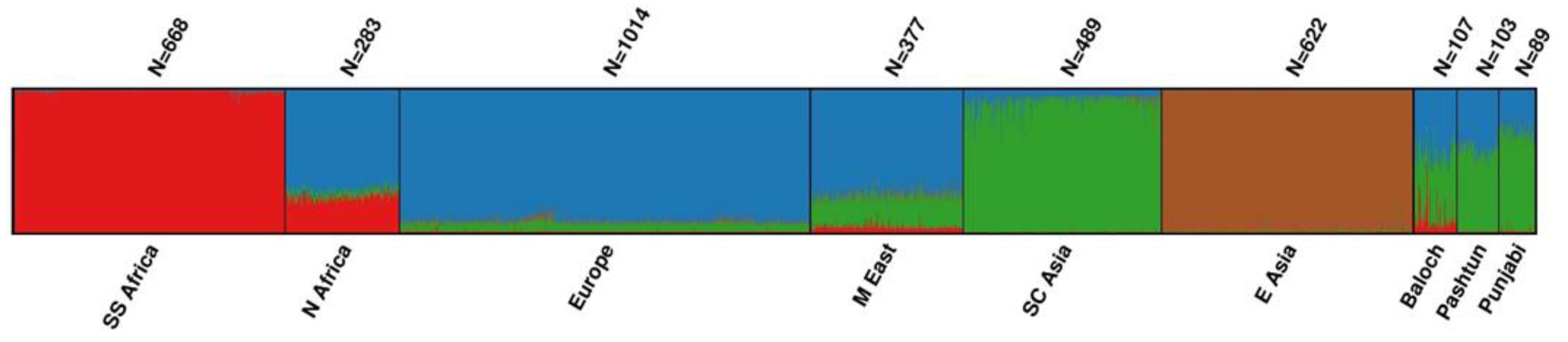
3.3. Proportion of Ancestry Components
3.4. The HuPi AmpliSeq™ Custom Panel
3.5. Correlation between Skin Pigmentation and Biogeographic Ancestry
3.6. Correlations among Skin Pigmentation, Pigmentary Variants, and Biogeographic Ancestry
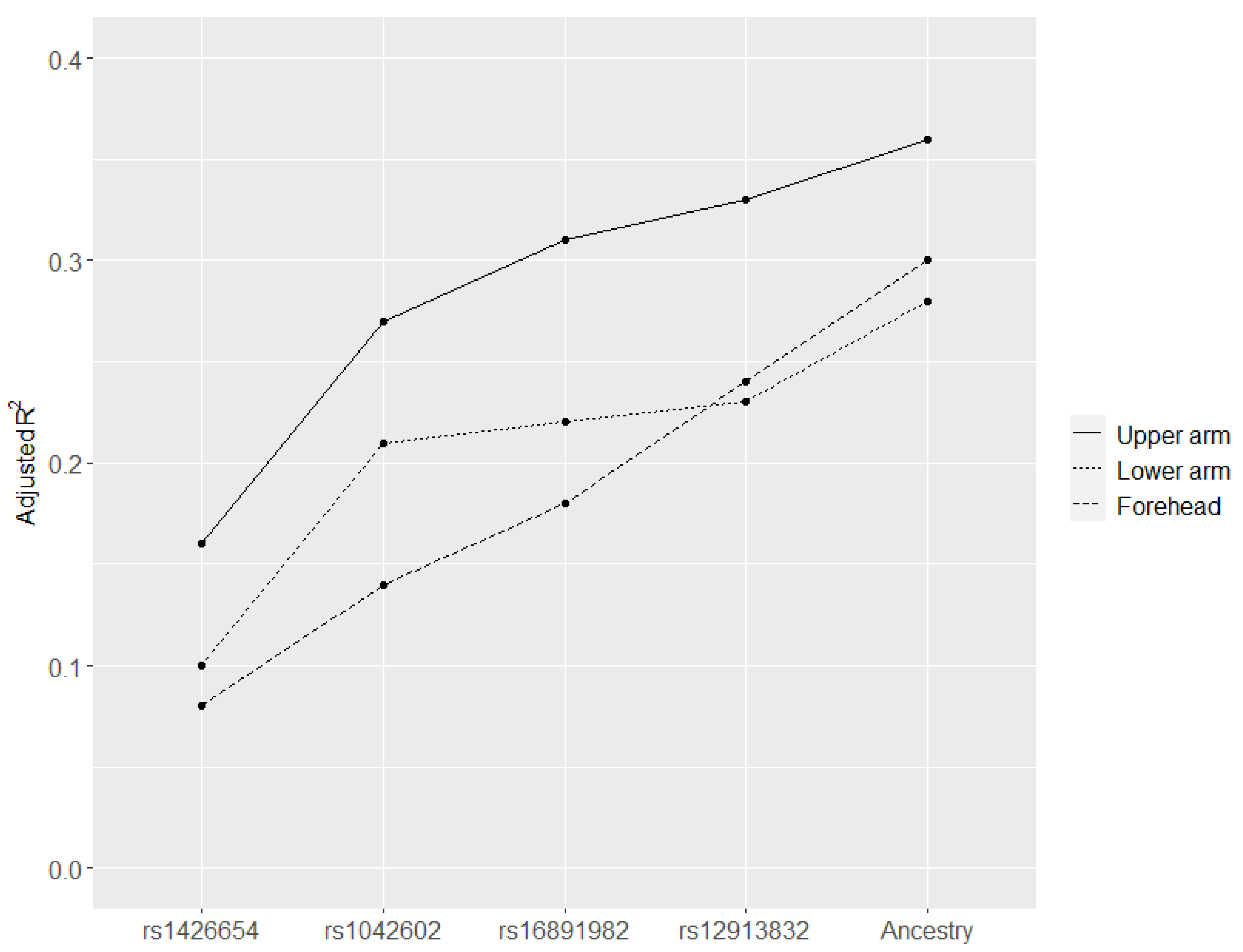
3.7. Correlation of Skin Pigmentation with SNPs from Existing Skin Colour Models
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kayser, M.; Schneider, P.M. DNA-based prediction of human externally visible characteristics in forensics: Motivations, scientific challenges, and ethical considerations. Forensic Sci. Int. Genet. 2009, 3, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Samuel, G.; Prainsack, B. Forensic DNA phenotyping in Europe: Views “on the ground” from those who have a professional stake in the technology. New Genet. Soc. 2019, 38, 119–141. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.D.; Meyer, O.S.; Simão, F.; Jannuzzi, J.; Carvalho, E.; Andersen, M.M.; Pereira, V.; Børsting, C.; Morling, N.; Gusmão, L. Skin pigmentation and genetic variants in an admixed Brazilian population of primarily European ancestry. Int. J. Legal Med. 2020, 134, 1569–1579. [Google Scholar] [CrossRef]
- Kayser, M.; De Knijff, P. Improving human forensics through advances in genetics, genomics and molecular biology. Nat. Rev. Genet. 2011, 12, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Maroñas, O.; Phillips, C.; Söchtig, J.; Gomez-Tato, A.; Cruz, R.; Alvarez-Dios, J.; de Cal, M.C.; Ruiz, Y.; Fondevila, M.; Carracedo, Á.; et al. Development of a forensic skin colour predictive test. Forensic Sci. Int. Genet. 2014, 13, 34–44. [Google Scholar] [CrossRef]
- Parra, E.J.; Kittles, R.A.; Shriver, M.D. Implications of correlations between skin color and genetic ancestry for biomedical research. Nat. Genet. 2004, 36, S54–S60. [Google Scholar] [CrossRef] [PubMed]
- Stokowski, R.P.; Pant, P.V.K.; Dadd, T.; Fereday, A.; Hinds, D.A.; Jarman, C.; Filsell, W.; Ginger, R.S.; Green, M.R.; Van Der Ouderaa, F.J.; et al. A genomewide association study of skin pigmentation in a South Asian population. Am. J. Hum. Genet. 2007, 81, 1119–1132. [Google Scholar] [CrossRef] [Green Version]
- Shriver, M.D.; Parra, E.J.; Dios, S.; Bonilla, C.; Norton, H.; Jovel, C.; Pfaff, C.; Jones, C.; Massac, A.; Cameron, N.; et al. Skin pigmentation, biogeographical ancestry and admixture mapping. Hum. Genet. 2003, 112, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, R.K.; Henderson, M.S.; Walsh, M.H.; Garrison, N.; Kelch, J.T.; Cohen-Barak, O.; Erickson, D.T.; John Meaney, F.; Bruce Walsh, J.; Cheng, K.C. Predicting phenotype from genotype: Normal pigmentation. J. Forensic Sci. 2010, 55, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Beleza, S.; Johnson, N.A.; Candille, S.I.; Absher, D.M.; Coram, M.A.; Lopes, J.; Campos, J.; Araújo, I.I.; Anderson, T.M.; Vilhjálmsson, B.J.; et al. Genetic Architecture of Skin and Eye Color in an African-European Admixed Population. PLOS Genet. 2013, 9, e1003372. [Google Scholar] [CrossRef] [Green Version]
