
Prospects in Connecting Genetic Variation to Variation in Fertility in Male Bees


Abstract
:1. Introduction
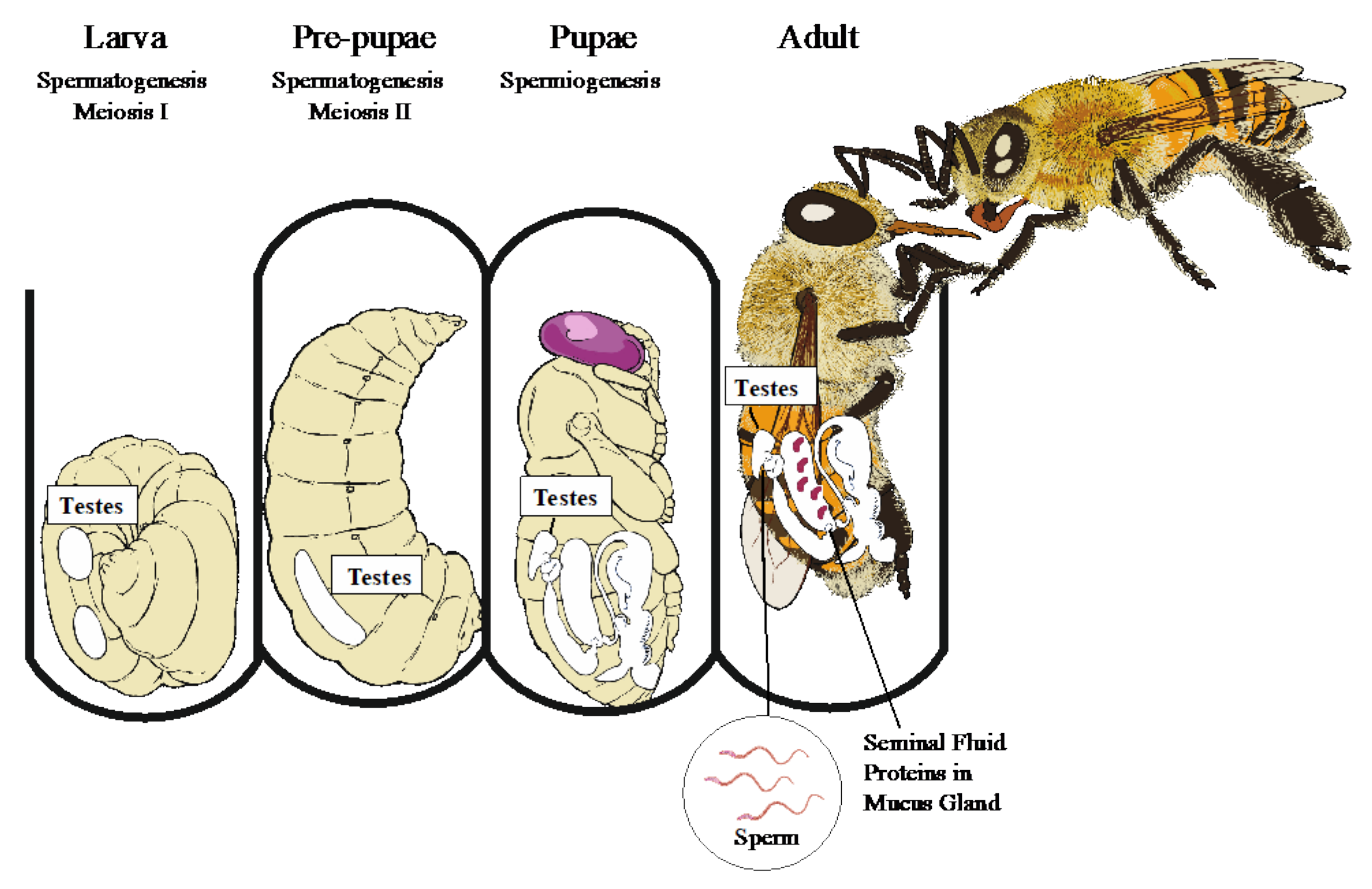
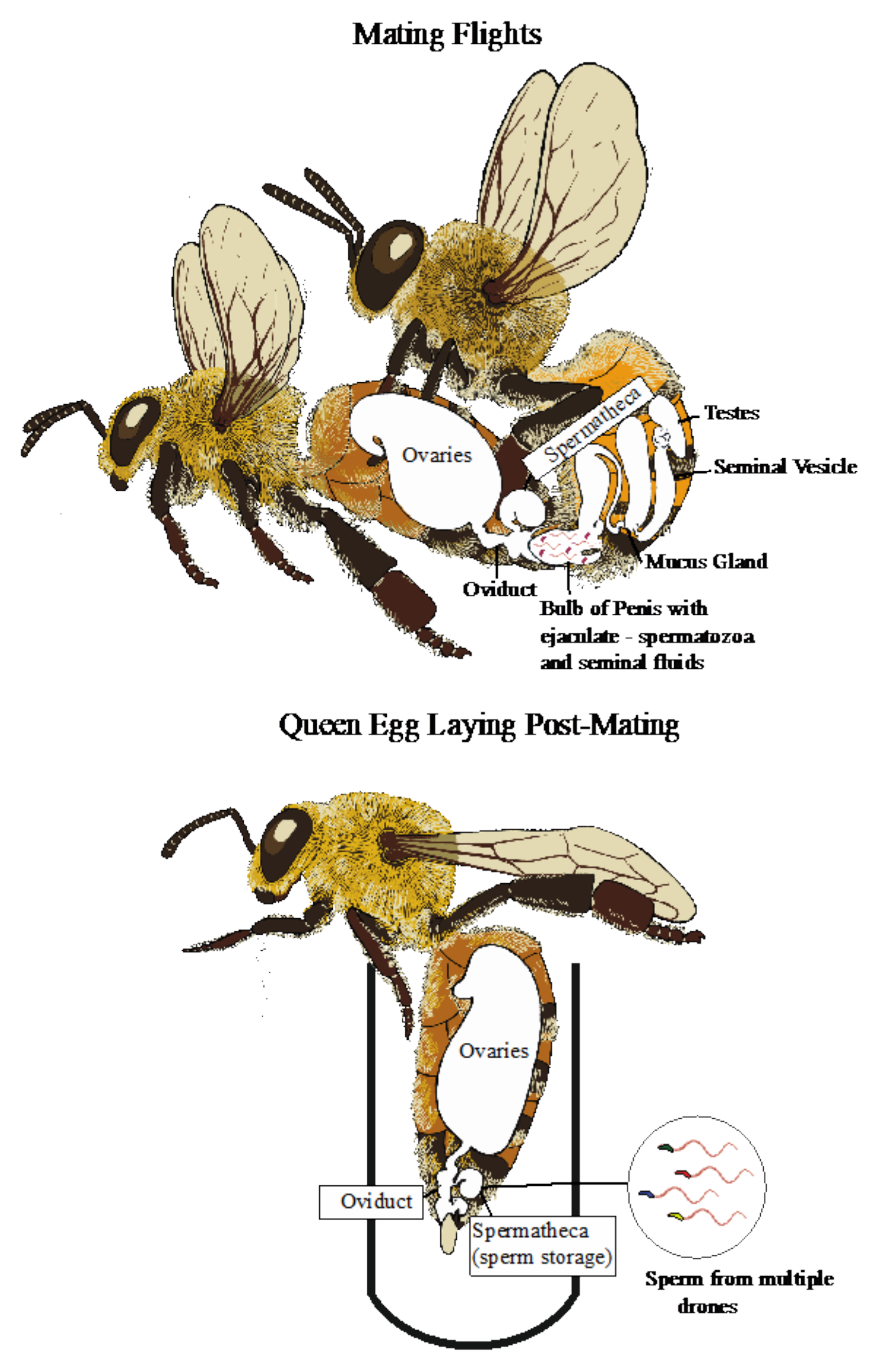
2. An Overview of Male Reproductive Biology in Honey Bees
3. Sperm Traits Vary Considerably among Honey Bee Populations
4. Connecting Genetic Variation to Phenotypic Variation
5. Importance and Determinants of Sperm Concentration and Quality to Drones
5.1. Tubule Number
5.2. Spermatogenesis Quality
5.3. Migration from the Testes
5.4. Ejaculation and Sperm Movement
5.5. Sperm Storage by the Queen
6. Importance of Sperm Morphology to Drone Fertility
Spermiogenesis
7. Importance of Seminal Fluids to Drone Fertility
8. Future Directions
8.1. On the Need to Connect Sperm Quality to Fertility in Bees
8.2. Identifying Candidate Genes
Funding
Acknowledgments
Conflicts of Interest
References
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2008, 68, 810–821. [Google Scholar] [CrossRef]
- Michener, C.D. The Bees of the World; John Hopkins University Press: Baltimore, MD, USA, 2007. [Google Scholar]
- Hein, L. Economic value of the pollination service, a review across scales. Open Ecol. J. 2009, 2, 74–82. [Google Scholar] [CrossRef]
- Carpenter, M.H.; Harpur, B.A. Genetic past, present, and future of the honey bee (Apis mellifera) in the United States of America. Apidologie 2021, 52, 63–79. [Google Scholar] [CrossRef]
- Bohart, G.E. How to Manage the Alfalfa Leaf-Cutting Bee (Megachile Rotundata Fabr.) for Alfalfa Pollination. Utah Agric. Exp. Station Circ. 1963, 144, 1. [Google Scholar]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and conservation of bumble bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Nayak, R.K.; Rana, K.; Bairwa, V.K.; Singh, P.; Bharthi, V.D. A review on role of bumblebee pollination in fruits and vegetables. Phytopathology 2020, 9, 1328–1334. [Google Scholar] [CrossRef]
- Reilly, J.R.; Artz, D.R.; Biddinger, D.; Bobiwash, K.; Boyle, N.K.; Brittain, C.; Brokaw, J.; Campbell, J.W.; Daniels, J.; Elle, E.; et al. Crop production in the USA is frequently limited by a lack of pollinators: Pollination limitation in US crops. Proc. R. Soc. B Biol. Sci. 2020, 287, 20200922. [Google Scholar] [CrossRef]
- Kremen, C.; Chaplin-Kramer, R. Insects as Providers of Ecosystem Services: Crop Pollination and Pest Control. In Insect Conservation Biology: Proceedings of The Royal Entomological Society’s 23rd Symposium; CABI Publishing: Oxfordshire, UK, 2007; pp. 349–382. ISBN 9781845932541. [Google Scholar]
- Hristov, P.; Shumkova, R.; Palova, N.; Neov, B. Factors associated with honey bee colony losses: A mini-review. Vet. Sci. 2020, 7, 166. [Google Scholar] [CrossRef]
- Pettis, J.S.; Rice, N.; Joselow, K. Colony Failure Linked to Low Sperm Viability in Honey Bee (Apis mellifera) Queens and an Exploration of Potential Causative Factors. PLoS ONE 2016, 11, e0147220. [Google Scholar] [CrossRef]
- Sandrock, C.; Tanadini, M.; Tanadini, L.G.; Fauser-Misslin, A.; Potts, S.G.; Neumann, P. Impact of chronic neonicotinoid exposure on honeybee colony performance and queen supersedure. PLoS ONE 2014, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kairo, G.; Provost, B.; Tchamitchian, S.; Ben Abdelkader, F.; Bonnet, M.; Cousin, M.; Sénéchal, J.; Benet, P.; Kretzschmar, A.; Belzunces, L.P.; et al. Drone exposure to the systemic insecticide Fipronil indirectly impairs queen reproductive potential. Sci. Rep. 2016, 6, 31904. [Google Scholar] [CrossRef] [Green Version]
- Rousseau, A.; Fournier, V.; Giovenazzo, P. Apis mellifera (Hymenoptera: Apidae) drone sperm quality in relation to age, genetic line, and time of breeding. Can. Entomol. 2015, 147, 702–711. [Google Scholar] [CrossRef]
- Brutscher, L.M.; Baer, B.; Niño, E.L.; Brutscher, L.M.; Baer, B.; Niño, E.L. Putative Drone Copulation Factors Regulating Honey Bee (Apis mellifera) Queen Reproduction and Health: A Review. Insects 2019, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Rangel, J.; Fisher, A. Factors affecting the reproductive health of honey bee (Apis mellifera) drones—A review. Apidologie 2019, 50, 759–778. [Google Scholar] [CrossRef] [Green Version]
- Belsky, J.E.; Camp, A.A.; Lehmann, D.M. The Importance of Males to Bumble Bee Colony Viability. Insects 2020, 11, 506. [Google Scholar] [CrossRef]
- Wilson, E.O. The Insect Societies; Belknap press of Harvard University press: Cambridge, MA, USA, 1971. [Google Scholar]
