
Multi-Scale Organization of the Drosophila melanogaster Genome


Abstract
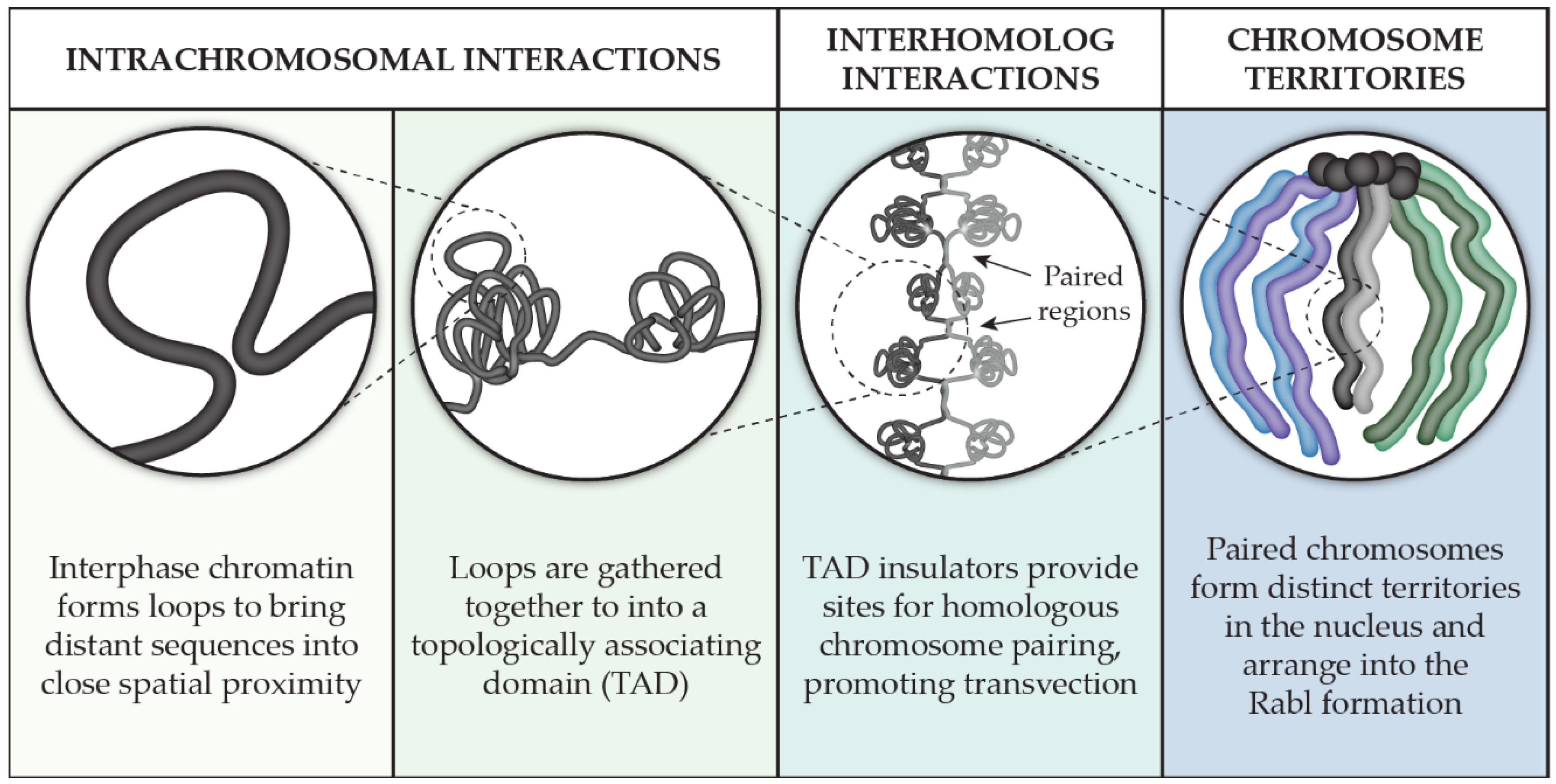
:1. Introduction
2. TADs in Drosophila
2.1. Sub-Chromosomal Organization and the Advent of TADs
2.2. Chromatin Context of TADs in D. melanogaster
2.3. Insulator Proteins Involved in TAD Formation, Regulation, and Maintenance

{kind=link}
{kind=link}
| Drosophila Gene Name | Alternative Name(s) | DNA Binding Activity | Interactions with Other Architectural Proteins |
|---|---|---|---|
| Boundary element-associated factor of 32kD (BEAF-32) | BEAF-32A/BEAF-32B (isoforms of BEAF-32), BEAF, boundary element-associated factor | Yes via a BED finger domain [79,80] | Chromator [81] CP190 [81] |
| Centrosomal protein 190kD (Cp190) | Rb188, DMAP190, Cen190, CP-190, Centrosomal Protein 190, CP190 | Yes, via zinc finger domains [39] | BEAF-32B [81] Chromator [81] CLAMP [42] dCTCF [82] HIPP1 [63] Mod(mdg4) [39] Pita [51] Su(Hw) [39] ZIPIC [51] |
| Chromatin-linked adaptor for MSL proteins (Clamp) | CLAMP | Yes, via zinc finger domains [83] | CP190) [42] Mod(mdg4) [42] Su(Hw) [42] |
| Chromator (Chro) | Chriz, Chro(Chriz) | None found | BEAF-32B [81] CP190 [81] dCTCF [84] JIL-1 kinase [77] Mod(mdg4) [85] |
| CTCF | dCTCF, CCCTC-binding factor | Yes, via zinc finger domains [34] | Chromator [84] CP190 [82] E(y)2 [86] HIPP1 [63] |
| deformed wings (dwg) | Zw5, l(1)zw5, zw-5, EG:95B7.6 , zeste-white 5 | Yes, via zinc finger domains [50] | (itself) [87] |
| enhancer of yellow 2 (E(y)2) | ENY2, late boundary complex, LBC | None found | dCTCF [86] GAF [45] Su(Hw) [61] |
| HP1 and insulator partner protein 1 (HIPP1) | N/A | None found | CP190 [63] dCTCF [63] Pita [63] Mod(mdg4) [63] Su(Hw) [63] |
| modifier of mdg4 (Mod(mdg4)) | E(var)3-93D, doom, mnm, Mod(mdg4)2.2, Mod(mdg4)-67.2 | Not directly [43] | Chromator [85] CLAMP [42] CP190 [39] GAF [44] HIPP1 [63] Su(Hw) [56,88] |
| Motif 1 Binding Protein (M1BP) | N/A | Yes, via zinc finger domains [47] | CP190 [49] Mod(mdg4) [49] Su(Hw) [49] |
| pita | Spdk, spotted dick | Yes, via zinc finger domains [89] | CP190 [51] HIPP1 [63] ZIPIC [51] |
| suppressor of Hairy wing (su(Hw)) | suHw | Yes, via zinc finger domains [90] | CLAMP [42] CP190 [39] E(y)2 [61] HIPP1 [63] Mod(mdg4) [56,88] |
| Trithorax-like (Trl) | GAF, GAGA, GAGA factor, Nc70F, GAGA-factor | Yes, via a zinc finger domain [91] | E(y)2 [45] Mod(mdg4) [45] |
| zelda (zld) | vfl, vielfaltig, EP134 | Yes, via zinc finger domains [92] | Ubx [93] |
| Zinc-finger protein interacting with CP190 (ZIPIC) | N/A | Yes, via zinc finger domains [94] | CP190 [51] Pita [51] |
3. Interhomolog Interactions
3.1. Homologous Chromosome Pairing in Somatic Tissue
3.2. Transvection and Interhomolog Communication
4. Chromosome Territories and the Rabl Configuration
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References and Note
- Flemming, W. Contributions to the Knowledge of the Cell and Its Vital Processes. (Translation of “Beitrage zur Kenntniss der Zelle und Ihrer Lebenserscheinungen, Theil II” originally published in 1880 in Archivfiir Mikroskopische Anatomie, Volume 18, pages 151–259). J. Cell Biol. 1965, 25, 3–69. [Google Scholar] [CrossRef]
- Szabo, Q.; Bantignies, F.; Cavalli, G. Principles of genome folding into topologically associating domains. Sci. Adv. 2019, 5, eaaw1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dame, R.T.; Rashid, F.Z.M.; Grainger, D.C. Chromosome organization in bacteria: Mechanistic insights into genome structure and function. Nat. Rev. Genet. 2020, 21, 227–242. [Google Scholar] [CrossRef]
- Takemata, N.; Bell, S.D. Emerging views of genome organization in Archaea. J. Cell Sci. 2020, 133, jcs243782. [Google Scholar] [CrossRef]
- Csink, A.K.; Henikoff, S. Large-scale chromosomal movements during interphase progression in Drosophila. J. Cell Biol. 1998, 143, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Marshall, W.F.; Straight, A.; Marko, J.F.; Swedlow, J.; Dernburg, A.; Belmont, A.; Murray, A.W.; Agard, D.A.; Sedat, J.W. Interphase chromosomes undergo constrained diffusional motion in living cells. Curr. Biol. 1997, 7, 930–939. [Google Scholar] [CrossRef] [Green Version]
- Dekker, J.; Misteli, T. Long-range chromatin interactions. Cold Spring Harb. Perspect. Biol. 2015, 7, a019356. [Google Scholar] [CrossRef] [Green Version]
- Dekker, J.; Rippe, K.; Dekker, M.; Kleckner, N. Capturing chromosome conformation. Science 2002, 295, 1306–1311. [Google Scholar] [CrossRef] [Green Version]
- Lieberman-Aiden, E.; van Berkum, N.L.; Williams, L.; Imakaev, M.; Ragoczy, T.; Telling, A.; Amit, I.; Lajoie, B.R.; Sabo, P.J.; Dorschner, M.O.; et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 2009, 326, 289–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Z.; Andronescu, M.; Schutz, K.; McIlwain, S.; Kim, Y.J.; Lee, C.; Shendure, J.; Fields, S.; Blau, C.A.; Noble, W.S. A three-dimensional model of the yeast genome. Nature 2010, 465, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Sexton, T.; Yaffe, E.; Kenigsberg, E.; Bantignies, F.; Leblanc, B.; Hoichman, M.; Parrinello, H.; Tanay, A.; Cavalli, G. Three-dimensional folding and functional organization principles of the Drosophila genome. Cell 2012, 148, 458–472. [Google Scholar] [CrossRef] [Green Version]
