
Chromosomal Aberrations in Cattle



 ,
,
Abstract
:1. Introduction
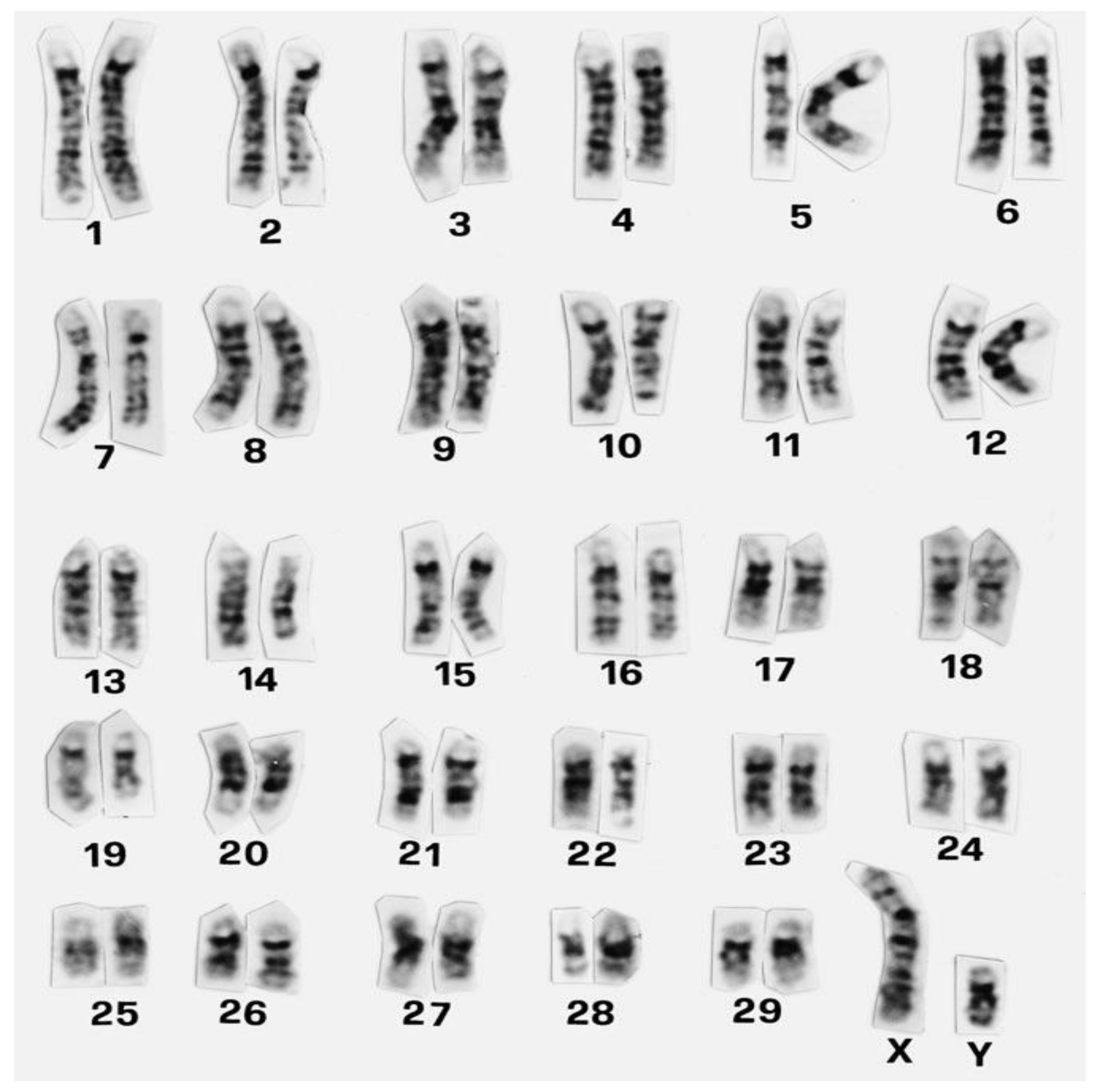
2. Animal Cytogenetics
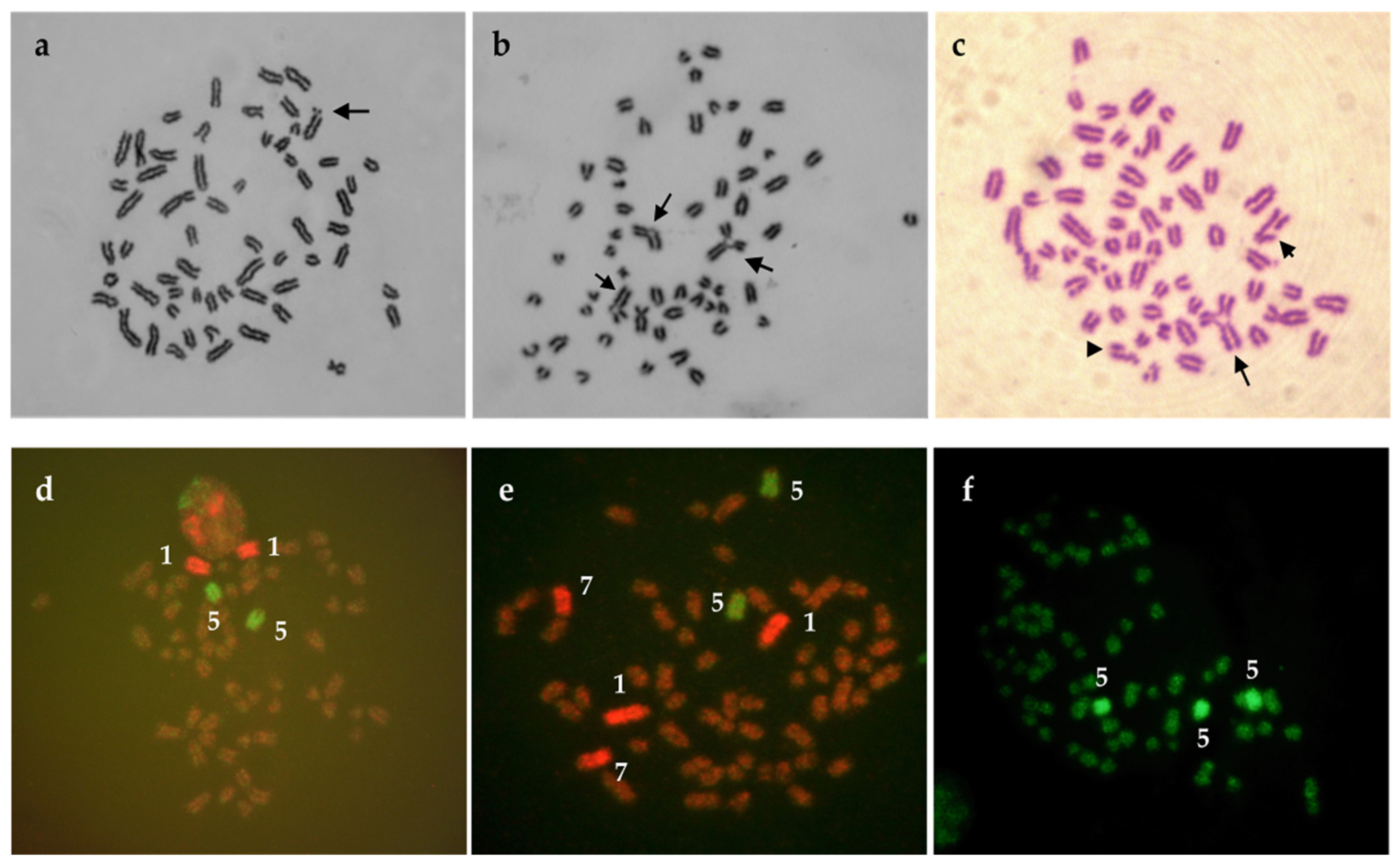
3. Chromosomal Aberrations