- Slater, G.P.; Yocum, G.D.; Bowsher, J.H.; Bowsher, J.H. Diet quantity influences caste determination in honeybees (Apis mellifera). Proc. R. Soc. B 2020, 287, 20200614. [Google Scholar] [CrossRef]
- Heidinger, I.M.M.; Meixner, M.D.; Berg, S.; Büchler, R.; Buchler, R. Observation of the Mating Behavior of Honey Bee (Apis mellifera L.) Queens Using Radio-Frequency Identification (RFID): Factors Influencing the Duration and Frequency of Nuptial Flights. Insects 2014, 5, 513–527. [Google Scholar] [CrossRef] [Green Version]
- Tarpy, D.R.; Nielsen, R.; Nielsen, D.I. A scientific note on the revised estimates of effective paternity frequency in Apis. Insectes Soc. 2004, 51, 203–204. [Google Scholar] [CrossRef]
- Kocher, S.D.; Richard, F.; Tarpy, D.R.; Grozinger, C.M. Genomic analysis of post-mating changes in the honey bee queen (Apis mellifera). BMC Genom. 2008, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mark, L.W. The Biology of The Honey Bee; Harvard University Press: Cambridge, MA, USA, 1991. [Google Scholar]
- Szabo, T.I.; Heikel, D.T. Numbers of Spermatozoa in Spermathecae of Queens Aged 0 to 3 Years Reared in Beaverlodge, Alberta. J. Apic. Res. 1987, 26, 79–82. [Google Scholar] [CrossRef]
- Al-Lawati, H.; Kamp, G.; Bienefeld, K. Characteristics of the spermathecal contents of old and young honeybee queens. J. Insect Physiol. 2009, 55, 117–122. [Google Scholar] [CrossRef]
- Lago, D.C.; Martins, J.R.; Dallacqua, R.P.; Santos, D.E.; Bitondi, M.M.G.; Hartfelder, K. Testis development and spermatogenesis in drones of the honey bee, Apis mellifera L. Apidologie 2020, 51, 935–955. [Google Scholar] [CrossRef]
- Jaycox, E.R. The Effects of Various Foods and Temperatures on Sexual Maturity of the Drone Honey Bee (Apis mellifera). Ann. Entomol. Soc. Am. 1961, 54, 519–523. [Google Scholar] [CrossRef]
- Garofalo, C.A. Comportamento e maturidade sexual de zangôes de Apis mellifera adansonii. In Proceedings of Homenagen à Warwick E. Kerr., Botucatu, Brazil, 8–9 September 1972; pp. 8–9. [Google Scholar]
- Woyke, J. Multiple Mating of the Honeybee Queen (Apis mellifica L.) in One Nuptial Flight. Bull. L’Académie Pol. Des Sci. 1955, 3, 175–180. [Google Scholar]
- Reyes, M.; Crauser, D.; Prado, A.; Le Conte, Y. Flight activity of honey bee (Apis mellifera) drones. Apidologie 2019, 50, 669–680. [Google Scholar] [CrossRef]
- Woodgate, J.L.; Makinson, J.C.; Rossi, N.; Lim, K.S.; Reynolds, A.M.; Rawlings, C.J.; Chittka, L. Harmonic radar tracking reveals that honeybee drones navigate between multiple aerial leks. iScience 2021, 102499. [Google Scholar] [CrossRef]
- Koeniger, N.; Koeniger, G.; Gries, M.; Tingek, S. Drone competition at drone congregation areas in four Apis species. Apidologie 2007, 36, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Zmarlicki, C.; Morse, R.A. Drone congregation areas. J. Apic. Res. 1963, 2, 64–66. [Google Scholar] [CrossRef]
- Olonelloa, N.A.; Artfelderb, K.H. She’s my girl—Male accessory gland products and their function in the reproductive biology of social bees. Apidologie 2005, 36, 231–244. [Google Scholar] [CrossRef]
- Koeninger, N.; Koenger, G. Reproductive isolation among species of the genus Apis. Apidologie 2000, 31, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Koeniger, N.; Koeninger, G.; Wongsiri, S. Mating and Sperm Transfer in Apis florea. Apidologie 1989, 20, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Baer, B. Sexual selection in Apis bees. Apidologie 2005, 36, 187–200. [Google Scholar] [CrossRef] [Green Version]
- Quickie, D.L.J.; Ingram, S.N.; Baillie, H.S.; Gaitens, P.V. Sperm structure and ultrastructure in the Hymenoptera (Insecta). Zool. Scr. 1992, 21, 381–402. [Google Scholar] [CrossRef]
- Simmons, L.W.; Fitzpatrick, J.L. Sperm wars and the evolution of male fertility. Reproduction 2012, 144, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Baer, B.; Collins, J.; Maalaps, K.; den Boer, S.P.A. Sperm use economy of honeybee (Apis mellifera) queens. Ecol. Evol. 2016, 6, 2877–2885. [Google Scholar] [CrossRef]
- Rousseau, A.; Giovenazzo, P. Optimizing Drone Fertility With Spring Nutritional Supplements to Honey Bee (Hymenoptera: Apidae) Colonies. J. Econ. Entomol. 2016, 109, 1009–1014. [Google Scholar] [CrossRef]
- Hoage, T.R.; Kessel, R.G. An electron microscope study of the process of differentiation during spermatogenesis in the drone honey bee (Apis mellifera L.) with special reference to centriole replication and elimination. J. Ultrasructure Res. 1968, 24, 6–32. [Google Scholar] [CrossRef]