- Nora, E.P.; Lajoie, B.R.; Schulz, E.G.; Giorgetti, L.; Okamoto, I.; Servant, N.; Piolot, T.; van Berkum, N.L.; Meisig, J.; Sedat, J.; et al. Spatial partitioning of the regulatory landscape of the X-inactivation centre. Nature 2012, 485, 381–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, J.R.; Selvaraj, S.; Yue, F.; Kim, A.; Li, Y.; Shen, Y.; Hu, M.; Liu, J.S.; Ren, B. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature 2012, 485, 376–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempfer, R.; Pombo, A. Methods for mapping 3D chromosome architecture. Nat. Rev. Genet. 2020, 21, 207–226. [Google Scholar] [CrossRef] [PubMed]
- McKay, D.J.; Stutzman, A.V.; Dowen, J.M. Advancements in mapping 3D genome architecture. Methods 2020, 170, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Cardozo Gizzi, A.M.; Cattoni, D.I.; Fiche, J.-B.; Espinola, S.M.; Gurgo, J.; Messina, O.; Houbron, C.; Ogiyama, Y.; Papadopoulos, G.L.; Cavalli, G.; et al. Microscopy-Based Chromosome Conformation Capture Enables Simultaneous Visualization of Genome Organization and Transcription in Intact Organisms. Mol. Cell 2019, 74, 212–222.e5. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.-H.S.; Weiner, A.; Lajoie, B.; Dekker, J.; Friedman, N.; Rando, O.J. Mapping Nucleosome Resolution Chromosome Folding in Yeast by Micro-C. Cell 2015, 162, 108–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Sun, Q.; Czajkowsky, D.M.; Shao, Z. Sub-kb Hi-C in D. melanogaster reveals conserved characteristics of TADs between insect and mammalian cells. Nat. Commun. 2018, 9, 188. [Google Scholar] [CrossRef] [Green Version]
- Matthews, N.E.; White, R. Chromatin Architecture in the Fly: Living without CTCF/Cohesin Loop Extrusion?: Alternating Chromatin States Provide a Basis for Domain Architecture in Drosophila. BioEssays 2019, 41, e1900048. [Google Scholar] [CrossRef] [Green Version]
- Luzhin, A.V.; Flyamer, I.M.; Khrameeva, E.E.; Ulianov, S.V.; Razin, S.V.; Gavrilov, A.A. Quantitative differences in TAD border strength underly the TAD hierarchy in Drosophila chromosomes. J. Cell. Biochem. 2019, 120, 4494–4503. [Google Scholar] [CrossRef]
- Stadler, M.R.; Haines, J.E.; Eisen, M.B. Convergence of topological domain boundaries, insulators, and polytene interbands revealed by high-resolution mapping of chromatin contacts in the early Drosophila melanogaster embryo. Elife 2017, 6, e29550. [Google Scholar] [CrossRef] [Green Version]
- Ulianov, S.V.; Zakharova, V.V.; Galitsyna, A.A.; Kos, P.I.; Polovnikov, K.E.; Flyamer, I.M.; Mikhaleva, E.A.; Khrameeva, E.E.; Germini, D.; Logacheva, M.D.; et al. Order and stochasticity in the folding of individual Drosophila genomes. Nat. Commun. 2021, 12, 41. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Perez-Rathke, A.; Czajkowsky, D.M.; Shao, Z.; Liang, J. High-resolution single-cell 3D-models of chromatin ensembles during Drosophila embryogenesis. Nat. Commun. 2021, 12, 205. [Google Scholar] [CrossRef] [PubMed]
- Fudenberg, G.; Imakaev, M.; Lu, C.; Goloborodko, A.; Abdennur, N.; Mirny, L.A. Formation of Chromosomal Domains by Loop Extrusion. Cell Rep. 2016, 15, 2038–2049. [Google Scholar] [CrossRef] [Green Version]
- Jost, D.; Carrivain, P.; Cavalli, G.; Vaillant, C. Modeling epigenome folding: Formation and dynamics of topologically associated chromatin domains. Nucleic Acids Res. 2014, 42, 9553–9561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulianov, S.V.; Doronin, S.A.; Khrameeva, E.E.; Kos, P.I.; Luzhin, A.V.; Starikov, S.S.; Galitsyna, A.A.; Nenasheva, V.V.; Ilyin, A.A.; Flyamer, I.M.; et al. Nuclear lamina integrity is required for proper spatial organization of chromatin in Drosophila. Nat. Commun. 2019, 10, 1176. [Google Scholar] [CrossRef] [Green Version]
- Ou, H.D.; Phan, S.; Deerinck, T.J.; Thor, A.; Ellisman, M.H.; O’Shea, C.C. ChromEMT: Visualizing 3D chromatin structure and compaction in interphase and mitotic cells. Science 2017, 357, eaag0025. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.K.; Jost, D. How epigenome drives chromatin folding and dynamics, insights from efficient coarse-grained models of chromosomes. PLoS Comput. Biol. 2018, 14, e1006159. [Google Scholar] [CrossRef] [Green Version]
- Ing-Simmons, E.; Vaid, R.; Bing, X.Y.; Levine, M.; Mannervik, M.; Vaquerizas, J.M. Independence of chromatin conformation and gene regulation during Drosophila dorsoventral patterning. Nat. Genet. 2021, 53, 487–499. [Google Scholar] [CrossRef]
- Espinola, S.M.; Götz, M.; Bellec, M.; Messina, O.; Fiche, J.B.; Houbron, C.; Dejean, M.; Reim, I.; Cardozo Gizzi, A.M.; Lagha, M.; et al. Cis-regulatory chromatin loops arise before TADs and gene activation, and are independent of cell fate during early Drosophila development. Nat. Genet. 2021, 53, 477–486. [Google Scholar] [CrossRef]
- Ulianov, S.V.; Khrameeva, E.E.; Gavrilov, A.A.; Flyamer, I.M.; Kos, P.; Mikhaleva, E.A.; Penin, A.A.; Logacheva, M.D.; Imakaev, M.V.; Chertovich, A.; et al. Active chromatin and transcription play a key role in chromosome partitioning into topologically associating domains. Genome Res. 2016, 26, 70–84. [Google Scholar] [CrossRef] [Green Version]
