
A Systematic Review on the Therapeutic Potentiality of PD-L1-Inhibiting MicroRNAs for Triple-Negative Breast Cancer: Toward Single-Cell Sequencing-Guided Biomimetic Delivery

 ,
, 
 , , , ,
, , , ,  , and
, and 
Abstract
:1. Introduction
2. Methods
2.1. Search Strategy
2.2. Study Selection
2.3. Eligibility Criteria
2.4. Data Extraction
2.5. Evaluating the Potential Bias among the Included Studies
2.6. In Silico Investigation
3. Results
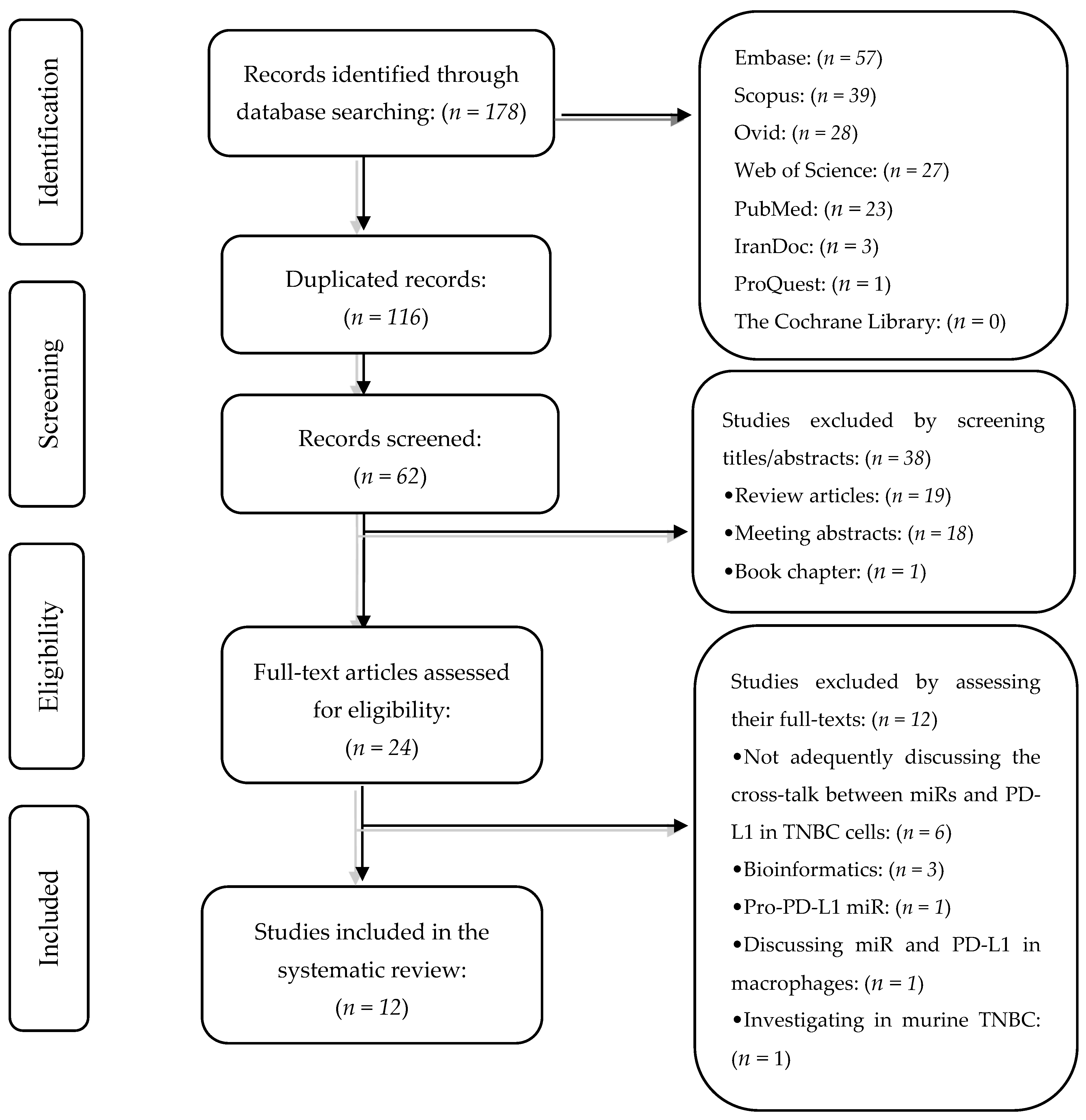
3.1. Selected Studies
3.2. Study Characteristic
3.3. The Evaluation of Potential Bias in the Included Studies
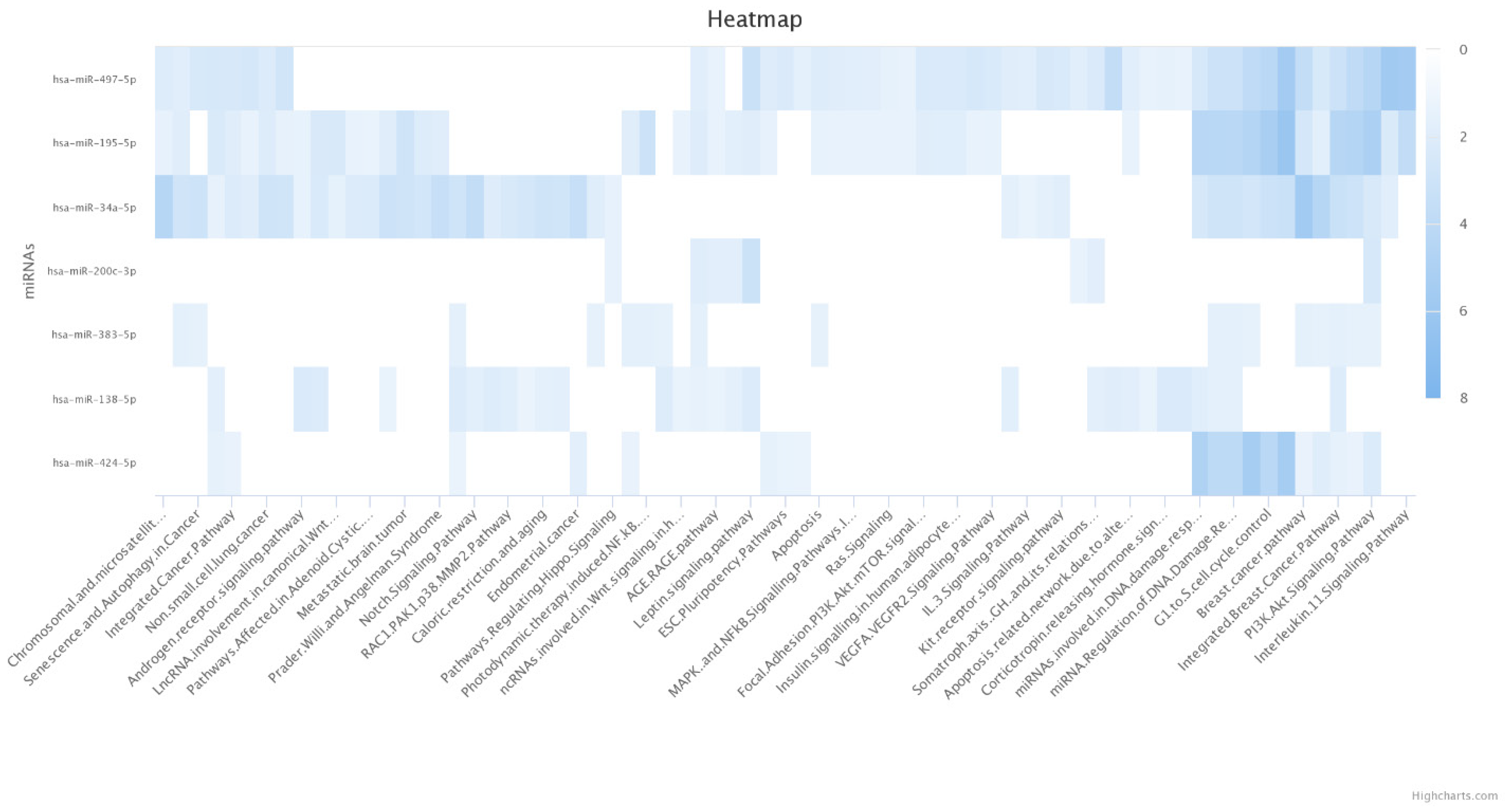
3.4. In Silico Investigation
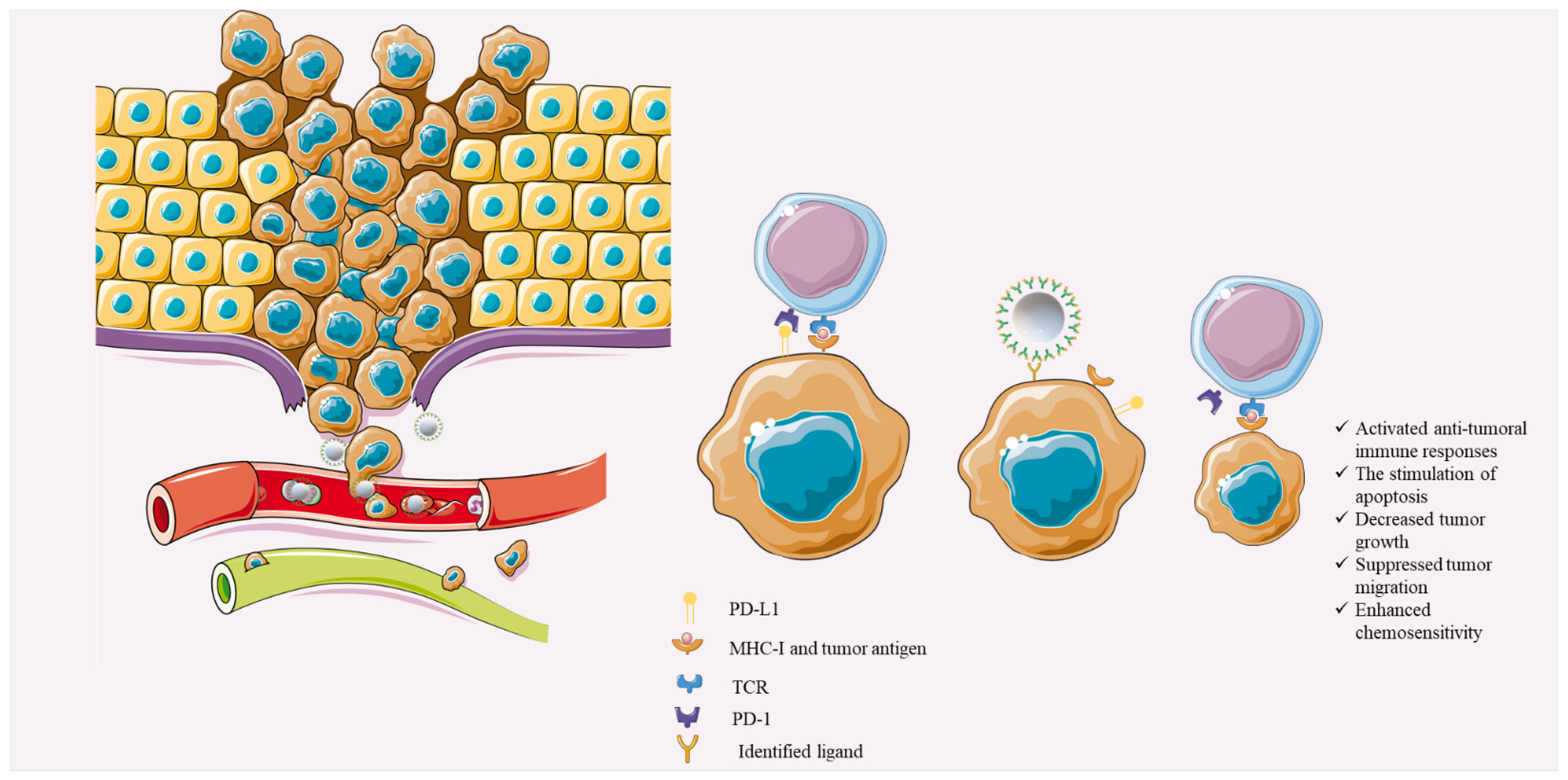
4. Discussion
4.1. miRs That Can Downregulate Tumoral PD-L1 Expression in TNBC Cells
4.1.1. miR-424-5p
4.1.2. miR-138-5p
4.1.3. miR-570-3p
4.1.4. miR-200c-3p
4.1.5. miR-383-5p
4.1.6. miR-34a-5p
4.1.7. miR-3609
4.1.8. miR-195-5p
4.1.9. miR-497-5p
4.2. The Delivery of miRs for Treating TNBC: Toward Effective, Biocompatible, and Safe Delivery
4.2.1. Biomimetic Carriers
4.2.2. Single-Cell Sequencing Technologies and Biomimetic Carriers: Future Perspectives
4.3. Some Considerations in miR-Based Therapy for Treating Cancer
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Yersal, O.; Barutca, S. Biological subtypes of breast cancer: Prognostic and therapeutic implications. World J. Clin. Oncol. 2014, 5, 412. [Google Scholar] [CrossRef]
- Penault-Llorca, F.; Viale, G. Pathological and molecular diagnosis of triple-negative breast cancer: A clinical perspective. Ann. Oncol. 2012, 23, vi19–vi22. [Google Scholar] [CrossRef] [PubMed]
- Pareja, F.; Geyer, F.C.; Marchiò, C.; Burke, K.A.; Weigelt, B.; Reis-Filho, J.S. Triple-negative breast cancer: The importance of molecular and histologic subtyping, and recognition of low-grade variants. NPJ Breast Cancer 2016, 2, 16036. [Google Scholar] [CrossRef] [PubMed]
- Hosseinkhani, N.; Derakhshani, A.; Kooshkaki, O.; Abdoli Shadbad, M.; Hajiasgharzadeh, K.; Baghbanzadeh, A.; Safarpour, H.; Mokhtarzadeh, A.; Brunetti, O.; Yue, S.C. Immune Checkpoints and CAR-T Cells: The Pioneers in Future Cancer Therapies? Int. J. Mol. Sci. 2020, 21, 8305. [Google Scholar] [CrossRef] [PubMed]
- Lotfinejad, P.; Asghari Jafarabadi, M.; Abdoli Shadbad, M.; Kazemi, T.; Pashazadeh, F.; Sandoghchian Shotorbani, S.; Jadidi Niaragh, F.; Baghbanzadeh, A.; Vahed, N.; Silvestris, N. Prognostic role and clinical significance of tumor-infiltrating lymphocyte (TIL) and programmed death ligand 1 (PD-L1) expression in triple-negative breast cancer (TNBC): A systematic review and meta-analysis study. Diagnostics 2020, 10, 704. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Xie, X.; Wang, H.; Xiao, X.; Yang, L.; Tian, Z.; Guo, X.; Zhang, L.; Tang, H.; Xie, X. PDL1 And LDHA act as ceRNAs in triple negative breast cancer by regulating miR-34a. J. Exp. Clin. Cancer Res. 2017, 36, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mediratta, K.; El-Sahli, S.; D’Costa, V.; Wang, L. Current progresses and challenges of immunotherapy in triple-negative breast cancer. Cancers 2020, 12, 3529. [Google Scholar] [CrossRef] [PubMed]
- Lotfinejad, P.; Kazemi, T.; Safaei, S.; Amini, M.; Baghbani, E.; Shotorbani, S.S.; Niaragh, F.J.; Derakhshani, A.; Shadbad, M.A.; Silvestris, N. PD-L1 silencing inhibits triple-negative breast cancer development and upregulates T-cell-induced pro-inflammatory cytokines. Biomed. Pharmacother. 2021, 138, 111436. [Google Scholar] [CrossRef]
- Cui, M.; Wang, H.; Yao, X.; Zhang, D.; Xie, Y.; Cui, R.; Zhang, X. Circulating microRNAs in cancer: Potential and challenge. Front. Genet. 2019, 10, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dastmalchi, N.; Safaralizadeh, R.; SM, B.K.; Sam, M.R.; Latifi-Navid, S.; Hussen, B.M.; Baradaran, B. An Updated Review of the Cross-talk between MicroRNAs and Epigenetic Factors in Cancers. Curr. Med. Chem. 2021. [CrossRef]
- Ahangar, N.K.; Hemmat, N.; Khalaj-Kondori, M.; Shadbad, M.A.; Sabaie, H.; Mokhtarzadeh, A.; Alizadeh, N.; Derakhshani, A.; Baghbanzadeh, A.; Dolatkhah, K. The Regulatory Cross-Talk between microRNAs and Novel Members of the B7 Family in Human Diseases: A Scoping Review. Int. J. Mol. Sci. 2021, 22, 2652. [Google Scholar] [CrossRef]
- Zhou, Y.; Yamamoto, Y.; Takeshita, F.; Yamamoto, T.; Xiao, Z.; Ochiya, T. Delivery of miR-424-5p via Extracellular vesicles promotes the apoptosis of MDA-MB-231 TNBC cells in the tumor microenvironment. Int. J. Mol. Sci. 2021, 22, 844. [Google Scholar] [CrossRef]