- da Cruz-Landim, C. Organization of the cysts in bee (Hymenoptera, Apidae) testis: Number of spermatozoa per cyst. Iheringia Série Zool. 2001, 91, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.; Gupta, B.L.; Kumbkarni, C.G. XXVII.-Cytology of spermatogenesis in the honey bee, Apis indica (F.). J. R. Microsc. Soc. 1961, 79, 337–351. [Google Scholar] [CrossRef]
- Colonello, N.A.; Hartfelder, K. Protein content and pattern during mucus gland maturation and its ecdysteroid control in honey bee drones. Apidologie 2003, 34, 257–267. [Google Scholar] [CrossRef]
- Blum, M.S.; Glowska, Z.; Taber, S., III. Chemistry of the drone honey bee reproductive system. II. Carbohydrates in the reproductive organs and semen. Ann. Entomol. Soc. Am. 1962, 55, 135–139. [Google Scholar] [CrossRef]
- Blum, M.S.; Bumgarner, J.E.; Taber, S., III. Composition and possible significance of fatty acids in the lipid classes in honey bee semen. J. Insect Physiol. 1967, 13, 1301–1308. [Google Scholar] [CrossRef]
- Baer, B.; Heazlewood, J.L.; Taylor, N.L.; Eubel, H.; Millar, A.H. The seminal fluid proteome of the honeybee Apis mellifera. Proteomics 2009, 9, 2085–2097. [Google Scholar] [CrossRef] [PubMed]
- den Boer, S.P.A.; Baer, B.; Boomsma, J.J. Seminal Fluid Mediates Ejaculate Competition in Social Insects. Science (80-) 2010, 327, 1506–1510. [Google Scholar] [CrossRef] [Green Version]
- Czekońska, K.; Chuda-Mickiewicz, B.; Samborski, J. Quality of honeybee drones reared in colonies with limited and unlimited access to pollen. Apidologie 2015, 46, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Schluns, H.; Schluns, E.A.; van Praagh, J.; Moritz, R.F.A. Sperm numbers in drone honeybees (Apis mellifera) depend on body size. Apidologie 2003, 34, 577–584. [Google Scholar] [CrossRef] [Green Version]
- Burley, L.M. The Effects of Miticides on the Reproductive Physiology of Honey Bee (Apis mellifera L.) Queens and Drones. Doctoral dissertation, Virginia Tech, Blacksburg, VA, USA, 2007. [Google Scholar]
- Shoukry, R.S.; Khattaby, A.M.; El-Sheakh, A.A.; Abo-Ghalia, A.H.; Elbanna, S.M. Effect of some materials for controlling varroa mite on the honey bee drones (Apis mellifera L.). Egypt. J. Agric. Res. 2013, 91, 825–834. [Google Scholar]
- Lars, S.; Laura, V.-B.; Selina, B.; Panuwan, C.; Laurent, G.; Kitiphong, K.; Gina, R.; Aline, T.; Beatriz, V.; Peter, N.; et al. Neonicotinoid insecticides can serve as inadvertent insect contraceptives. Proc. R. Soc. B-Biol. Sci. 2016, 283, 20160506. [Google Scholar] [CrossRef] [Green Version]
- Taha, E.A.; Alqarni, A.S. Morphometric and Reproductive Organs Characters of Apis mellifera jemenitica drones in Comparison to Apis mellifera carnica. Int. J. Sci. Eng. Res. 2014, 4, 411–415. [Google Scholar]
- Quartuccio, M.; Cristarella, S.; Scrofani, A.; Biondi, V.; De Majo, M.; Mannarino, C.; Cravana, C.; Medica, P.; Fazio, E. The sperm of Apis mellifera siciliana and Apis mellifera ligustica: A preliminary and comparative note. J. Apic. Res. 2020, 59, 1011–1016. [Google Scholar] [CrossRef]
- Rinderer, T.E.; Collins, A.M.; Daniel, R.; Lancaster, V. A comparison of africanized and european drones: Weights, mucus gland and seminal vesicle weights, and counts of spermatozoa. Apidologie 1985, 16, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Stürup, M.; Baer-Imhoof, B.; Nash, D.R.; Boomsma, J.J.; Baer, B. When every sperm counts: Factors affecting male fertility in the honeybee Apis mellifera. Behav. Ecol. 2013, 24, 1192–1198. [Google Scholar] [CrossRef] [Green Version]
- Baer, B.; Schmid-Hempel, P.; Høeg, J.T.; Boomsma, J.J. Sperm length, sperm storage and mating system characteristics in bumblebees. Insectes Soc. 2003, 50, 101–108. [Google Scholar] [CrossRef]
- Rhodes, J.W.; Harden, S.; Spooner-Hart, R.; Anderson, D.L.; Wheen, G. Effects of age, season and genetics on semen and sperm production in Apis mellifera drones. Apidologie 2011, 42, 29–38. [Google Scholar] [CrossRef]
- Grassl, J.; Peng, Y.; Baer-Imhoof, B.; Welch, M.; Millar, A.H.; Baer, B. Infections with the Sexually Transmitted Pathogen Nosema apis Trigger an Immune Response in the Seminal Fluid of Honey Bees (Apis mellifera). J. Proteome Res. 2017, 16, 319–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinderer, T.E.; Oldroyd, B.P.; Wongsiri, S.; Sylvester, H.A.; de Guzman, L.I.; Potichot, S.; Sheppard, W.S.; Buchmann, S.L. Time of drone flight in four honey bee species in south-eastern Thailand. J. Apic. Res. 1993, 32, 27–33. [Google Scholar] [CrossRef]