- El-Sharnouby, S.; Fischer, B.; Magbanua, J.P.; Umans, B.; Flower, R.; Choo, S.W.; Russell, S.; White, R. Regions of very low H3K27me3 partition the Drosophila genome into topological domains. PLoS ONE 2017, 12, e0172725. [Google Scholar] [CrossRef] [Green Version]
- Rowley, M.J.; Nichols, M.H.; Lyu, X.; Ando-Kuri, M.; Rivera, I.S.M.; Hermetz, K.; Wang, P.; Ruan, Y.; Corces, V.G. Evolutionarily Conserved Principles Predict 3D Chromatin Organization. Mol. Cell 2017, 67, 837–852.e7. [Google Scholar] [CrossRef] [Green Version]
- Moon, H.; Filippova, G.; Loukinov, D.; Pugacheva, E.; Chen, Q.; Smith, S.T.; Munhall, A.; Grewe, B.; Bartkuhn, M.; Arnold, R.; et al. CTCF is conserved from Drosophila to humans and confers enhancer blocking of the Fab-8 insulator. EMBO Rep. 2005, 6, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyrchanova, O.; Maksimenko, O.; Ibragimov, A.; Sokolov, V.; Postika, N.; Lukyanova, M.; Schedl, P.; Georgiev, P. The insulator functions of the Drosophila polydactyl C2H2 zinc finger protein CTCF: Necessity versus sufficiency. Sci. Adv. 2020, 6, eaaz3152. [Google Scholar] [CrossRef] [Green Version]
- Kaushal, A.; Mohana, G.; Dorier, J.; Özdemir, I.; Omer, A.; Cousin, P.; Semenova, A.; Taschner, M.; Dergai, O.; Marzetta, F.; et al. CTCF loss has limited effects on global genome architecture in Drosophila despite critical regulatory functions. Nat. Commun. 2021, 12, 1101. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Hart, C.M.; Laemmli, U.K. Visualization of chromosomal domains with boundary element-associated factor BEAF-32. Cell 1995, 81, 879–889. [Google Scholar] [CrossRef] [Green Version]
- Van Bortle, K.; Ramos, E.; Takenaka, N.; Yang, J.; Wahi, J.E.; Corces, V.G. Drosophila CTCF tandemly aligns with other insulator proteins at the borders of H3K27me3 domains. Genome Res. 2012, 22, 2176–2187. [Google Scholar] [CrossRef] [Green Version]
- Pai, C.-Y.; Lei, E.P.; Ghosh, D.; Corces, V.G. The centrosomal protein CP190 is a component of the gypsy chromatin insulator. Mol. Cell 2004, 16, 737–748. [Google Scholar] [CrossRef]
- Ahanger, S.H.; Günther, K.; Weth, O.; Bartkuhn, M.; Bhonde, R.R.; Shouche, Y.S.; Renkawitz, R. Ectopically tethered CP190 induces large-scale chromatin decondensation. Sci. Rep. 2014, 4, 3917. [Google Scholar] [CrossRef] [Green Version]
- Bartkuhn, M.; Straub, T.; Herold, M.; Herrmann, M.; Rathke, C.; Saumweber, H.; Gilfillan, G.D.; Becker, P.B.; Renkawitz, R. Active promoters and insulators are marked by the centrosomal protein 190. EMBO J. 2009, 28, 877–888. [Google Scholar] [CrossRef] [Green Version]
- Bag, I.; Dale, R.K.; Palmer, C.; Lei, E.P. The zinc-finger protein CLAMP promotes gypsy chromatin insulator function in Drosophila. J. Cell Sci. 2019, 132, jcs226092. [Google Scholar] [CrossRef] [Green Version]
- Gause, M.; Morcillo, P.; Dorsett, D. Insulation of enhancer-promoter communication by a gypsy transposon insert in the Drosophila cut gene: Cooperation between suppressor of hairy-wing and modifier of mdg4 proteins. Mol. Cell. Biol. 2001, 21, 4807–4817. [Google Scholar] [CrossRef] [Green Version]
- Melnikova, L.; Juge, F.; Gruzdeva, N.; Mazur, A.; Cavalli, G.; Georgiev, P. Interaction between the GAGA factor and Mod(mdg4) proteins promotes insulator bypass in Drosophila. Proc. Natl. Acad. Sci. USA 2004, 101, 14806–14811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomaev, D.; Mikhailova, A.; Erokhin, M.; Shaposhnikov, A.V.; Moresco, J.J.; Blokhina, T.; Wolle, D.; Aoki, T.; Ryabykh, V.; Yates, J.R.; et al. The GAGA factor regulatory network: Identification of GAGA factor associated proteins. PLoS ONE 2017, 12, e0173602. [Google Scholar] [CrossRef] [PubMed]
- Gaskill, M.M.; Gibson, T.J.; Larson, E.D.; Harrison, M.M. GAF is essential for zygotic genome activation and chromatin accessibility in the early Drosophila embryo. eLife 2021, 10. [Google Scholar] [CrossRef]
- Li, J.; Gilmour, D.S. Distinct mechanisms of transcriptional pausing orchestrated by GAGA factor and M1BP, a novel transcription factor. EMBO J. 2013, 32, 1829–1841. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, F.; Bhardwaj, V.; Arrigoni, L.; Lam, K.C.; Grüning, B.A.; Villaveces, J.; Habermann, B.; Akhtar, A.; Manke, T. High-resolution TADs reveal DNA sequences underlying genome organization in flies. Nat. Commun. 2018, 9, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bag, I.; Chen, S.; Rosin, L.F.; Chen, Y.; Liu, C.-Y.; Yu, G.-Y.; Lei, E.P. M1BP cooperates with CP190 to activate transcription at TAD borders and promote chromatin insulator activity. bioRxiv 2020. [Google Scholar] [CrossRef]
- Gaszner, M.; Vazquez, J.; Schedl, P. The Zw5 protein, a component of the scs chromatin domain boundary, is able to block enhancer-promoter interaction. Genes Dev. 1999, 13, 2098–2107. [Google Scholar] [CrossRef] [Green Version]
- Maksimenko, O.; Bartkuhn, M.; Stakhov, V.; Herold, M.; Zolotarev, N.; Jox, T.; Buxa, M.K.; Kirsch, R.; Bonchuk, A.; Fedotova, A.; et al. Two new insulator proteins, Pita and ZIPIC, target CP190 to chromatin. Genome Res. 2015, 25, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nègre, N.; Brown, C.D.; Shah, P.K.; Kheradpour, P.; Morrison, C.A.; Henikoff, J.G.; Feng, X.; Ahmad, K.; Russell, S.; White, R.A.H.; et al. A comprehensive map of insulator elements for the Drosophila genome. PLoS Genet. 2010, 6, e1000814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, N.; Emberly, E.; Cuvier, O.; Hart, C.M. Genome-wide mapping of boundary element-associated factor (BEAF) binding sites in Drosophila melanogaster links BEAF to transcription. Mol. Cell. Biol. 2009, 29, 3556–3568. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Ramos, E.; Corces, V.G. The BEAF-32 insulator coordinates genome organization and function during the evolution of Drosophila species. Genome Res. 2012, 22, 2199–2207. [Google Scholar] [CrossRef] [Green Version]