- Wang, L.L.; Huang, W.W.; Huang, J.; Huang, R.F.; Li, N.N.; Hong, Y.; Chen, M.L.; Wu, F.; Liu, J. Protective effect of hsa-miR-570-3p targeting CD274 on triple negative breast cancer by blocking PI3K/AKT/mTOR signaling pathway. Kaohsiung J. Med. Sci. 2020, 36, 581–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavez, K.J.; Garimella, S.V.; Lipkowitz, S. Triple negative breast cancer cell lines: One tool in the search for better treatment of triple negative breast cancer. Breast Dis. 2010, 32, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raj, A.T.; Kheur, S.; Bhonde, R.; Gupta, A.A.; Patil, S. Assessing the effect of human mesenchymal stem cell-derived conditioned media on human cancer cell lines: A systematic review. Tissue Cell 2021, 71, 101505. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.Q.; Su, P.; Yang, Y.H.; Yao, X.; Zhang, Y.Q.; Jin, F.; Yang, B.W. Identification of CTLA-4 associated with tumor microenvironment and competing interactions in triple negative breast cancer by co-expression network analysis. J. Cancer 2020, 11, 6365–6375. [Google Scholar] [CrossRef] [PubMed]
- Qattan, A.; Al-Tweigeri, T.; Alkhayal, W.; Suleman, K.; Tulbah, A.; Amer, S. Clinical Identification of Dysregulated Circulating microRNAs and Their Implication in Drug Response in Triple Negative Breast Cancer (TNBC) by Target Gene Network and Meta-Analysis. Genes 2021, 12, 549. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Mi, M.; Li, X.; Zheng, X.; Wu, G.; Zhang, L. lncRNA OSTN-AS1 May Represent a Novel Immune-Related Prognostic Marker for Triple-Negative Breast Cancer Based on Integrated Analysis of a ceRNA Network. Front. Genet. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naba, N.M.; Tolay, N.; Erman, B.; Sayi Yazgan, A. Doxorubicin inhibits miR-140 expression and upregulates PD-L1 expression in HCT116 cells, opposite to its effects on MDA-MB-231 cells. Turk. J. Biol. 2020, 44, 15–23. [Google Scholar] [CrossRef]
- Yang, Y.; Fang, T.; Cao, Y.L.; Lv, Y.X.; Chang, Q.Q.; Zhang, D.D. Ethyl Acetate Fraction from Hedyotis diffusa plus Scutellaria barbata Exerts Anti-Breast Cancer Effect via miR-200c-PDE7B/PD-L1-AKT/MAPK Axis. Evid.-Based Complement. Altern. Med. 2020, 2020, 3587095. [Google Scholar] [CrossRef]
- Noman, M.Z.; Janji, B.; Abdou, A.; Hasmim, M.; Terry, S.; Tan, T.Z.; Mami-Chouaib, F.; Thiery, J.P.; Chouaib, S. The immune checkpoint ligand PD-l1 is upregulated in EMT-activated human breast cancer cells by a mechanism involving ZEB-1 and miR-200. Oncoimmunology 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, N.; Song, P.; Fu, Y.; Ren, Y.; Li, Z.; Wang, J. LncRNA GATA3-AS1 facilitates tumour progression and immune escape in triple-negative breast cancer through destabilization of GATA3 but stabilization of PD-L1. Cell Prolif. 2020, 53. [Google Scholar] [CrossRef]
- Nafea, H.; Youness, R.A.; Abou-Aisha, K.; Gad, M.Z. LncRNA HEIH/miR-939-5p interplay modulates triple-negative breast cancer progression through NOS2-induced nitric oxide production. J. Cell. Physiol. 2021, 236, 5362–5372. [Google Scholar] [CrossRef] [PubMed]
- Youness, R.A.; Gad, A.Z.; Sanber, K.; Ahn, Y.J.; Lee, G.J.; Khallaf, E.; Hafez, H.M.; Motaal, A.A.; Ahmed, N.; Gad, M.Z. Targeting hydrogen sulphide signaling in breast cancer. J. Adv. Res. 2021, 27, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Hamed, M.M.; Handoussa, H.; Hussein, N.H.; Eissa, R.A.; Abdel-Aal, L.K.; El Tayebi, H.M. Oleuropin controls miR-194/XIST/PD-L1 loop in triple negative breast cancer: New role of nutri-epigenetics in immune-oncology. Life Sci. 2021, 277. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Tu, Y.; Xu, Y.; Guo, Y.; Yao, F.; Zhang, X. Endoplasmic reticulum stress-induced exosomal miR-27a-3p promotes immune escape in breast cancer via regulating PD-L1 expression in macrophages. J. Cell. Mol. Med. 2020, 24, 9560–9573. [Google Scholar] [CrossRef] [PubMed]
- Rogers, T.J.; Christenson, J.L.; Greene, L.I.; O’Neill, K.I.; Williams, M.M.; Gordon, M.A.; Nemkov, T.; D’Alessandro, A.; Degala, G.D.; Shin, J. Reversal of triple-negative breast cancer EMT by miR-200c decreases tryptophan catabolism and a program of immunosuppression. Mol. Cancer Res. 2019, 17, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Dastmalchi, N.; Safaralizadeh, R.; Hosseinpourfeizi, M.A.; Baradaran, B.; Khojasteh, S.M.B. MicroRNA-424-5p enhances chemosensitivity of breast cancer cells to Taxol and regulates cell cycle, apoptosis, and proliferation. Mol. Biol. Rep. 2021, 48, 1345–1357. [Google Scholar] [CrossRef] [PubMed]