- Li-Byarlay, H.; Huang, M.H.; Simone-Finstrom, M.; Strand, M.K.; Tarpy, D.R.; Rueppell, O. Honey bee (Apis mellifera) drones survive oxidative stress due to increased tolerance instead of avoidance or repair of oxidative damage. Exp. Gerontol. 2016, 83, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Woyke, J. Lengths of haploid and diploid spermatozoa of the honeybee and the question of the production of triploid workers. J. Apic. Res. 1983, 22, 146–149. [Google Scholar] [CrossRef]
- Zaitoun, S.; Al-Majeed Al-Ghzawi, A.; Kridli, R. Monthly changes in various drone characteristics of Apis mellifera ligustica and Apis mellifera syriaca. Entomol. Sci. 2009, 12, 208–214. [Google Scholar] [CrossRef]
- Gençer, H.V.; Kahya, Y. Sperm competition in honey bees (Apis mellifera L.): The role of body size dimorphism in drones. Apidologie 2020, 51, 1–17. [Google Scholar] [CrossRef]
- Gençer, H.V.; Kahya, Y. The viability of sperm in lateral oviducts and spermathecae of instrumentally inseminated and naturally mated honey bee (Apis mellifera L.) queens. J. Apic. Res. 2011, 50, 190–194. [Google Scholar] [CrossRef]
- Jarolimek, J.; Otis, G.W. A comparison of fitness components in large and small honey-bee drones. Am. Bee J. 2001, 141, 891–892. [Google Scholar]
- Pizzari, T.; Foster, K.R. Sperm sociality: Cooperation, altruism, and spite. PLoS Biol. 2008, 6, 0925–0931. [Google Scholar] [CrossRef]
- Garofalo, C.A. Reproductive aspects and evolution of social behavior in bees. Rev. Bras. Genet. 1980, 3, 139–152. [Google Scholar]
- Starr, C.K. Sperm Competition, Kinship, and Sociality in the Aculeate Hymenoptera. In Sperm Competition and the Evolution of Animal Mating Systems; Elsevier: Amsterdam, The Netherlands; CABI Publishing: Oxfordshire, UK, 1984; pp. 428–464. [Google Scholar]
- Woyke, J. Reproductive Organs of Haploid and Diploid Drone Honeybees. J. Apic. Res. 1973, 12, 35–51. [Google Scholar] [CrossRef]
- Tvedte, E.S.; Forbes, A.A.; Logsdon, J.M., Jr. Retention of Core Meiotic Genes across Diverse Hymenoptera. J. Hered. 2017, 791–806. [Google Scholar] [CrossRef]
- Schurko, A.M.; Mazur, D.J.; Logsdon, J.M., Jr. Inventory and phylogenomic distribution of meiotic genes in Nasonia vitripennis and among diverse arthropods. Insect Mol. Biol. 2010, 19, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Vangompel, M.J.W.; Xu, E.Y. The roles of the DAZ family in spermatogenesis. More than just translation? Spermatogenesis 2011, 1, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Sekiné, K.; Furusawa, T.; Hatakeyama, M. The boule gene is essential for spermatogenesis of haploid insect male. Dev. Biol. 2015, 399, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Orgad, S.; Rosenfeld, G.; Greenspan, R.J.; Segal, D. Courtless, the Drosophila UBC7 Homolog, Is Involved in Male Courtship Behavior and Spermatogenesis. Genetics 2000, 155, 1267–1280. [Google Scholar] [CrossRef]
- Bishop, G.H. Fertilization in the Honey-bee: The Male Sexual Organs: Their Histological Structure and Physiological Functioning. J. Exp. Zool. 1920, 31, 224–265. [Google Scholar] [CrossRef] [Green Version]
- Page, R.E.; Peng, Y.S.C. Aging and development in social insects with emphasis on the honey bee, Apis mellifera L. Exp. Gerontol. 2011, 36, 695–711. [Google Scholar] [CrossRef]
- Metz, B.N.; Tarpy, D.R.; Metz, B.N.; Tarpy, D.R. Reproductive Senescence in Drones of the Honey Bee (Apis mellifera). Insects 2019, 10, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paynter, E.; Millar, A.H.; Welch, M.; Baer-imhoof, B.; Cao, D.; Baer, B. Insights into the molecular basis of long-term storage and survival of sperm in the honeybee (Apis mellifera). Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tourmente, M.; Rowe, M.; González-Barroso, M.M.; Rial, E.; Gomendio, M.; Roldan, E.R. Postcopulatory sexual selection increases atp content in rodent spermatozoa. Evolution 2013, 67, 1838–1846. [Google Scholar] [CrossRef] [Green Version]