- Vorobyeva, N.E.; Mazina, M.U.; Golovnin, A.K.; Kopytova, D.V.; Gurskiy, D.Y.; Nabirochkina, E.N.; Georgieva, S.G.; Georgiev, P.G.; Krasnov, A.N. Insulator protein Su(Hw) recruits SAGA and Brahma complexes and constitutes part of Origin Recognition Complex-binding sites in the Drosophila genome. Nucleic Acids Res. 2013, 41, 5717–5730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgiev, P.; Kozycina, M. Interaction between mutations in the suppressor of Hairy wing and modifier of mdg4 genes of Drosophila melanogaster affecting the phenotype of gypsy-induced mutations. Genetics 1996, 142, 425–436. [Google Scholar] [CrossRef]
- Geyer, P.K.; Corces, V.G. DNA position-specific repression of transcription by a Drosophila zinc finger protein. Genes Dev. 1992, 6, 1865–1873. [Google Scholar] [CrossRef] [Green Version]
- Melnikova, L.; Kostyuchenko, M.; Molodina, V.; Parshikov, A.; Georgiev, P.; Golovnin, A. Multiple interactions are involved in a highly specific association of the Mod(mdg4)-67.2 isoform with the Su(Hw) sites in Drosophila. Open Biol. 2017, 7, 170150. [Google Scholar] [CrossRef] [Green Version]
- Melnikova, L.; Kostyuchenko, M.; Parshikov, A.; Georgiev, P.; Golovnin, A. Role of Su(Hw) zinc finger 10 and interaction with CP190 and Mod(mdg4) proteins in recruiting the Su(Hw) complex to chromatin sites in Drosophila. PLoS ONE 2018, 13, e0193497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyrchanova, O.; Maksimenko, O.; Stakhov, V.; Ivlieva, T.; Parshikov, A.; Studitsky, V.M.; Georgiev, P. Effective blocking of the white enhancer requires cooperation between two main mechanisms suggested for the insulator function. PLoS Genet. 2013, 9, e1003606. [Google Scholar] [CrossRef] [Green Version]
- Kurshakova, M.; Maksimenko, O.; Golovnin, A.; Pulina, M.; Georgieva, S.; Georgiev, P.; Krasnov, A. Evolutionarily conserved E(y)2/Sus1 protein is essential for the barrier activity of Su(Hw)-dependent insulators in Drosophila. Mol. Cell 2007, 27, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Parnell, T.J.; Viering, M.M.; Skjesol, A.; Helou, C.; Kuhn, E.J.; Geyer, P.K. An endogenous suppressor of hairy-wing insulator separates regulatory domains in Drosophila. Proc. Natl. Acad. Sci. USA 2003, 100, 13436–13441. [Google Scholar] [CrossRef] [Green Version]
- Alekseyenko, A.A.; Gorchakov, A.A.; Zee, B.M.; Fuchs, S.M.; Kharchenko, P.V.; Kuroda, M.I. Heterochromatin-associated interactions of Drosophila HP1a with dADD1, HIPP1, and repetitive RNAs. Genes Dev. 2014, 28, 1445–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerasimova, T.I.; Byrd, K.; Corces, V.G. A chromatin insulator determines the nuclear localization of DNA. Mol. Cell 2000, 6, 1025–1035. [Google Scholar] [CrossRef]
- Byrd, K.; Corces, V.G. Visualization of chromatin domains created by the gypsy insulator of Drosophila. J. Cell Biol. 2003, 162, 565–574. [Google Scholar] [CrossRef]
- Kaye, E.G.; Kurbidaeva, A.; Wolle, D.; Aoki, T.; Schedl, P.; Larschan, E. Drosophila Dosage Compensation Loci Associate with a Boundary-Forming Insulator Complex. Mol. Cell. Biol. 2017, 37, e00253-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolle, D.; Cleard, F.; Aoki, T.; Deshpande, G.; Schedl, P.; Karch, F. Functional Requirements for Fab-7 Boundary Activity in the Bithorax Complex. Mol. Cell. Biol. 2015, 35, 3739–3752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanzuolo, C.; Roure, V.; Dekker, J.; Bantignies, F.; Orlando, V. Polycomb response elements mediate the formation of chromosome higher-order structures in the bithorax complex. Nat. Cell Biol. 2007, 9, 1167–1174. [Google Scholar] [CrossRef]
- Mihaly, J.; Hogga, I.; Gausz, J.; Gyurkovics, H.; Karch, F. In situ dissection of the Fab-7 region of the bithorax complex into a chromatin domain boundary and a Polycomb-response element. Development 1997, 124, 1809–1820. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Barolo, S.; Szymanski, P.; Levine, M. The Fab-7 element of the bithorax complex attenuates enhancer-promoter interactions in the Drosophila embryo. Genes Dev. 1996, 10, 3195–3201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, T.; Sarkeshik, A.; Yates, J.; Schedl, P. Elba, a novel developmentally regulated chromatin boundary factor is a hetero-tripartite DNA binding complex. eLife 2012, 1, e00171. [Google Scholar] [CrossRef]
- Gambetta, M.C.; Furlong, E.E.M. The Insulator Protein CTCF Is Required for Correct Hox Gene Expression, but Not for Embryonic Development in Drosophila. Genetics 2018, 210, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franke, A.; Baker, B.S. The rox1 and rox2 RNAs are essential components of the compensasome, which mediates dosage compensation in Drosophila. Mol. Cell 1999, 4, 117–122. [Google Scholar] [CrossRef]