- Soufiani, B.; Pourfathollah, A.A.; Arefian, E. Tumor Microenvironment Changing through Application of MicroRNA-34a Related Mesenchymal Stem Cells Conditioned Medium: Modulation of Breast Cancer Cells toward Non-aggressive Behavior. Iran. J. Allergy Asthma Immunol. 2021, 20, 221–232. [Google Scholar]
- Rasoolnezhad, M.; Safaralizadeh, R.; Hosseinpourfeizi, M.A.; Banan-Khojasteh, S.M.; Baradaran, B. MiRNA-138–5p: A strong tumor suppressor targeting PD-L-1 inhibits proliferation and motility of breast cancer cells and induces apoptosis. Eur. J. Pharmacol. 2021, 896, 173933. [Google Scholar] [CrossRef]
- Peng, F.; Xiong, L.; Peng, C. (-)-Sativan Inhibits Tumor Development and Regulates miR-200c/PD-L1 in Triple Negative Breast Cancer Cells. Front Pharmacol. 2020, 11, 251. [Google Scholar] [CrossRef] [PubMed]
- Dastmalchi, N.; Hosseinpourfeizi, M.A.; Khojasteh, S.M.B.; Baradaran, B.; Safaralizadeh, R. Tumor suppressive activity of miR-424-5p in breast cancer cells through targeting PD-L1 and modulating PTEN/PI3K/AKT/mTOR signaling pathway. Life Sci. 2020, 259, 118239. [Google Scholar] [CrossRef] [PubMed]
- Azarbarzin, S.; Hosseinpour-Feizi, M.A.; Banan Khojasteh, S.M.; Baradaran, B.; Safaralizadeh, R. MicroRNA -383-5p restrains the proliferation and migration of breast cancer cells and promotes apoptosis via inhibition of PD-L1. Life Sci. 2021, 267. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.Y.; Guo, Y.W.; Liu, C.P.; Huang, T. Prognostic value of PD-L1 for invasive breast cancer and its miR-34a-related mechanism of regulation. Int. J. Clin. Exp. Med. 2019, 12, 9984–9997. [Google Scholar]
- Li, D.; Wang, X.; Yang, M.; Kan, Q.; Duan, Z. miR3609 sensitizes breast cancer cells to adriamycin by blocking the programmed death-ligand 1 immune checkpoint. Exp. Cell Res. 2019, 380, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Cai, Y.; Zhang, D.; Sun, J.; Xu, C.; Zhao, W.; Jiang, W.; Pan, C. miR-195/miR-497 regulate CD274 expression of immune regulatory ligands in triple-negative breast cancer. J. Breast Cancer 2018, 21, 371. [Google Scholar] [CrossRef] [PubMed]
- FDA Grants Accelerated Approval to Pembrolizumab for Locally Recurrent Unresectable or Metastatic Triple Negative Breast Cancer. 2020. Available online: https://www.fda.gov/drugs/resources-information-approved-drugs/fda-grants-accelerated-approval-pembrolizumab-locally-recurrent-unresectable-or-metastatic-triple (accessed on 7 March 2021).
- Wang, J.; Wang, S.; Zhou, J.; Qian, Q. miR-424-5p regulates cell proliferation, migration and invasion by targeting doublecortin-like kinase 1 in basal-like breast cancer. Biomed. Pharmacother. 2018, 102, 147–152. [Google Scholar] [CrossRef]
- Xie, D.; Song, H.; Wu, T.; Li, D.; Hua, K.; Xu, H.; Zhao, B.; Wu, C.; Hu, J.; Ji, C. MicroRNA-424 serves an anti-oncogenic role by targeting cyclin-dependent kinase 1 in breast cancer cells. Oncol. Rep. 2018, 40, 3416–3426. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Ling, X.; Li, X.; Hou, X.; Zhao, D. MicroRNA-138-5p inhibits cell migration, invasion and EMT in breast cancer by directly targeting RHBDD1. Breast Cancer 2019, 26, 817–825. [Google Scholar] [CrossRef]
- Li, Y.; Yang, D.M.; Chen, J.; Li, Q.Q.; Yu, M.C. Inhibition of the proliferation and migration of triple negative breast cancer cells by targeting ezh2 using MIR-138-5P. Acta Med. Mediterr. 2021, 37, 1589–1595. [Google Scholar] [CrossRef]
- Li, D.; Song, H.; Wu, T.; Xie, D.; Hu, J.; Zhao, J.; Fang, L. MiR-138-5p targeting LIMK1 suppresses breast cancer cell proliferation and motility. RSC Adv. 2017, 7, 52030–52038. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Han, X.; Yu, K.; Sun, S.a.; Zhen, L.; Li, Z.; Wang, S. microRNA-200c downregulates XIAP expression to suppress proliferation and promote apoptosis of triple-negative breast cancer cells. Mol. Med. Rep. 2014, 10, 315–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Wang, F.; Xiang, Z.; Huang, T.; Zhou, W.B. LncRNA XIST promotes chemoresistance of breast cancer cells to doxorubicin by sponging miR-200c-3p to upregulate ANLN. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1464–1472. [Google Scholar] [CrossRef] [PubMed]
- Chang, B.-p.; Wang, D.-s.; Xing, J.-w.; Yang, S.-h.; Chu, Q.; Yu, S.-y. miR-200c inhibits metastasis of breast cancer cells by targeting HMGB1. J. Huazhong Univ. Sci. Technol. Med. Sci. 2014, 34, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Berber, U.; Yilmaz, I.; Narli, G.; Haholu, A.; Kucukodaci, Z.; Demirel, D. miR-205 and miR-200c: Predictive Micro RNAs for Lymph Node Metastasis in Triple Negative Breast Cancer. J. Breast Cancer 2014, 17, 143–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Kong, X.; Shi, Q.; Zhao, B. MicroRNA-383-5p acts as a potential prognostic biomarker and an inhibitor of tumor cell proliferation, migration, and invasion in breast cancer. Cancer Biomark. 2020, 27, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Xia, S.; Ma, M.; Zuo, Y. LINC00096 promotes the proliferation and invasion by sponging miR-383-5p and regulating RBM3 expression in triple-negative breast cancer. Onco Targets Ther. 2019, 12, 10569. [Google Scholar] [CrossRef] [Green Version]