- Poland, V.; Eubel, H.; King, M.; Solheim, C.; Millar, A.H.; Baer, B. Stored sperm differs from ejaculated sperm by proteome alterations associated with energy metabolism in the honeybee Apis mellifera. Mol. Ecol. 2011, 20, 2643–2654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baer, B.; Eubel, H.; Taylor, N.L.; O’Toole, N.; Millar, A.H. Insights into female sperm storage from the spermathecal fluid proteome of the honeybee Apis mellifera. Genome Biol. 2009, 10, R67. [Google Scholar] [CrossRef] [Green Version]
- Wasbrough, E.R.; Dorus, S.; Hester, S.; Howard-Murkin, J.; Lilley, K.; Wilkin, E.; Polpitiya, A.; Petritis, K.; Karr, T.L. The Drosophila melanogaster sperm proteome-II (DmSP-II). J. Proteomics 2010, 73, 2171–2185. [Google Scholar] [CrossRef]
- Dorus, S.; Busby, S.A.; Gerike, U.; Shabanowitz, J.; Hunt, D.F.; Karr, T.L. Genomic and functional evolution of the Drosophila melanogaster sperm proteome. Nat. Genet. 2006, 38, 1440–1445. [Google Scholar] [CrossRef]
- Whittington, E.; Zhao, Q.; Borziak, K.; Walters, J.R.; Dorus, S. Characterisation of the Manduca sexta sperm proteome: Genetic novelty underlying sperm composition in Lepidoptera. Insect Biochem. Mol. Biol. 2015, 62, 183–193. [Google Scholar] [CrossRef]
- Degner, E.C.; Ahmed-Braimah, Y.H.; Borziak, K.; Wolfner, M.F.; Harrington, L.C.; Dorus, S. Reproductive functions and genetic architecture of the seminal fluid and sperm proteomes of the mosquito Aedes aegypti. bioRxiv 2018, 405431. [Google Scholar] [CrossRef] [Green Version]
- Rettie, E.C.; Dorus, S. Drosophila sperm proteome evolution. Spermatogenesis 2012, 2, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Laurinyecz, B.; Vedelek, V.; Kovács, A.L.; Szilasi, K.; Lipinszki, Z.; Slezák, C.; Darula, Z.; Juhász, G.; Sinka, R. Sperm-Leucylaminopeptidases are required for male fertility as structural components of mitochondrial paracrystalline material in Drosophila melanogaster sperm. PLoS Genet. 2019, 15, 1–24. [Google Scholar] [CrossRef]
- Dorus, S.; Wilkin, E.C.; Karr, T.L. Expansion and functional diversification of a leucyl aminopeptidase family that encodes the major protein constituents of Drosophila sperm. BMC Genom. 2011, 12, 171. [Google Scholar] [CrossRef] [Green Version]
- Civetta, A.; Ranz, J.M. Genetic Factors Influencing Sperm Competition. Front. Genet. 2019, 10, 820. [Google Scholar] [CrossRef] [Green Version]
- Pascini, T.V.; Martins, G.F. The insect spermatheca: An overview. Zoology 2017, 121, 56–71. [Google Scholar] [CrossRef]
- Juliana, M.; Martins, G.F.; Marques, A.E.; Games, P.D.; Zanuncio, J.C.; Baracat-Pereira, M.C.; Fernandes, T.M. IInsights into the proteome of the spermatheca of the leaf-cutting ant Atta sexdens rubropilosa (Hymenoptera: Formicidae). Florida Entomol. 2014, 97, 1856–1861. [Google Scholar]
- Weirich, G.F.; Collins, A.M.; Williams, V.P. Antioxidant enzymes in the honey bee, Apis mellifera. Apidologie 2002, 33, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Juliana, R.; Shepherd, T.F.; Gonzalez, A.N.; Hillhouse, A.; Konganti, K.; Ing, N.H. Transcriptomic analysis of the honey bee (Apis mellifera) queen spermathecae reveals genes that may be involved in sperm storage after mating. PLoS ONE 2021, 16, e0244648. [Google Scholar] [CrossRef]
- Collins, A.M.; Williams, V.; Evans, J.D. Sperm storage and antioxidative enzyme expression in the honey bee, Apis mellifera. Insect Mol. Biol. 2004, 13, 141–146. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.N.; Ing, N.; Rangel, J. Upregulation of antioxidant genes in the spermathecae of honey bee (Apis mellifera) queens after mating. Apidologie 2018, 49, 224–234. [Google Scholar] [CrossRef] [Green Version]
- Tirmarche, S.; Kimura, S.; Dubruille, R.; Horard, B.; Loppin, B. Unlocking sperm chromatin at fertilization requires a dedicated egg thioredoxin in Drosophila. Nat. Commun. 2016, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Corona, M.; Robinson, G.E. Genes of the antioxidant system of the honey bee: Annotation and phylogeny. Insect Mol. Biol. 2006, 15, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, F.; Li, G.; Chi, X.; Wang, Y.; Wang, H.; Ma, L.; Han, K.; Zhao, G.; Guo, X.; et al. Metabolite Support of Long-Term Storage of Sperm in the Spermatheca of Honeybee (Apis mellifera) Queens. Front. Physiol. 2020, 11, 1303. [Google Scholar] [CrossRef]
- Ferreira, G.; Costa, C.; Bassaizteguy, V.; Santos, M.; Cardozo, R.; Montes, J.; Settineri, R.; Nicolson, G.L. Incubation of human sperm with micelles made from glycerophospholipid mixtures increases sperm motility and resistance to oxidative stress. PLoS ONE 2018, 13, e0197897. [Google Scholar] [CrossRef] [Green Version]