- Ramírez, F.; Lingg, T.; Toscano, S.; Lam, K.C.; Georgiev, P.; Chung, H.-R.; Lajoie, B.R.; de Wit, E.; Zhan, Y.; de Laat, W.; et al. High-Affinity Sites Form an Interaction Network to Facilitate Spreading of the MSL Complex across the X Chromosome in Drosophila. Mol. Cell 2015, 60, 146–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, J.J.; Ilik, I.A.; Qu, K.; Georgiev, P.; Chu, C.; Akhtar, A.; Chang, H.Y. Revealing long noncoding RNA architecture and functions using domain-specific chromatin isolation by RNA purification. Nat. Biotechnol. 2014, 32, 933–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendjan, S.; Taipale, M.; Kind, J.; Holz, H.; Gebhardt, P.; Schelder, M.; Vermeulen, M.; Buscaino, A.; Duncan, K.; Mueller, J.; et al. Nuclear pore components are involved in the transcriptional regulation of dosage compensation in Drosophila. Mol. Cell 2006, 21, 811–823. [Google Scholar] [CrossRef]
- Rath, U.; Ding, Y.; Deng, H.; Qi, H.; Bao, X.; Zhang, W.; Girton, J.; Johansen, J.; Johansen, K.M. The chromodomain protein, Chromator, interacts with JIL-1 kinase and regulates the structure of Drosophila polytene chromosomes. J. Cell Sci. 2006, 119, 2332–2341. [Google Scholar] [CrossRef] [Green Version]
- Larkin, A.; Marygold, S.J.; Antonazzo, G.; Attrill, H.; Dos Santos, G.; Garapati, P.V.; Goodman, J.L.; Gramates, L.S.; Millburn, G.; Strelets, V.B.; et al. FlyBase: Updates to the Drosophila melanogaster knowledge base. Nucleic Acids Res. 2021, 49, D899–D907. [Google Scholar] [CrossRef] [PubMed]
- Aravind, L. The BED finger, a novel DNA-binding domain in chromatin-boundary-element-binding proteins and transposases. Trends Biochem. Sci. 2000, 25, 421–423. [Google Scholar] [CrossRef]
- Hart, C.M.; Zhao, K.; Laemmli, U.K. The scs’ boundary element: Characterization of boundary element-associated factors. Mol. Cell. Biol. 1997, 17, 999–1009. [Google Scholar] [CrossRef] [Green Version]
- Vogelmann, J.; Le Gall, A.; Dejardin, S.; Allemand, F.; Gamot, A.; Labesse, G.; Cuvier, O.; Nègre, N.; Cohen-Gonsaud, M.; Margeat, E.; et al. Chromatin insulator factors involved in long-range DNA interactions and their role in the folding of the Drosophila genome. PLoS Genet. 2014, 10, e1004544. [Google Scholar] [CrossRef] [PubMed]
- Mohan, M.; Bartkuhn, M.; Herold, M.; Philippen, A.; Heinl, N.; Bardenhagen, I.; Leers, J.; White, R.A.H.; Renkawitz-Pohl, R.; Saumweber, H.; et al. The Drosophila insulator proteins CTCF and CP190 link enhancer blocking to body patterning. EMBO J. 2007, 26, 4203–4214. [Google Scholar] [CrossRef] [Green Version]
- Soruco, M.M.L.; Chery, J.; Bishop, E.P.; Siggers, T.; Tolstorukov, M.Y.; Leydon, A.R.; Sugden, A.U.; Goebel, K.; Feng, J.; Xia, P.; et al. The CLAMP protein links the MSL complex to the X chromosome during Drosophila dosage compensation. Genes Dev. 2013, 27, 1551–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohla, D.; Herold, M.; Panzer, I.; Buxa, M.K.; Ali, T.; Demmers, J.; Krüger, M.; Scharfe, M.; Jarek, M.; Bartkuhn, M.; et al. A functional insulator screen identifies NURF and dREAM components to be required for enhancer-blocking. PLoS ONE 2014, 9, e107765. [Google Scholar] [CrossRef] [Green Version]
- Golovnin, A.K.; Shapovalov, I.S.; Kostyuchenko, M.V.; Shamsutdinov, M.F.; Georgiev, P.G.; Melnikova, L.S. Chromator protein directly interacts with the common part of the Drosophila melanogaster Mod(mdg4) family proteins. Dokl. Biochem. Biophys. 2014, 454, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Maksimenko, O.; Kyrchanova, O.; Bonchuk, A.; Stakhov, V.; Parshikov, A.; Georgiev, P. Highly conserved ENY2/Sus1 protein binds to Drosophila CTCF and is required for barrier activity. Epigenetics 2014, 9, 1261–1270. [Google Scholar] [CrossRef] [Green Version]
- Zolotarev, N.A.; Maksimenko, O.G.; Georgiev, P.G.; Bonchuk, A.N. ZAD-domain is essential for nuclear localization of insulator proteins in Drosophila melanogaster. Acta Nat. 2016, 8, 97–102. [Google Scholar] [CrossRef]
- Ghosh, D.; Gerasimova, T.I.; Corces, V.G. Interactions between the Su(Hw) and Mod(mdg4) proteins required for gypsy insulator function. EMBO J. 2001, 20, 2518–2527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laundrie, B.; Peterson, J.S.; Baum, J.S.; Chang, J.C.; Fileppo, D.; Thompson, S.R.; McCall, K. Germline cell death is inhibited by P-element insertions disrupting the dcp-1/pita nested gene pair in Drosophila. Genetics 2003, 165, 1881–1888. [Google Scholar] [CrossRef]
- Parkhurst, S.M.; Harrison, D.A.; Remington, M.P.; Spana, C.; Kelley, R.L.; Coyne, R.S.; Corces, V.G. The Drosophila su(Hw) gene, which controls the phenotypic effect of the gypsy transposable element, encodes a putative DNA-binding protein. Genes Dev. 1988, 2, 1205–1215. [Google Scholar] [CrossRef] [Green Version]
- Soeller, W.C.; Oh, C.E.; Kornberg, T.B. Isolation of cDNAs encoding the Drosophila GAGA transcription factor. Mol. Cell. Biol. 1993, 13, 7961–7970. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.-L.; Nien, C.-Y.; Liu, H.-Y.; Metzstein, M.M.; Kirov, N.; Rushlow, C. The zinc-finger protein Zelda is a key activator of the early zygotic genome in Drosophila. Nature 2008, 456, 400–403. [Google Scholar] [CrossRef] [Green Version]