- Si, W.; Li, Y.; Shao, H.; Hu, R.; Wang, W.; Zhang, K.; Yang, Q. MiR-34a inhibits breast cancer proliferation and progression by targeting Wnt1 in Wnt/β-catenin signaling pathway. Am. J. Med. Sci. 2016, 352, 191–199. [Google Scholar] [CrossRef]
- Haghi, M.; Taha, M.F.; Javeri, A. Suppressive effect of exogenous miR-16 and miR-34a on tumorigenesis of breast cancer cells. J. Cell. Biochem. 2019, 120, 13342–13353. [Google Scholar] [CrossRef]
- Derakhshani, A.; Silvestris, N.; Hemmat, N.; Asadzadeh, Z.; Abdoli Shadbad, M.; Nourbakhsh, N.S.; Mobasheri, L.; Vahedi, P.; Shahmirzaie, M.; Brunetti, O. Targeting TGF-β-Mediated SMAD Signaling pathway via novel recombinant cytotoxin II: A potent protein from naja naja oxiana venom in Melanoma. Molecules 2020, 25, 5148. [Google Scholar] [CrossRef] [PubMed]
- Abdoli Shadbad, M.; Hajiasgharzadeh, K.; Baradaran, B. Cross-talk between myeloid-derived suppressor cells and Mucin1 in breast cancer vaccination: On the verge of a breakthrough. Life Sci. 2020, 258, 118128. [Google Scholar] [CrossRef] [PubMed]
- Mansoori, B.; Silvestris, N.; Mohammadi, A.; Khaze, V.; Baghbani, E.; Mokhtarzadeh, A.; Shanehbandi, D.; Derakhshani, A.; Duijf, P.H.; Baradaran, B. miR-34a and miR-200c Have an Additive Tumor-Suppressive Effect on Breast Cancer Cells and Patient Prognosis. Genes 2021, 12, 267. [Google Scholar] [CrossRef]
- Vakhshiteh, F.; Khabazian, E.; Atyabi, F.; Ostad, S.N.; Madjd, Z.; Dinarvand, R. Peptide-conjugated liposomes for targeted miR-34a delivery to suppress breast cancer and cancer stem-like population. J. Drug Deliv. Sci. Technol. 2020, 57, 101687. [Google Scholar] [CrossRef]
- Vahidian, F.; Safarzadeh, E.; Mohammadi, A.; Najjary, S.; Mansoori, B.; Majidi, J.; Babaloo, Z.; Aghanejad, A.; Shadbad, M.A.; Mokhtarzadeh, A. siRNA-mediated silencing of CD44 delivered by Jet Pei enhanced Doxorubicin chemo sensitivity and altered miRNA expression in human breast cancer cell line (MDA-MB468). Mol. Biol. Rep. 2020, 47, 9541–9551. [Google Scholar] [CrossRef] [PubMed]
- Yaghobi, Z.; Movassaghpour, A.; Talebi, M.; Shadbad, M.A.; Hajiasgharzadeh, K.; Pourvahdani, S.; Baradaran, B. The role of CD44 in cancer chemoresistance: A concise review. Eur. J. Pharmacol. 2021, 174147. [Google Scholar] [CrossRef] [PubMed]
- Valcourt, D.M.; Day, E.S. Dual regulation of miR-34a and Notch signaling in triple-negative breast cancer by antibody/miRNA nanocarriers. Mol. Ther.-Nucleic Acids 2020, 21, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Cao, M.; Deng, X.; Xiao, X.; Yin, Z.; Hu, Q.; Zhou, Z.; Zhang, F.; Zhang, R.; Wu, Y. Degradable Hyaluronic Acid/Protamine Sulfate Interpolyelectrolyte Complexes as miRNA-Delivery Nanocapsules for Triple-Negative Breast Cancer Therapy. Adv. Healthc. Mater. 2015, 4, 281–290. [Google Scholar] [CrossRef]
- Fitzpatrick, C.; Bendek, M.F.; Briones, M.; Farfán, N.; Silva, V.A.; Nardocci, G.; Montecino, M.; Boland, A.; Deleuze, J.-F.; Villegas, J. Mitochondrial ncRNA targeting induces cell cycle arrest and tumor growth inhibition of MDA-MB-231 breast cancer cells through reduction of key cell cycle progression factors. Cell Death Dis. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Luo, Q.; Wei, C.; Li, X.; Li, J.; Chen, L.; Huang, Y.; Song, H.; Li, D.; Fang, L. MicroRNA-195-5p is a potential diagnostic and therapeutic target for breast cancer. Oncol. Rep. 2014, 31, 1096–1102. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, X.; Zou, C.; Kung, H.-F.; Lin, M.C.; Dress, A.; Wardle, F.; Jiang, B.-H.; Lai, L. miR-195 inhibits tumor growth and angiogenesis through modulating IRS1 in breast cancer. Biomed. Pharmacother. 2016, 80, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Jian, W.; Deng, X.-C.; Munankarmy, A.; Borkhuu, O.; Ji, C.-L.; Wang, X.-H.; Zheng, W.-F.; Yu, Y.-H.; Zhou, X.-Q.; Fang, L. KIF23 promotes triple negative breast cancer through activating epithelial-mesenchymal transition. Gland Surg. 2021, 10, 1941. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, Q.; Rui, Y.; Zhang, C.; Wang, W.; Gu, J.; Tang, J.; Ding, Y. HOXC13-AS promotes breast cancer cell growth through regulating miR-497-5p/PTEN axis. J. Cell. Physiol. 