- Boomsma, J.J.; Baer, B.; Heinze, J. the Evolution of Male Traits in Social Insects. Annu. Rev. Entomol. 2005, 50, 395–420. [Google Scholar] [CrossRef] [PubMed]
- Franck, P.; Solignac, M.; Vautrin, D.; Cornuet, J.-M.; Koeniger, G.; Koeniger, N. Sperm competition and last-male precedence in the honeybee. Anim. Behav. 2002, 64, 503–509. [Google Scholar] [CrossRef] [Green Version]
- Le Conte, Y.; Franck, P.; Garnery, L.; Coussy, H.; Solignac, M.; Cornuet, J.-M. Microsatellite analysis of sperm admixture in honeybee. Insect Mol. Biol. 2003, 8, 419–421. [Google Scholar] [CrossRef]
- Miller, G.T.; Pitnick, S. Sperm-female coevolution in Drosophila. Science (80-) 2002, 298, 1230–1233. [Google Scholar] [CrossRef]
- Pitnick, S.; Markow, T.; Spicer, G.S. Evolution of multiple kinds of female sperm-storage organs in drosophila. Evolution 1999, 53, 1804–1822. [Google Scholar] [CrossRef]
- Pitnick, S.; Markow, T.A.; Spicert, G.S. Delayed male maturity is a cost of producing large sperm in Drosophila. Proc. Natl. Acad. Sci. USA 1995, 92, 10614–10618. [Google Scholar] [CrossRef] [Green Version]
- Dybas, L.K.; Dybas, H.S. Coadaptation and Taxonomic Differentiation of Sperm and Spermathecae in Featherwing Beetles. Evolution 1981, 35, 168–174. [Google Scholar] [CrossRef]
- Sasakawa, K. Sperm bundle and reproductive organs of carabid beetles tribe Pterostichini (Coleoptera: Carabidae). Naturwissenschaften 2007, 94, 384–391. [Google Scholar] [CrossRef]
- Minder, A.M.; Hosken, D.J.; Ward, P.I. Co-evolution of male and female reproductive characters across the Scathophagidae (Diptera). Eur. Soc. Evol. Biol. 2005, 18, 60–69. [Google Scholar] [CrossRef]
- Martins, J.R.; Nunes, F.M.F.; Cristino, A.S.; Simões, Z.L.P.; Bitondi, M.M.G. A honey bee hexamerin, HEX 70a, is likely to play an intranuclear role in developing and mature ovarioles and testioles. BMC Mol. Biol. 2010, 11, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, J.R.; Anhezini, L.; Dallacqua, R.P.; Simões, Z.L.; Bitondi, M.M. A Honey Bee Hexamerin, HEX 70a, Is Likely to Play an Intranuclear Role in Developing and Mature Ovarioles and Testioles. PLoS ONE 2011, 6, e29006. [Google Scholar] [CrossRef] [Green Version]
- den Boer, S.P.A.; Boomsma, J.J.; Baer, B. Seminal fluid enhances sperm viability in the leafcutter ant Atta colombica. Behav. Ecol. Sociobiol. 2008, 62, 1843–1849. [Google Scholar] [CrossRef] [Green Version]
- den Boer, S.P.A.; Boomsma, J.J.; Baer, B. Honey bee males and queens use glandular secretions to enhance sperm viability before and after storage. J. Insect Physiol. 2009, 55, 538–543. [Google Scholar] [CrossRef] [Green Version]
- Liberti, J.; Gorner, J.; Welch, M.; Dosselli, R.; Schiøtt, M.; Ogawa, Y.; Castleden, I.; Hemmi, J.M.; Baer-Imhoof, B.; Boomsma, J.J.; et al. Seminal fluid compromises visual perception in honeybee queens reducing their survival during additional mating flights. Elife 2019, 8, e45009. [Google Scholar] [CrossRef] [PubMed]
- Mueller, J.L.; Page, J.L.; Wolfner, M.F. An ectopic expression screen reveals the protective and toxic effects of Drosophila seminal fluid proteins. Genetics 2007, 175, 777–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otti, O.; Naylor, R.A.; Siva-Jothy, M.T.; Reinhardt, K. Bacteriolytic activity in the ejaculate of an insect. Am. Nat. 2009, 174, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Holman, L. Drosophila melanogaster seminal fluid can protect the sperm of other males. Funct. Ecol. 2009, 23, 180–186. [Google Scholar] [CrossRef]
- Xue, L.; Noll, M. Drosophila female sexual behavior induced by sterile males showing copulation complementation. Proc. Natl. Acad. Sci. USA 2000, 97, 3272–3275. [Google Scholar] [CrossRef]
- Yang, C.H.; Rumpf, S.; Xiang, Y.; Gordon, M.D.; Song, W.; Jan, L.Y.; Jan, Y.-N. Control of the Postmating Behavioral Switch in Drosophila Females by Internal Sensory Neurons. Neuron 2009, 61, 519–526. [Google Scholar] [CrossRef] [Green Version]
- Avila, F.W.; Sirot, L.K.; LaFlamme, B.A.; Rubinstein, C.D.; Wolfner, M.F. Insect Seminal Fluid Proteins: Identification and Function. Annu. Rev. Entomol. 2011, 56, 21–40. [Google Scholar] [CrossRef] [Green Version]
- Ravi Ram, K.; Wolfner, M.F. A network of interactions among seminal proteins underlies the long-term postmating response in Drosophila. Proc. Natl. Acad. Sci. USA 2009, 106, 15384–15389. [Google Scholar] [CrossRef] [Green Version]