- Carnesecchi, J.; Sigismondo, G.; Domsch, K.; Baader, C.E.P.; Rafiee, M.-R.; Krijgsveld, J.; Lohmann, I. Multi-level and lineage-specific interactomes of the Hox transcription factor Ubx contribute to its functional specificity. Nat. Commun. 2020, 11, 1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krystel, J.; Ayyanathan, K. Global analysis of target genes of 21 members of the ZAD transcription factor family in Drosophila melanogaster. Gene 2013, 512, 373–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubin, T.; Macaisne, N.; Huynh, J.-R. Mixing and Matching Chromosomes during Female Meiosis. Cells 2020, 9, 696. [Google Scholar] [CrossRef] [Green Version]
- Stevens, N.M. A study of the germ cells of certain diptera, with reference to the heterochromosomes and the phenomena of synapsis. J. Exp. Zool. 1908, 5, 359–374. [Google Scholar] [CrossRef] [Green Version]
- Metz, C.W. Chromosome studies on the Diptera. II. The paired association of chromosomes in the Diptera, and its significance. J. Exp. Zool. 1916, 21, 213–279. [Google Scholar] [CrossRef] [Green Version]
- Halfer, C.; Barigozzi, C. Prophase synapsis in somatic cells of Drosophila melanogaster. Chromosom. Today 1973, 4, 181–186. [Google Scholar]
- Williams, B.R.; Bateman, J.R.; Novikov, N.D.; Wu, C.T. Disruption of topoisomerase II perturbs pairing in Drosophila cell culture. Genetics 2007, 177, 31–46. [Google Scholar] [CrossRef] [Green Version]
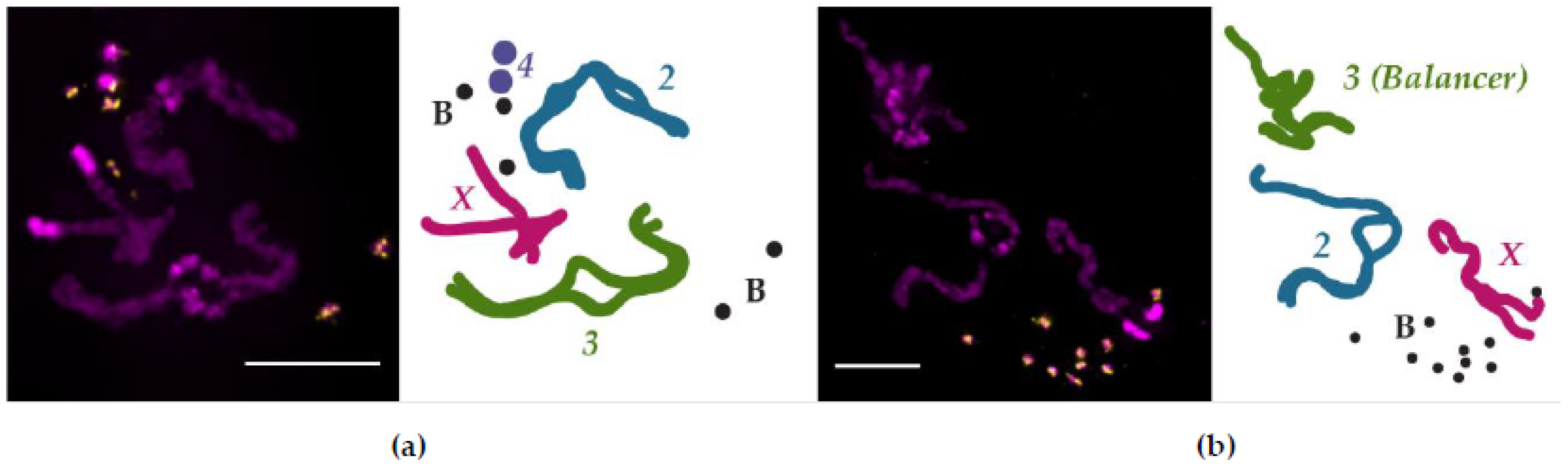
- Larval neuroblast preparations were performed as previously described in Hanlon et al., 2018 [101].
- Hanlon, S.L.; Miller, D.E.; Eche, S.; Hawley, R.S. Origin, composition, and structure of the supernumerary B chromosome of Drosophila melanogaster. Genetics 2018, 210, 1197–1212. [Google Scholar] [CrossRef] [Green Version]
- Bauerly, E.; Hughes, S.E.; Vietti, D.R.; Miller, D.E.; McDowell, W.; Scott Hawley, R. Discovery of supernumerary B chromosomes in Drosophila melanogaster. Genetics 2014, 196, 1007–1016. [Google Scholar] [CrossRef] [Green Version]
- Hiraoka, Y.; Dernburg, A.F.; Parmelee, S.J.; Rykowski, M.C.; Agard, D.A.; Sedat, J.W. The onset of homologous chromosome pairing during Drosophila melanogaster embryogenesis. J. Cell Biol. 1993, 120, 591–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fung, J.C.; Marshall, W.F.; Dernburg, A.; Agard, D.A.; Sedat, J.W. Homologous Chromosome Pairing in Drosophila melanogaster Proceeds through Multiple Independent Initiations. J. Cell Biol. 1998, 141, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Gemkow, M.J.; Verveer, P.J.; Arndt-Jovin, D.J. Homologous association of the Bithorax-Complex during embryogenesis: Consequences for transvection in Drosophila melanogaster. Development 1998, 125, 4541–4552. [Google Scholar] [CrossRef] [PubMed]
- Farrell, J.A.; O’Farrell, P.H. From egg to gastrula: How the cell cycle is remodeled during the Drosophila mid-blastula transition. Annu. Rev. Genet. 2014, 48, 269–294. [Google Scholar] [CrossRef] [Green Version]
- Bateman, J.R.; Wu, C.T. A genomewide survey argues that every zygotic gene product is dispensable for the initiation of somatic homolog pairing in Drosophila. Genetics 2008, 180, 1329–1342. [Google Scholar] [CrossRef] [Green Version]
- Hug, C.B.; Grimaldi, A.G.; Kruse, K.; Vaquerizas, J.M. Chromatin Architecture Emerges during Zygotic Genome Activation Independent of Transcription. Cell 2017, 169, 216–228.e19. [Google Scholar] [CrossRef] [Green Version]
- Ogiyama, Y.; Schuettengruber, B.; Papadopoulos, G.L.; Chang, J.M.; Cavalli, G. Polycomb-Dependent Chromatin Looping Contributes to Gene Silencing during Drosophila Development. Mol. Cell 2018, 71, 73–88.e5. [Google Scholar] [CrossRef] [Green Version]
- Fritsch, C.; Ploeger, G.; Arndt-Jovin, D.J. Drosophila under the lens: Imaging from chromosomes to whole embryos. Chromosom. Res. 2006, 14, 451–464. [Google Scholar] [CrossRef]
- Ramos, E.; Torre, E.A.; Bushey, A.M.; Gurudatta, B.V.; Corces, V.G. DNA Topoisomerase II modulates insulator function in Drosophila. PLoS ONE 2011, 6, e16562. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T. Condensin-Based Chromosome Organization from Bacteria to Vertebrates. Cell 2016, 164, 847–857. [Google Scholar] [CrossRef] [Green Version]
- Hartl, T.A.; Smith, H.F.; Bosco, G. Chromosome alignment and transvection are antagonized by condensin II. Science 2008, 322, 1384–1387. [Google Scholar] [CrossRef]
- Buster, D.W.; Daniel, S.G.; Nguyen, H.Q.; Windler, S.L.; Skwarek, L.C.; Peterson, M.; Roberts, M.; Meserve, J.H.; Hart, T.; Klebba, J.E.; et al. SCFSlimb ubiquitin ligase suppresses condensin II-mediated nuclear reorganization by degrading Cap-H2. J. Cell Biol. 2013, 201, 49–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, C.R.; Hartl, T.A.; Bosco, G. Condensin II Promotes the Formation of Chromosome Territories by Inducing Axial Compaction of Polyploid Interphase Chromosomes. PLoS Genet. 2012, 8, e1002873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, H.A.; Klebba, J.E.; Kusch, T.; Rogers, G.C.; Bosco, G. Condensin II regulates interphase chromatin organization through the Mrg-binding motif of Cap-H2. G3 Genes Genomes Genet. 2015, 5, 803–817. [Google Scholar] [CrossRef] [Green Version]