2019, 234, 22343–22351. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-Y.; Kuo, W.-H.; Hung, J.-H.; Lee, C.-Y.; Lee, Y.-H.; Chang, Y.-C.; Lin, W.-C.; Shen, C.-Y.; Huang, C.-S.; Hsieh, F.-J. Deregulated microRNAs in triple-negative breast cancer revealed by deep sequencing. Mol. Cancer 2015, 14, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, H.; Yang, J.; Zhang, B.; Wang, X.; Pei, L.; Zhang, L.; Lin, Z.; Wang, Y.; Wang, C. LncRNA GACAT3 predicts poor prognosis and promotes cell proliferation in breast cancer through regulation of miR-497/CCND2. Cancer Biomark. 2018, 22, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Zhao, W.; Li, W.; Ying, Z.; Jiang, D. Long noncoding RNA LINC00473 indicates a poor prognosis of breast cancer and accelerates tumor carcinogenesis by competing endogenous sponging miR-497. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3410–3420. [Google Scholar] [PubMed]
- Li, Y.; Hua, K.; Jin, J.; Fang, L. miR-497 inhibits proliferation and invasion in triple-negative breast cancer cells via YAP1. Oncol. Lett. 2021, 22, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.; Hossain, G.S.; Kocerha, J. The Potential for microRNA Therapeutics and Clinical Research. Front. Genet. 2019, 10, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forterre, A.; Komuro, H.; Aminova, S.; Harada, M. A comprehensive review of cancer MicroRNA therapeutic delivery strategies. Cancers 2020, 12, 1852. [Google Scholar] [CrossRef] [PubMed]
- Jha, A.; Nikam, A.N.; Kulkarni, S.; Mutalik, S.P.; Pandey, A.; Hegde, M.; Rao, B.S.S.; Mutalik, S. Biomimetic nanoarchitecturing: A disguised attack on cancer cells. J. Control. Release 2021, 329, 413–433. [Google Scholar] [CrossRef]
- Molinaro, R.; Martinez, J.O.; Zinger, A.; De Vita, A.; Storci, G.; Arrighetti, N.; De Rosa, E.; Hartman, K.A.; Basu, N.; Taghipour, N. Leukocyte-mimicking nanovesicles for effective doxorubicin delivery to treat breast cancer and melanoma. Biomater. Sci. 2020, 8, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.O.; Molinaro, R.; Hartman, K.A.; Boada, C.; Sukhovershin, R.; De Rosa, E.; Kirui, D.; Zhang, S.; Evangelopoulos, M.; Carter, A.M.; et al. Biomimetic nanoparticles with enhanced affinity towards activated endothelium as versatile tools for theranostic drug delivery. Theranostics 2018, 8, 1131–1145. [Google Scholar] [CrossRef]
- Wang, K.; Lei, Y.; Xia, D.; Xu, P.; Zhu, T.; Jiang, Z.; Ma, Y. Neutrophil membranes coated, antibiotic agent loaded nanoparticles targeting to the lung inflammation. Colloids Surf. B Biointerfaces 2020, 188, 110755. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.; Zhu, Q.; Wei, D.; Feng, J.; Yao, J.; Jiang, T.; Song, Q.; Wei, X.; Chen, H.; Gao, X. Nanoparticles coated with neutrophil membranes can effectively treat cancer metastasis. ACS Nano 2017, 11, 1397–1411. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Hu, Y.; Luo, S.; Wang, Y.; Gong, T.; Sun, X.; Fu, Y.; Zhang, Z. Neutrophil-mimicking therapeutic nanoparticles for targeted chemotherapy of pancreatic carcinoma. Acta Pharm. Sin. B 2019, 9, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Oroojalian, F.; Beygi, M.; Baradaran, B.; Mokhtarzadeh, A.; Shahbazi, M.A. Immune Cell Membrane-Coated Biomimetic Nanoparticles for Targeted Cancer Therapy. Small 2021, 17, 2006484. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Pan, H.; Li, W.; Chen, Z.; Ma, A.; Yin, T.; Liang, R.; Chen, F.; Ma, Y.; Jin, Y. T cell membrane mimicking nanoparticles with bioorthogonal targeting and immune recognition for enhanced photothermal therapy. Adv. Sci. 2019, 6, 1900251. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Hong, J.; Jung, M.; Kwon, S.P.; Song, S.Y.; Kim, H.Y.; Lee, J.R.; Kang, S.; Han, J.; Koo, J.H. T-Cell-Mimicking Nanoparticles for Cancer Immunotherapy. Adv. Mater. 2020, 32, 2003368. [Google Scholar] [CrossRef]
- Ma, W.; Zhu, D.; Li, J.; Chen, X.; Xie, W.; Jiang, X.; Wu, L.; Wang, G.; Xiao, Y.; Liu, Z. Coating biomimetic nanoparticles with chimeric antigen receptor T cell-membrane provides high specificity for hepatocellular carcinoma photothermal therapy treatment. Theranostics 2020, 10, 1281. [Google Scholar] [CrossRef] [PubMed]
- Derakhshani, A.; Rostami, Z.; Safarpour, H.; Shadbad, M.A.; Nourbakhsh, N.S.; Argentiero, A.; Taefehshokr, S.; Tabrizi, N.J.; Kooshkaki, O.; Astamal, R.V. From Oncogenic Signaling Pathways to Single-Cell Sequencing of Immune Cells: Changing the Landscape of Cancer Immunotherapy. Molecules 2021, 26, 2278. [Google Scholar] [CrossRef] [PubMed]