- Chapman, T.; Bangham, J.; Vinti, G.; Seifried, B.; Lung, O.; Wolfner, M.F.; Smith, H.K.; Partridge, L. The sex peptide of Drosophila melanogaster: Female post-mating responses analyzed by using RNA interference. Proc. Natl. Acad. Sci. USA 2003, 100, 9923–9928. [Google Scholar] [CrossRef] [Green Version]
- Ram, K.R.; Wolfner, M.F. Sustained post-mating response in Drosophila melanogaster requires multiple seminal fluid proteins. PLoS Genet. 2007, 3, e238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Boer, S.P.A.; Stürup, M.; Boomsma, J.J.; Baer, B. The ejaculatory biology of leafcutter ants. J. Insect Physiol. 2015, 74, 56–62. [Google Scholar] [CrossRef]
- Rogers, D.W.; Whitten, M.M.A.; Thailayil, J.; Soichot, J.; Levashina, E.A.; Catteruccia, F. Molecular and cellular components of the mating machinery in Anopheles gambiae females. Proc. Natl. Acad. Sci. USA 2008, 105, 19390–19395. [Google Scholar] [CrossRef] [Green Version]
- Avila, F.W.; Wolfner, M.F. Acp36DE is required for uterine conformational changes in mated Drosophila females. Proc. Natl. Acad. Sci. USA 2009, 106, 15796–15800. [Google Scholar] [CrossRef] [Green Version]
- Adams, E.M.; Wolfner, M.F. Seminal proteins but not sperm induce morphological changes in the Drosophila melanogaster female reproductive tract during sperm storage. J. Insect Physiol. 2007, 53, 319–331. [Google Scholar] [CrossRef] [Green Version]
- Wigby, S.; Brown, N.C.; Allen, S.E.; Misra, S.; Sitnik, J.L.; Sepil, I.; Clark, A.G.; Wolfner, M.F. The Drosophila seminal proteome and its role in postcopulatory sexual selection. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20200072. [Google Scholar] [CrossRef] [PubMed]
- Sirot, L.K.; Hardstone, M.C.; Helinski, M.E.H.; Ribeiro, J.M.C.; Kimura, M.; Deewatthanawong, P.; Wolfner, M.F.; Harrington, L.C. Towards a semen proteome of the dengue vector mosquito: Protein identification and potential functions. PLoS Negl. Trop. Dis. 2011, 5, e989. [Google Scholar] [CrossRef] [Green Version]
- Dottorini, T.; Nicolaides, L.; Ranson, H.; Rogers, D.W.; Crisanti, A.; Catteruccia, F. A genome-wide analysis in Anopheles gambiae mosquitoes reveals 46 male accessory gland genes, possible modulators of female behavior. Proc. Natl. Acad. Sci. USA 2007, 104, 16215–16220. [Google Scholar] [CrossRef] [Green Version]
- Simmons, L.W.; Tan, Y.; Millar, A.H. Sperm and seminal fluid proteomes of the field cricket Teleogryllus oceanicus: Identification of novel proteins transferred to females at mating. Insect Mol. Biol. 2013, 22, 15–130. [Google Scholar] [CrossRef]
- Robert, M.; Gibbs, B.F.; Jacobson, E.; Gagnon, C. Characterization of prostate-specific antigen proteolytic activity on its major physiological substrate, the sperm motility inhibitor precursor/semenogelin I. Biochemistry 1997, 36, 3811–3819. [Google Scholar] [CrossRef]
- Malm, J.; Hellman, J.; Hogg, P.; Lilja, H. Enzymatic action of prostate-specific antigen (PSA or hK3): Substrate specificity and regulation by Zn2+, a tight-binding inhibitor. Prostate 2000, 45, 132–139. [Google Scholar] [CrossRef]
- Gotoh, A.; Shigenobu, S.; Yamaguchi, K.; Kobayashi, S.; Ito, F.; Tsuji, K. Transcriptome characterization of male accessory glands in ants to identify molecules involved in their reproductive success. Insect Mol. Biol. 2018, 27, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S.; Kato, K.; Takata, Y.; Kamei, K. Identification of the sperm-activating factor initiatorin, a prostatic endopeptidase of the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2012, 42, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Miyata, H.; Thaler, C.D.; Haimo, L.T.; Cardullo, R.A. Protease activation and the signal transduction pathway regulating motility in sperm from the water strider Aquarius remigis. Cytoskeleton 2012, 69, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, K.; Naylor, R.A.; Siva-Jothy, M.T. Ejaculate components delay reproductive senescence while elevating female reproductive rate in an insect. Proc. Natl. Acad. Sci. USA 2009, 106, 21743–21747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, M.J.; Rylett, C.M.; Keen, J.N.; Audsley, N.; Sajid, M.; Shirras, A.D.; Isaac, R.E. Proteomic identification of Drosophila melanogaster male accessory gland proteins, including a pro-cathepsin and a soluble γ-glutamyl transpeptidase. Proteome Sci. 2006, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ben Abdelkader, F.; Kairo, G.; Tchamitchian, S.; Cousin, M.; Senechal, J.; Crauser, D.; Vermandere, J.P.; Alaux, C.; Le Conte, Y.; Belzunces, L.P.; et al. Semen quality of honey bee drones maintained from emergence to sexual maturity under laboratory, semi-field and field conditions. Apidologie 2014, 45, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Yániz, J.L.; Silvestre, M.A.; Santolaria, P. Sperm quality assessment in honey bee drones. Biology 2020, 9, 174. [Google Scholar] [CrossRef]