- Lona-Durazo, F.; Hernandez-Pacheco, N.; Fan, S.; Zhang, T.; Choi, J.; Kovacs, M.A.; Loftus, S.K.; Le, P.; Edwards, M.; Fortes-Lima, C.A.; et al. Meta-analysis of GWA studies provides new insights on the genetic architecture of skin pigmentation in recently admixed populations. BMC Genet. 2019, 20, 59. [Google Scholar] [CrossRef] [Green Version]
- Wright, R.P. The Ancient Indus: Urbanism, Economy, and Society; Case Studies in Early Societies; Cambridge University Press: Cambridge, UK, 2010; ISBN 9780521576529. [Google Scholar]
- Wynbrandt, J. A Brief History of Pakistan; Brief history; Infobase Publishing: New York, NY, USA, 2009; ISBN 9780816061846. [Google Scholar]
- Pereira, V.; Mogensen, H.S.; Børsting, C.; Morling, N. Evaluation of the Precision ID Ancestry Panel for crime case work: A SNP typing assay developed for typing of 165 ancestral informative markers. Forensic Sci. Int. Genet. 2017, 28, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, H.S.; Tvedebrink, T.; Børsting, C.; Pereira, V.; Morling, N. Ancestry prediction efficiency of the software GenoGeographer using a z-score method and the ancestry informative markers in the Precision ID Ancestry Panel. Forensic Sci. Int. Genet. 2020, 44, 102154. [Google Scholar] [CrossRef] [PubMed]
- Al-Asfi, M.; McNevin, D.; Mehta, B.; Power, D.; Gahan, M.E.; Daniel, R. Assessment of the Precision ID Ancestry panel. Int. J. Legal Med. 2018, 132, 1581–1594. [Google Scholar] [CrossRef] [PubMed]
- Meyer, O.S.; Andersen, J.D.; Børsting, C. Presentation of the Human Pigmentation (HuPi) AmpliSeqTM custom panel. Forensic Sci. Int. Genet. Suppl. Ser. 2019, 7, 478–479. [Google Scholar] [CrossRef]
- Walsh, S.; Chaitanya, L.; Breslin, K.; Muralidharan, C.; Bronikowska, A.; Pospiech, E.; Koller, J.; Kovatsi, L.; Wollstein, A.; Branicki, W.; et al. Global skin colour prediction from DNA. Hum. Genet. 2017, 136, 847–863. [Google Scholar] [CrossRef] [Green Version]
- Chaitanya, L.; Breslin, K.; Zuñiga, S.; Wirken, L.; Pośpiech, E.; Kukla-Bartoszek, M.; Sijen, T.; de Knijff, P.; Liu, F.; Branicki, W.; et al. The HIrisPlex-S system for eye, hair and skin colour prediction from DNA: Introduction and forensic developmental validation. Forensic Sci. Int. Genet. 2018, 35, 123–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kongshoj, B.; Thorleifsson, A.; Wulf, H.C. Pheomelanin and eumelanin in human skin determined by high-performance liquid chromatography and its relation to in vivo reflectance measurements. Photodermatol. Photoimmunol. Photomed. 2006, 22, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Ravnbak, M.H. Objective determination of Fitzpatrick skin type. Dan. Med. Bull. 2010, 57, B4153. [Google Scholar]