- Shadbad, M.A.; Hajiasgharzadeh, K.; Derakhshani, A.; Silvestris, N.; Baghbanzadeh, A.; Racanelli, V.; Baradaran, B. From Melanoma Development to RNA-Modified Dendritic Cell Vaccines: Highlighting the Lessons from the Past. Front. Immunol. 2021, 12, 331. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-Y.; Zhang, J.; Hou, L.-D.; Zhang, R.; Chen, W.; Fan, H.-N.; Huang, Y.-X.; Liu, H.; Zhu, J.-S. Upregulation of PD-L1 predicts poor prognosis and is associated with miR-191-5p dysregulation in colon adenocarcinoma. Int. J. Immunopathol. Pharmacol. 2018, 32, 2058738418790318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, S.; Zhu, Y.; Ai, F.; Li, Y.; Bai, B.; Yao, W.; Dong, L. MicroRNA-191 correlates with poor prognosis of colorectal carcinoma and plays multiple roles by targeting tissue inhibitor of metalloprotease 3. Neoplasma 2014, 61, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.-F.; Li, K.-k.; Gao, L.; Li, S.-Z.; Chen, K.; Zhang, J.-B.; Wang, D.; Tu, R.-F.; Zhang, J.-X.; Tao, K.-X. miR-191 promotes tumorigenesis of human colorectal cancer through targeting C/EBPβ. Oncotarget 2015, 6, 4144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Chen, J.-S.; Wen, J.-K.; Gao, H.-T.; Zheng, B.; Qu, C.-B.; Liu, K.-L.; Zhang, M.-L.; Gu, J.-F.; Li, J.-D. Silencing of miR-193a-5p increases the chemosensitivity of prostate cancer cells to docetaxel. J. Exp. Clin. Cancer Res. 2017, 36, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Xu, X.; Xu, X.; Li, S.; Liang, Z.; Hu, Z.; Wu, J.; Zhu, Y.; Jin, X.; Wang, X. MicroRNA-193a-3p inhibits cell proliferation in prostate cancer by targeting cyclin D1. Oncol. Lett. 2017, 14, 5121–5128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansoori, B.; Duijf, P.H.; Mohammadi, A.; Safarzadeh, E.; Ditzel, H.J.; Gjerstorff, M.F.; Cho, W.C.-S.; Baradaran, B. MiR-142-3p targets HMGA2 and suppresses breast cancer malignancy. Life Sci. 2021, 276, 119431. [Google Scholar] [CrossRef]
- Zhu, J.; Tian, X.; Zhang, Y.; Bai, Y.; Pei, K. MiR-142-3p is a tumor promoter by directly targeting FOXO4 to promote gastric cancer cell growth and suppress cell apoptosis in gastric cancer. Int. J. Clin. Exp. Pathol. 2016, 9, 12079–12087. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| No. | Excluded Study in the Second Phase | The Reason for Its Exclusion |
|---|---|---|
| 1 | Peng et al. [18] | They were solely based on bioinformatics. |
| 2 | Qattan et al. [19] | |
| 3 | Liu et al. [20] | |
| 4 | Naba et al. [21] | The PD-L1-inhibiting miRs were not adequately investigated. |
| 5 | Yang et al. [22] | |
| 6 | Noman et al. [23] | |
| 7 | Zhang et al. [24] | |
| 8 | Nafea et al. [25] | |
| 9 | Youness et al. [26] | |
| 10 | Hamed et al. [27] | Pro-PD-L1 miR was studied. |
| 11 | Yao et al. [28] | The cross-talk between miR and PD-L1 in macrophages was studied. |
| 12 | Rogers, 2018 [29] | The cross-talk between miR and PD-L1 was evaluated only in murine TNBC. |
| No. | First Author and Year | The Studied miR | The Effect of Studied miR on TNBC Cell/the Tumor Microenvironment of TNBC | The Studied Cell Line(s) |
|---|---|---|---|---|
| 1 | Zhou, 2021 [12] | miR-424-5p | This miR can inhibit tumoral PD-L1, induce a pro-inflammatory tumor microenvironment, and stimulate apoptosis in vitro. Also, its intratumoral administration can decrease tumor size in animal models. | MDA-MB-231 |
| 2 | Dastmalchi, 2021 [30] | miR-424-5p | This miR can inhibit PD-L1 expression and enhance the chemosensitivity of tumoral cells to taxol. The combination therapy with miR-424-5p and taxol has shown superiority in terms of stimulating apoptosis, arresting cell-cycle, suppressing the clonogenicity of tumoral cells, and inhibiting the PI3K/Akt signaling pathway over monotherapy with taxol. | MDA-MB-231 |
| 3 | Soufiani, 2021 [31] | miR-34a | This miR can substantially inhibit tumoral PD-L1 expression, decrease tumor migration, and stimulate apoptosis in tumoral cells. | MDA-MB-231 |
| 4 | Rasoolnezhad, 2021 [32] | miR-138-5p | This miR can downregulate PD-L1 expression and inhibit the PI3K/Akt signaling pathway. Restoration of miR-138-5p has been associated with decreased MMP-2, MMP-9, and vimentin and increased E-cadherin expression. Besides, its restoration has stimulated apoptosis, arrested cell-cycle, upregulated INF-γ/TNF-α, and downregulated IL-10 in a co-culture system with T-cells. | MDA-MB-231 |
| 5 | Wang, 2020 [13] | miR-570-3p | This miR can inhibit PD-L1 expression, stimulate apoptosis, and decrease tumor proliferation and migration via inhibiting the PI3K/Akt/mTOR signaling pathway. | MDA-MB-231, and MDA-MB-468 |