- Collins, A.M. Functional longevity of honey bee, apis mellifera, queens inseminated with low viability semen. J. Apic. Res. 2004, 43, 167–171. [Google Scholar] [CrossRef]
- Mazeed, A.M.; Mohanny, K.M. Some reproductive characteristics of honeybee drones in. Entomol. Res. 2010, 40, 245–250. [Google Scholar] [CrossRef]
- Collins, A.M.; Donoghue, A.M. Viability assessment of honey bee, Apis mellifera, sperm using dual fluorescent staining. Theriogenology 1999, 51, 1513–1523. [Google Scholar] [CrossRef]
- Oldroyd, B.P.; Cornuet, J.-M.; Rowe, D.; Rinderer, T.E.; Crozier, R.H. Racial admixture of Apis mellifera in Tasmania, Australia: Similarities and differences with natural hybrid zones in Europe. Heredity (Edinb.) 1995, 74, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Visscher, P.M.; Hill, W.G.; Wray, N.R. Heritability in the genomics era—Concepts and misconceptions. Nat. Rev. Genet. 2008, 9, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Neubaum, D.M.; Wolfner, M.F. Mated Drosophila melanogaster females require a seminal fluid protein, Acp36DE, to store sperm efficiently. Genetics 1999, 153, 845–857. [Google Scholar] [CrossRef]
- Bloch Qazi, M.C.; Wolfner, M.F. An early role for the Drosophila melanogaster male seminal protein Acp36DE in female sperm storage. J. Exp. Biol. 2003, 206, 3521–3528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, H.S.; Jacobs-Palmer, E.; Lassance, J.M.; Hoekstra, H.E. The genetic basis and fitness consequences of sperm midpiece size in deer mice. Nat. Commun. 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, C.Y.; Wolfner, M.F.; Clark, A.G. Large neurological component to genetic differences underlying biased sperm use in rosophila. Genetics 2013, 193, 177–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawano, N.; Araki, N.; Yoshida, K.; Hibino, T.; Ohnami, N.; Makino, M.; Kanai, S.; Hasuwa, H.; Yoshida, M.; Miyado, K.; et al. Seminal vesicle protein SVS2 is required for sperm survival in the uterus. Proc. Natl. Acad. Sci. USA 2014, 111, 4145–4150. [Google Scholar] [CrossRef] [Green Version]
- Dosselli, R.; Grassl, J.; den Boer, S.P.A.; Kratz, M.; Moran, J.M.; Boomsma, J.J.; Baer, B. Protein-level interactions as mediators of sexual conflict in ants. Mol. Cell. Proteom. 2019, 18, S34–S45. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Reproductive Tract Morphology | Sperm Morphology and Semen Traits | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Comparison | Location | Weight | Testis | Seminal Vesicle | Mucus Gland | Length | Count | Volume | Longevity | Citation |
| Apis mellifera jemenitica vs. A. m. carnica | Saudi Arabia | sig | sig | sig | sig | NA | sig | NA | NA | [55] |
| A. m. caucasica vs. hybrid A. m. carnica x caucasica | Poland | NA | NA | NA | NA | nsig | NA | NA | NA | [64] |
| A. m. siciliana vs. A. m. ligustica | Sicily | NA | NA | NA | NA | NA | sig | nsig | sig | [56] |
| A. m. syriaca vs. A. m. ligustica | Jordan | sig | NA | NA | NA | NA | sig | NA | NA | [65] |
| A. m. scutellata x managed vs. managed | Venezuela | sig | NA | nsig | nsig | NA | sig | NA | NA | [57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slater, G.P.; Smith, N.M.A.; Harpur, B.A. Prospects in Connecting Genetic Variation to Variation in Fertility in Male Bees. Genes 2021, 12, 1251. https://doi.org/10.3390/genes12081251
Slater GP, Smith NMA, Harpur BA. Prospects in Connecting Genetic Variation to Variation in Fertility in Male Bees. Genes. 2021; 12(8):1251. https://doi.org/10.3390/genes12081251
Chicago/Turabian StyleSlater, Garett P., Nicholas M. A. Smith, and Brock A. Harpur. 2021. "Prospects in Connecting Genetic Variation to Variation in Fertility in Male Bees" Genes 12, no. 8: 1251. https://doi.org/10.3390/genes12081251
APA StyleSlater, G. P., Smith, N. M. A., & Harpur, B. A. (2021). Prospects in Connecting Genetic Variation to Variation in Fertility in Male Bees. Genes, 12(8), 1251. https://doi.org/10.3390/genes12081251