- Farzad, M.S.; Pedersen, B.M.; Mogensen, H.S.; Børsting, C. Development of an automated AmpliSeqTM library building workflow for biological stain samples on the Biomek(®) 3000. Biotechniques 2020, 68, 342–344. [Google Scholar] [CrossRef] [Green Version]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, N.A. DISTRUCT: A program for the graphical display of population structure. Mol. Ecol. Notes 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Simayijiang, H.; Børsting, C.; Tvedebrink, T.; Morling, N. Analysis of Uyghur and Kazakh populations using the Precision ID Ancestry Panel. Forensic Sci. Int. Genet. 2019, 43, 102144. [Google Scholar] [CrossRef] [PubMed]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S; Springer: New York, NY, USA, 2002. [Google Scholar]
- Shan, M.A.; Børsting, C.; Morling, N. Forensic application and genetic diversity of 21 autosomal STR loci in five major population groups of Pakistan. Int. J. Legal Med. 2021, 135, 775–777. [Google Scholar] [CrossRef]
- Korn, A.; Nourzaei, M. “Those were the hungry years”: A glimpse of Coastal Afro-Balochi. J. R. Asiat. Soc. 2018, 28, 661–695. [Google Scholar] [CrossRef] [Green Version]
- Siddiqi, M.H.; Akhtar, T.; Rakha, A.; Abbas, G.; Ali, A.; Haider, N.; Ali, A.; Hayat, S.; Masooma, S.; Ahmad, J.; et al. Genetic characterization of the Makrani people of Pakistan from mitochondrial DNA control-region data. Leg. Med. 2015, 17, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Mallick, C.B.; Iliescu, F.M.; Möls, M.; Hill, S.; Tamang, R.; Chaubey, G.; Goto, R.; Ho, S.Y.W.; Gallego Romero, I.; Crivellaro, F.; et al. The light skin allele of SLC24A5 in South Asians and Europeans shares identity by descent. PLoS Genet. 2013, 9, e1003912. [Google Scholar] [CrossRef] [Green Version]
- Jonnalagadda, M.; Faizan, M.A.; Ozarkar, S.; Ashma, R.; Kulkarni, S.; Norton, H.L.; Parra, E. A Genome-Wide Association Study of Skin and Iris Pigmentation among Individuals of South Asian Ancestry. Genome Biol. Evol. 2019, 11, 1066–1076. [Google Scholar] [CrossRef] [Green Version]
- Lamason, R.L.; Mohideen, M.-A.P.K.; Mest, J.R.; Wong, A.C.; Norton, H.L.; Aros, M.C.; Jurynec, M.J.; Mao, X.; Humphreville, V.R.; Humbert, J.E.; et al. SLC24A5, a Putative Cation Exchanger, Affects Pigmentation in Zebrafish and Humans. Science 2005, 310, 1782–1786. [Google Scholar] [CrossRef] [Green Version]