| 6 | Peng, 2020 [33] | miR-200c-3p | The miR-200c-3p inhibitor can stimulate PD-L1 expression. | MDA-MB-231, and BT549 |
| 7 | Dastmalchi, 2020 [34] | miR-424-5p | This miR can stimulate autophagy and apoptosis in tumoral cells. Its overexpression can decrease tumor proliferation via the suppressing of the PI3K/Akt pathway, arresting the cell cycle, inhibiting colony-formation of tumor cells, and repressing angiogenesis. With the inhibitory effect on tumoral PD-L1, this miR can upregulate INF-γ, TNF-α, and IL-2 and downregulate IL-10 expression in co-cultured T-cells. | MDA-MB-231 |
| 8 | Azarbarzin, 2021 [35] | miR-383-5p | The restoration of this miR can inhibit PD-L1 expression, tumor invasion, clonogenicity, and proliferation of tumoral cells. Furthermore, this miR can arrest the cell cycle and stimulate apoptosis. In a co-culture system with T-cells, this miR can upregulate the expression of pro-inflammatory cytokines and downregulate the anti-inflammatory cytokines. The anti-tumoral effects of this miR might be attributable to its inhibitory effect on the PI3k/Akt signaling pathway. | MDA-MB-231 |
| 9 | Zhao, 2019 [36] | miR-34a-5p | This miR can suppress tumoral PD-L1 expression, leading to the inhibition of tumor proliferation and migration both in vitro and in vivo. | MDA-MB-231 |
| 10 | Li, 2019 [37] | miR-3609 | This miR can inhibit the tumoral PD-L1 and improve the survival of mice bearing TNBC. | MDA-MB-231, and MDA-MB-468 |
| 11 | Yang, 2018 [38] | miR-195-5p | This miR can downregulate tumoral PD-L1 expression in TNBC. | MDA-MB-231 |
| 12 | Yang, 2018 [38] | miR-497-5p | This miR can downregulate tumoral PD-L1 expression in TNBC. | MDA-MB-231 |
| 13 | Huang, 2017 [6] | miR-34a | This miR can inhibit tumoral PD-L1 and decrease the viability and invasion of tumor cells. In animal models, miR-34a restoration can inhibit tumoral PD-L1, increase CD8+ and CD4+ T-cells, and inhibit the recruitment of macrophages and Tregs into the tumor microenvironment. | MDA-MB-231, and HCC38 |
| No. | First Author, Publication Year | Was the Studied Cancer Cell Line(s) Reported? | Was the Duration of Exposure to the Studied miR to Tumoral Cells Reported? | Was the ConCentration of the Studied miR Reported? | Was a Standard Culture Media Used for the Study? | Were Reliable Tools Used to Assess the Outcome? | Were the Experiments Conducted More than Once? | Was More than One Independent Experiment Performed? | The Overall Risk of Bias |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Narges Dastmalchi, 2021 [30] | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias |
| 2 | Katayoun Bahman Soufiani, 2021 [31] | Without bias | Without bias | With bias | Without bias | Without bias | Without bias | Without bias | Low-bias |
| 3 | Mina Rasoolnezhad, 2021 [32] | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias |
| 4 | Li-Li Wang, 2020 [13] | Without bias | Without bias | With bias | Without bias | Without bias | Without bias | Without bias | Low-bias |
| 5 | Fu Peng, 2020 [33] | Without bias | With bias | With bias | Without bias | Without bias | Without bias | Without bias | Low-bias |
| 6 | Narges Dastmalchi, 2020 [34] | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias |
| 7 | Shirin Azarbarzin, 2021 [35] | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias |
| 8 | Lianzhou Yang, 2018 [38] | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias | Without bias |
| 9 | Xiaojia Huang, 2017 [6] | Without bias | Without bias | With bias | Without bias | Without bias | Without bias | Without bias | Low-bias |
| No. | First Author and Publication Year | Sequence Generation | Baseline Characteristics | Allocation Concealment | Random Housing | Blinding (Performance Bias) | Random Outcome Assessment | Blinding (Detection Bias) | Incomplete Outcome Data | Selective Outcome Reporting | Other Sources of Bias |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Yueyuan Zhou, 2021 [12] | No bias | No bias | No bias | With Bias | No bias | No bias | With Bias | No bias | No bias | No bias |
| 2 | Qiuyang Zhao, 2019 [36] | No bias | No bias | No bias | With Bias | No bias | No bias | With Bias | No bias | No bias | No bias |
| 3 | Duolu Li, 2019 [37] | No bias | No bias | No bias | No bias | No bias | No bias | With Bias | No bias | No bias | No bias |
| No. | Intervention | Mechanism of Action | Phase | Study Start Date | The Status | Clinicaltrials.gov Identifier |
|---|---|---|---|---|---|---|
| 1 | Atezolizumab and Paclitaxel | PD-L1 blockade and disrupting mitosis | III | 25 Aug 2017 | Active, not recruiting | NCT03125902 |