- Bateman, J.R.; Larschan, E.; D’Souza, R.; Marshall, L.S.; Dempsey, K.E.; Johnson, J.E.; Mellone, B.G.; Kuroda, M.I. A genome-wide screen identifies genes that affect somatic homolog pairing in Drosophila. G3 Genes Genomes Genet. 2012, 2, 731–740. [Google Scholar] [CrossRef] [Green Version]
- Joyce, E.F.; Williams, B.R.; Xie, T.; Wu, C.-T. Identification of genes that promote or antagonize somatic homolog pairing using a high-throughput FISH-based screen. PLoS Genet. 2012, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erceg, J.; AlHaj Abed, J.; Goloborodko, A.; Lajoie, B.R.; Fudenberg, G.; Abdennur, N.; Imakaev, M.; McCole, R.B.; Nguyen, S.C.; Saylor, W.; et al. The genome-wide multi-layered architecture of chromosome pairing in early Drosophila embryos. Nat. Commun. 2019, 10, 4486. [Google Scholar] [CrossRef] [Green Version]
- AlHaj Abed, J.; Erceg, J.; Goloborodko, A.; Nguyen, S.C.; McCole, R.B.; Saylor, W.; Fudenberg, G.; Lajoie, B.R.; Dekker, J.; Mirny, L.A.; et al. Highly structured homolog pairing reflects functional organization of the Drosophila genome. Nat. Commun. 2019, 10, 4485. [Google Scholar] [CrossRef] [Green Version]
- Viets, K.; Sauria, M.E.G.; Chernoff, C.; Rodriguez Viales, R.; Echterling, M.; Anderson, C.; Tran, S.; Dove, A.; Goyal, R.; Voortman, L.; et al. Characterization of Button Loci that Promote Homologous Chromosome Pairing and Cell-Type-Specific Interchromosomal Gene Regulation. Dev. Cell 2019, 51, 341–356.e7. [Google Scholar] [CrossRef]
- Ghavi-Helm, Y.; Jankowski, A.; Meiers, S.; Viales, R.R.; Korbel, J.O.; Furlong, E.E.M. Highly rearranged chromosomes reveal uncoupling between genome topology and gene expression. Nat. Genet. 2019, 51, 1272–1282. [Google Scholar] [CrossRef] [PubMed]
- Schoborg, T.; Kuruganti, S.; Rickels, R.; Labrador, M. The Drosophila gypsy insulator supports transvection in the presence of the vestigial enhancer. PLoS ONE 2013, 8, e81331. [Google Scholar] [CrossRef] [Green Version]
- Lim, B.; Heist, T.; Levine, M.; Fukaya, T. Visualization of Transvection in Living Drosophila embryos. Mol. Cell 2018, 70, 287–296.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, E.B. The Theory and Application of a New Method of Detecting Chromosomal Rearrangements in Drosophila melanogaster. Am. Nat. 1954, 88, 225–239. [Google Scholar] [CrossRef]
- Geyer, P.K.; Green, M.M.; Corces, V.G. Tissue-specific transcriptional enhancers may act in trans on the gene located in the homologous chromosome: The molecular basis of transvection in Drosophila. EMBO J. 1990, 9, 2247–2256. [Google Scholar] [CrossRef]
- Morris, J.R.; Chen, J.; Filandrinos, S.T.; Dunn, R.C.; Fisk, R.; Geyer, P.K.; Wu, C. An analysis of transvection at the yellow locus of Drosophila melanogaster. Genetics 1999, 151, 633–651. [Google Scholar] [CrossRef]
- Gelbart, W.M. Synapsis-dependent allelic complementation at the decapentaplegic gene complex in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1982, 79, 2636–2640. [Google Scholar] [CrossRef] [Green Version]
- Locke, J.; Tartof, K.D. Molecular analysis of cubitus interruptus (ci) mutations suggests an explanation for the unusual ci position effects. Mol. Gen. Genet. 1994, 243, 234–243. [Google Scholar] [CrossRef]
- Henikoff, S.; Dreesen, T.D. Trans-inactivation of the Drosophila brown gene: Evidence for transcriptional repression and somatic pairing dependence. Proc. Natl. Acad. Sci. USA 1989, 86, 6704–6708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Condie, J.M.; Brower, D.L. Allelic interactions at the engrailed locus of Drosophila: Engrailed protein expression in imaginal discs. Dev. Biol. 1989, 135, 31–42. [Google Scholar] [CrossRef]
- Zimmerman, J.E.; Bui, Q.T.; Liu, H.; Bonini, N.M. Molecular genetic analysis of Drosophila eyes absent mutants reveals an eye enhancer element. Genetics 2000, 154, 237–246. [Google Scholar] [CrossRef]
- Gibson, J.B.; Reed, D.S.; Bartoszewski, S.; Wilks, A. V Structural changes in the promoter region mediate transvection at the sn-glycerol-3-phosphate dehydrogenase gene of Drosophila melanogaster. Biochem. Genet. 1999, 37, 301–315. [Google Scholar] [CrossRef]
- Lee, J.J.; von Kessler, D.P.; Parks, S.; Beachy, P.A. Secretion and localized transcription suggest a role in positional signaling for products of the segmentation gene hedgehog. Cell 1992, 71, 33–50. [Google Scholar] [CrossRef]
- Scholz, H.; Deatrick, J.; Klaes, A.; Klämbt, C. Genetic dissection of pointed, a Drosophila gene encoding two ETS-related proteins. Genetics 1993, 135, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Southworth, J.W.; Kennison, J.A. Transvection and silencing of the Scr homeotic gene of Drosophila melanogaster. Genetics 2002, 161, 733–746. [Google Scholar] [CrossRef]
- Ronshaugen, M.; Levine, M. Visualization of trans-homolog enhancer-promoter interactions at the Abd-B hox locus in the Drosophila embryo. Dev. Cell 2004, 7, 925–932. [Google Scholar] [CrossRef]