- Nan, H.; Kraft, P.; Qureshi, A.A.; Guo, Q.; Chen, C.; Hankinson, S.E.; Hu, F.B.; Thomas, G.; Hoover, R.N.; Chanock, S.; et al. Genome-wide association study of tanning phenotype in a population of European ancestry. J. Investig. Dermatol. 2009, 129, 2250–2257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, A.L.; Chen, W.; Thurber, A.E.; Smit, D.J.; Smith, A.G.; Bladen, T.G.; Brown, D.L.; Duffy, D.L.; Pastorino, L.; Bianchi-Scarra, G. Analysis of cultured human melanocytes based on polymorphisms within the SLC45A2/MATP, SLC24A5/NCKX5, and OCA2/P loci. J. Investig. Dermatol. 2009, 129, 392–405. [Google Scholar] [CrossRef] [Green Version]
- Chaki, M.; Sengupta, M.; Mondal, M.; Bhattacharya, A.; Mallick, S. Molecular and functional studies of tyrosinase variants among Indian oculocutaneous albinism type 1 patients. J. Investig. Dermatol. 2011, 131, 260–262. [Google Scholar] [CrossRef]
- Jagirdar, K.; Smit, D.J.; Ainger, S.A.; Lee, K.J.; Brown, D.L.; Chapman, B.; Zhen Zhao, Z.; Montgomery, G.W.; Martin, N.G.; Stow, J.L. Molecular analysis of common polymorphisms within the human Tyrosinase locus and genetic association with pigmentation traits. Pigment Cell Melanoma Res. 2014, 27, 552–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eiberg, H.; Troelsen, J.; Nielsen, M.; Mikkelsen, A.; Mengel-From, J.; Kjaer, K.W.; Hansen, L. Blue eye color in humans may be caused by a perfectly associated founder mutation in a regulatory element located within the HERC2 gene inhibiting OCA2 expression. Hum. Genet. 2008, 123, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Sturm, R.A.; Duffy, D.L.; Zhao, Z.Z.; Leite, F.P.N.; Stark, M.S.; Hayward, N.K.; Martin, N.G.; Montgomery, G.W. A single SNP in an evolutionary conserved region within intron 86 of the HERC2 gene determines human blue-brown eye color. Am. J. Hum. Genet. 2008, 82, 424–431. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; van Duijn, K.; Vingerling, J.R.; Hofman, A.; Uitterlinden, A.G.; Janssens, A.C.J.W.; Kayser, M. Eye color and the prediction of complex phenotypes from genotypes. Curr. Biol. 2009, 19, R192–R193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visser, M.; Kayser, M.; Palstra, R.-J. HERC2 rs12913832 modulates human pigmentation by attenuating chromatin-loop formation between a long-range enhancer and the OCA2 promoter. Genome Res. 2012, 22, 446–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavan, W.J.; Sturm, R.A. The genetics of human skin and hair pigmentation. Annu. Rev. Genomics Hum. Genet. 2019, 20, 41–72. [Google Scholar] [CrossRef]
- Kosoy, R.; Nassir, R.; Tian, C.; White, P.A.; Butler, L.M.; Silva, G.; Kittles, R.; Alarcon-Riquelme, M.E.; Gregersen, P.K.; Belmont, J.W.; et al. Ancestry informative marker sets for determining continental origin and admixture proportions in common populations in America. Hum. Mutat. 2009, 30, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidd, K.K.; Speed, W.C.; Pakstis, A.J.; Furtado, M.R.; Fang, R.; Madbouly, A.; Maiers, M.; Middha, M.; Friedlaender, F.R.; Kidd, J.R. Progress toward an efficient panel of SNPs for ancestry inference. Forensic Sci. Int. Genet. 