| 2 | Atezolizumab and nab-Paclitaxel | PD-L1 blockade and disrupting mitosis | III | 17 Dec 2019 | Recruiting | NCT04148911 |
| 3 | Atezolizumab and nab-Paclitaxel | PD-L1 blockade and disrupting mitosis | III | 23 Jun 2015 | Active, not recruiting | NCT02425891 |
| 4 | Atezolizumab, radiation, and Talazoparib | PD-L1 blockade and inducing DNA damage | II | 1 Apr 2021 | Recruiting | NCT04690855 |
| 5 | Atezolizumab, Paclitaxel, Doxorubicin/Epirubicin, and Cyclophosphamide | PD-L1 blockade, disrupting mitosis, DNA synthesis inhibition, and protein synthesis inhibition | III | 2 Aug 2018 | Recruiting | NCT03498716 |
| 6 | Avelumab | PD-L1 blockade | III | Jun 2016 | Active, not recruiting | NCT02926196 |
| 7 | Atezolizumab | PD-L1 blockade | III | 19 Dec 2017 | Recruiting | NCT03281954 |
| 8 | Atezolizumab, Pegylated liposomal doxorubicin, and Cyclophosphamide | PD-L1 blockade, DNA synthesis inhibition, and protein synthesis inhibition | II | 1 Jun 2017 | Recruiting | NCT03164993 |
| 9 | Durvalumab and Olaparib | PD-L1 blockade and DNA repair inhibition | II | 4 Oct 2018 | Active, not recruiting | NCT03167619 |
| 10 | Avelumab and Palbociclib | PD-L1 blockade and inhibiting DNA replication | I | 11 Aug 2020 | Recruiting | NCT04360941 |
| 11 | Atezolizumab, Bevacizumab, Gemcitabine, and Carboplatin | PD-L1 blockade, inhibiting angiogenesis, DNA synthesis inhibition, and suppressing DNA synthesis | II | Feb 2021 | Not yet recruiting | NCT04739670 |
| 12 | Durvalumab and Carboplatin | PD-L1 blockade and DNA synthesis inhibition | II | 29 Aug 2017 | Active, not recruiting | NCT03206203 |
| 13 | Durvalumab, Oleclumab, Paclitaxel, and Carboplatin | PD-L1 blockade, CD73 blockade, disrupting mitosis, and DNA synthesis inhibition | I/II | 28 Dec 2018 | Recruiting | NCT03616886 |
| 14 | Atezolizumab and nab-Paclitaxel | PD-L1 blockade and disrupting mitosis | II | 4 Feb 2016 | Active, not recruiting | NCT02530489 |
| 15 | Durvalumab and CFI-400945 | PD-L1 blockade and PLK4 inhibition | II | 19 Dec 2019 | Recruiting | NCT04176848 |
| 16 | Durvalumab | PD-L1 blockade | I/II | Nov 2015 | Active, not recruiting | NCT02489448 |
| 17 | Atezolizumab and Capecitabine | PD-L1 blockade and inhibiting DNA synthesis | II | 15 Jan 2019 | Recruiting | NCT03756298 |
| 18 | Atezolizumab, Ipatasertib, and Paclitaxel | PD-L1 blockade, Akt inhibition and disrupting mitosis | III | 25 Nov 2019 | Active, not recruiting | NCT04177108 |
| 19 | Atezolizumab, Bevacizumab, and Paclitaxel | PD-L1 blockade, inhibiting angiogenesis and disrupting mitosis | II | 5 Oct 2020 | Recruiting | NCT04408118 |
| 20 | Atezolizumab, Gemcitabine, Capecitabine, and Carboplatin | PD-L1 blockade, DNA synthesis inhibition, inhibiting DNA synthesis and suppressing DNA synthesis | III | 11 Jan 2018 | Recruiting | NCT03371017 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shadbad, M.A.; Safaei, S.; Brunetti, O.; Derakhshani, A.; Lotfinejad, P.; Mokhtarzadeh, A.; Hemmat, N.; Racanelli, V.; Solimando, A.G.; Argentiero, A.; et al. A Systematic Review on the Therapeutic Potentiality of PD-L1-Inhibiting MicroRNAs for Triple-Negative Breast Cancer: Toward Single-Cell Sequencing-Guided Biomimetic Delivery. Genes 2021, 12, 1206. https://doi.org/10.3390/genes12081206
Shadbad MA, Safaei S, Brunetti O, Derakhshani A, Lotfinejad P, Mokhtarzadeh A, Hemmat N, Racanelli V, Solimando AG, Argentiero A, et al. A Systematic Review on the Therapeutic Potentiality of PD-L1-Inhibiting MicroRNAs for Triple-Negative Breast Cancer: Toward Single-Cell Sequencing-Guided Biomimetic Delivery. Genes. 2021; 12(8):1206. https://doi.org/10.3390/genes12081206
Chicago/Turabian StyleShadbad, Mahdi Abdoli, Sahar Safaei, Oronzo Brunetti, Afshin Derakhshani, Parisa Lotfinejad, Ahad Mokhtarzadeh, Nima Hemmat, Vito Racanelli, Antonio Giovanni Solimando, Antonella Argentiero, and et al. 2021. "A Systematic Review on the Therapeutic Potentiality of PD-L1-Inhibiting MicroRNAs for Triple-Negative Breast Cancer: Toward Single-Cell Sequencing-Guided Biomimetic Delivery" Genes 12, no. 8: 1206. https://doi.org/10.3390/genes12081206
APA StyleShadbad, M. A., Safaei, S., Brunetti, O., Derakhshani, A., Lotfinejad, P., Mokhtarzadeh, A., Hemmat, N., Racanelli, V., Solimando, A. G., Argentiero, A., Silvestris, N., & Baradaran, B. (2021). A Systematic Review on the Therapeutic Potentiality of PD-L1-Inhibiting MicroRNAs for Triple-Negative Breast Cancer: Toward Single-Cell Sequencing-Guided Biomimetic Delivery. Genes, 12(8), 1206. https://doi.org/10.3390/genes12081206