- Johnston, R.J.; Desplan, C. Interchromosomal communication coordinates intrinsically stochastic expression between alleles. Science 2014, 343, 661–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, J.A.; Bell, J.B.; Carroll, S.B. Control of Drosophila wing and haltere development by the nuclear vestigial gene product. Genes Dev. 1991, 5, 2481–2495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, P.; Bhat, S. Effect of zeste on white complementation. Basic Life Sci. 1980, 16, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Buratovich, M.A.; Phillips, R.G.; Whittle, J.R. Genetic relationships between the mutations spade and Sternopleural and the wingless gene in Drosophila development. Dev. Biol. 1997, 185, 244–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bantignies, F.; Grimaud, C.; Lavrov, S.; Gabut, M.; Cavalli, G. Inheritance of polycomb-dependent chromosomal interactions in Drosophila. Genes Dev. 2003, 17, 2406–2420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, J.; Müller, M.; Pirrotta, V.; Sedat, J.W. The Mcp element mediates stable long-range chromosome-chromosome interactions in Drosophila. Mol. Biol. Cell 2006, 17, 2158–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, T.D.; Johnson, J.E.; Bateman, J.R. Position Effects Influence Transvection in Drosophila melanogaster. Genetics 2019, 213, 1289–1299. [Google Scholar] [CrossRef] [PubMed]
- Goldsborough, A.S.; Kornberg, T.B. Reduction of transcription by homologue asynapsis in Drosophila imaginal discs. Nature 1996, 381, 807–810. [Google Scholar] [CrossRef]
- Chen, J.L.; Huisinga, K.L.; Viering, M.M.; Ou, S.A.; Wu, C.T.; Geyer, P.K. Enhancer action in trans is permitted throughout the Drosophila genome. Proc. Natl. Acad. Sci. USA 2002, 99, 3723–3728. [Google Scholar] [CrossRef] [Green Version]
- Ou, S.A.; Chang, E.; Lee, S.; So, K.; Wu, C.T.; Morris, J.R. Effects of chromosomal rearrangements on transvection at the yellow gene of Drosophila melanogaster. Genetics 2009, 183, 483–496. [Google Scholar] [CrossRef] [Green Version]
- Tian, K.; Henderson, R.E.; Parker, R.; Brown, A.; Johnson, J.E.; Bateman, J.R. Two modes of transvection at the eyes absent gene of Drosophila demonstrate plasticity in transcriptional regulatory interactions in cis and in trans. PLoS Genet. 2019, 15, e1008152. [Google Scholar] [CrossRef] [Green Version]
- Blick, A.J.; Mayer-Hirshfeld, I.; Malibiran, B.R.; Cooper, M.A.; Martino, P.A.; Johnson, J.E.; Bateman, J.R. The capacity to act in trans varies among Drosophila enhancers. Genetics 2016, 203, 203–218. [Google Scholar] [CrossRef] [Green Version]
- Kravchenko, E.; Savitskaya, E.; Kravchuk, O.; Parshikov, A.; Georgiev, P.; Savitsky, M. Pairing between gypsy insulators facilitates the enhancer action in trans throughout the Drosophila genome. Mol. Cell. Biol. 2005, 25, 9283–9291. [Google Scholar] [CrossRef] [Green Version]
- Piwko, P.; Vitsaki, I.; Livadaras, I.; Delidakis, C. The Role of Insulators in Transgene Transvection in Drosophila. Genetics 2019, 212, 489–508. [Google Scholar] [CrossRef]
- Cremer, T.; Cremer, M. Chromosome territories. Cold Spring Harb. Perspect. Biol. 2010, 2, a003889. [Google Scholar] [CrossRef] [Green Version]
- Rabl, C. Über Zellteilung. Morphol. Jahrb. 1885, 214–330. [Google Scholar]
- Boveri, T. Die Blastomerenkerne von Ascaris megalocephala und die Theorie der Chromosomenindividualität. Arch. Zellforsch. 1909, 3, 181–268. [Google Scholar]
- Rosin, L.F.; Nguyen, S.C.; Joyce, E.F. Condensin II drives large-scale folding and spatial partitioning of interphase chromosomes in Drosophila nuclei. PLoS Genet. 2018, 14, e1007393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Tjong, H.; Li, X.; Gong, K.; Zhou, X.J.; Chiolo, I.; Alber, F. The three-dimensional genome organization of Drosophila melanogaster through data integration. Genome Biol. 2017, 18, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosin, L.F.; Crocker, O.; Isenhart, R.L.; Nguyen, S.C.; Xu, Z.; Joyce, E.F. Chromosome territory formation attenuates the translocation potential of cells. eLife 2019, 8, e49553. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.G.; Ogiyama, Y.; Martins, N.M.C.; Beliveau, B.J.; Acevedo, D.; Wu, C.-T.; Cavalli, G.; Karpen, G.H. Pericentromeric heterochromatin is hierarchically organized and spatially contacts H3K9me2 islands in euchromatin. PLoS Genet. 2020, 16, e1008673. [Google Scholar] [CrossRef] [Green Version]
- Marshall, W.F.; Dernburg, A.F.; Harmon, B.; Agard, D.A.; Sedat, J.W. Specific interactions of chromatin with the nuclear envelope: Positional determination within the nucleus in Drosophila melanogaster. Mol. Biol. Cell 1996, 7, 825–842. [Google Scholar] [CrossRef] [Green Version]
- Khosraviani, N.; Ostrowski, L.A.; Mekhail, K. Roles for Non-coding RNAs in Spatial Genome Organization. Front. Cell Dev. Biol. 2019, 7, 336. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peterson, S.C.; Samuelson, K.B.; Hanlon, S.L. Multi-Scale Organization of the Drosophila melanogaster Genome. Genes 2021, 12, 817. https://doi.org/10.3390/genes12060817
Peterson SC, Samuelson KB, Hanlon SL. Multi-Scale Organization of the Drosophila melanogaster Genome. Genes. 2021; 12(6):817. https://doi.org/10.3390/genes12060817
Chicago/Turabian StylePeterson, Samantha C., Kaylah B. Samuelson, and Stacey L. Hanlon. 2021. "Multi-Scale Organization of the Drosophila melanogaster Genome" Genes 12, no. 6: 817. https://doi.org/10.3390/genes12060817
APA StylePeterson, S. C., Samuelson, K. B., & Hanlon, S. L. (2021). Multi-Scale Organization of the Drosophila melanogaster Genome. Genes, 12(6), 817. https://doi.org/10.3390/genes12060817