2014, 10, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Phillips, C.; Parson, W.; Lundsberg, B.; Santos, C.; Freire-Aradas, A.; Torres, M.; Eduardoff, M.; Børsting, C.; Johansen, P.; Fondevila, M.; et al. Building a forensic ancestry panel from the ground up: The EUROFORGEN Global AIM-SNP set. Forensic Sci. Int. Genet. 2014, 11, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Phillips, C.; McNevin, D.; Kidd, K.K.; Lagacé, R.; Wootton, S.; de la Puente, M.; Freire-Aradas, A.; Mosquera-Miguel, A.; Eduardoff, M.; Gross, T.; et al. MAPlex—A massively parallel sequencing ancestry analysis multiplex for Asia-Pacific populations. Forensic Sci. Int. Genet. 2019, 42, 213–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Upper Arm | Lower Arm | Forehead | |||||
|---|---|---|---|---|---|---|---|
| Gene | SNP | Adjusted R2 | Sign.1 | Adjusted R2 | Sign. 1 | Adjusted R2 | Sign. 1 |
| TYR | rs1042602 | 0.13 | *** | 0.11 | ** | 0.071 | ** |
| GRM5 | rs10831496 | 0.029 | – | 0.047 | * | 0.014 | – |
| HERC2 | rs12913832 | 0.025 | – | 0.016 | – | 0.064 | * |
| SLC24A5 | rs1426654 | 0.16 | *** | 0.10 | ** | 0.082 | ** |
| SLC45A2 | rs16891982 | 0.075 | ** | 0.028 | – | 0.058 | ** |
| Upper Arm | Lower Arm | Forehead | ||||
|---|---|---|---|---|---|---|
| Adjusted R2 | Sign. 1 | Adjusted R2 | Sign. 1 | Adjusted R2 | Sign. 1 | |
| Five SNPs 2 | 0.33 | *** | 0.24 | *** | 0.24 | *** |
| Best model 3 | 0.33 | *** | 0.23 | *** | 0.24 | *** |
| Best model + ancestry 4 | 0.37 | *** | 0.28 | *** | 0.30 | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shan, M.A.; Meyer, O.S.; Refn, M.; Morling, N.; Andersen, J.D.; Børsting, C. Analysis of Skin Pigmentation and Genetic Ancestry in Three Subpopulations from Pakistan: Punjabi, Pashtun, and Baloch. Genes 2021, 12, 733. https://doi.org/10.3390/genes12050733
Shan MA, Meyer OS, Refn M, Morling N, Andersen JD, Børsting C. Analysis of Skin Pigmentation and Genetic Ancestry in Three Subpopulations from Pakistan: Punjabi, Pashtun, and Baloch. Genes. 2021; 12(5):733. https://doi.org/10.3390/genes12050733
Chicago/Turabian StyleShan, Muhammad Adnan, Olivia Strunge Meyer, Mie Refn, Niels Morling, Jeppe Dyrberg Andersen, and Claus Børsting. 2021. "Analysis of Skin Pigmentation and Genetic Ancestry in Three Subpopulations from Pakistan: Punjabi, Pashtun, and Baloch" Genes 12, no. 5: 733. https://doi.org/10.3390/genes12050733
APA StyleShan, M. A., Meyer, O. S., Refn, M., Morling, N., Andersen, J. D., & Børsting, C. (2021). Analysis of Skin Pigmentation and Genetic Ancestry in Three Subpopulations from Pakistan: Punjabi, Pashtun, and Baloch. Genes, 12(5), 733. https://doi.org/10.3390/genes12050733