3.1. Classification of Chromosomal Abnormalities
3.2. Robertsonian Translocations in Cattle

{kind=link}
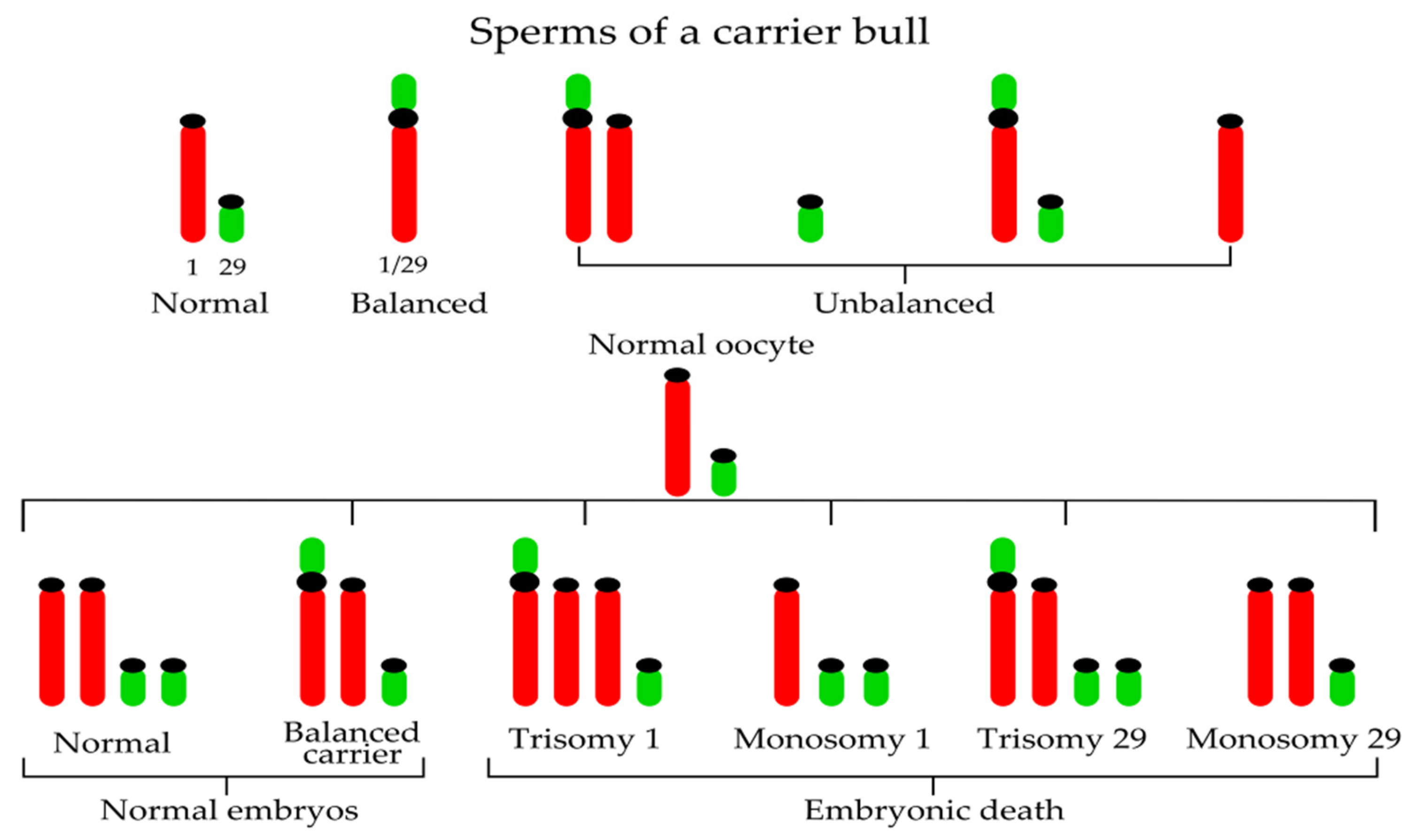{kind=link}
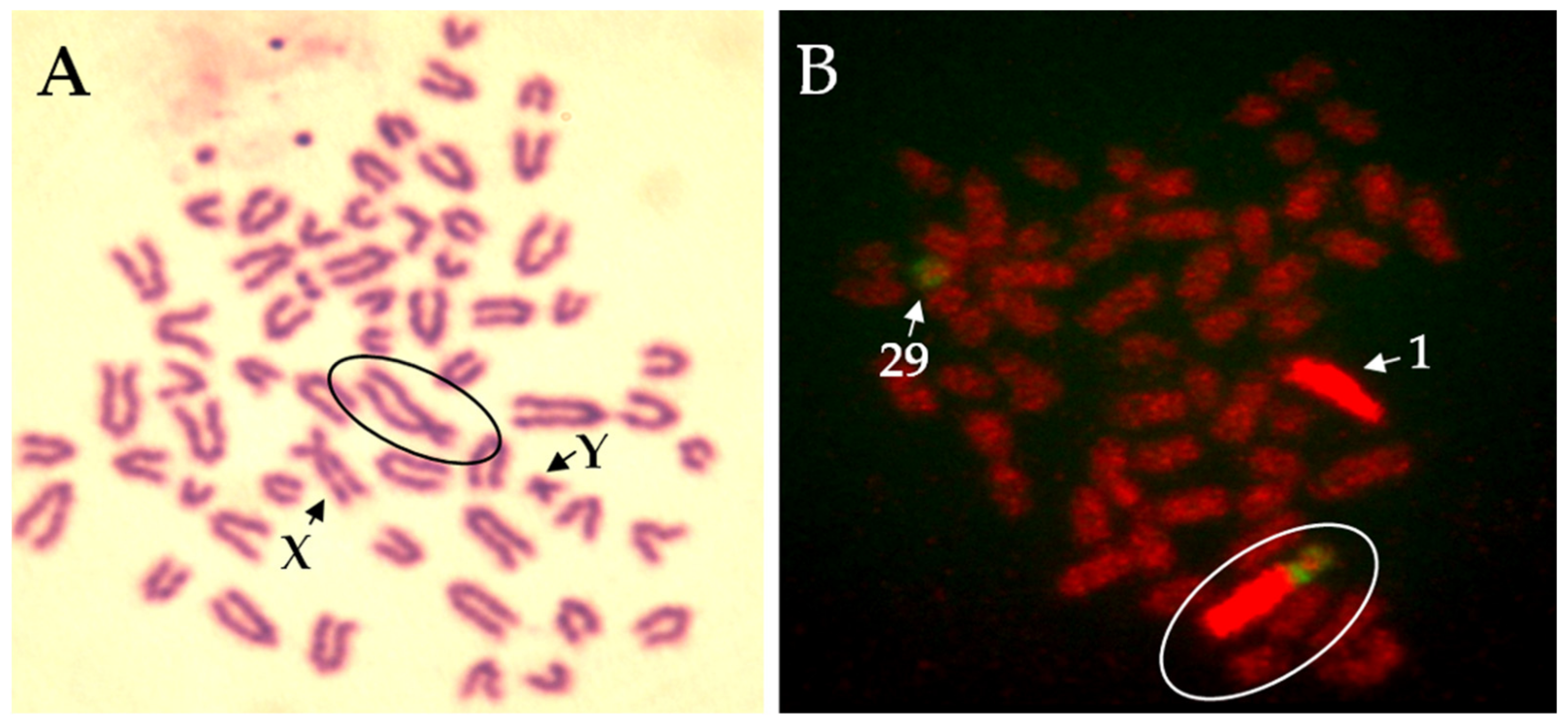{kind=link}
{kind=link}
| Chromosomes Involved in Rob Translocation (Centric Fusion) | Breed of Cattle | Frequency | Year | Reference |
|---|---|---|---|---|
| rob(1;29) | Swedish Red and White cattle | 13−14% | 1964 | [35] |
| Over 50 breeds of cattle | 1964−2014 | [31,32] | ||
| British White | Up to 60% | 1975 | [33] | |
| Corsican | Up to 60% | 1984 | [34] | |
| local Portuguese cattle | above 50% | 2008 | [42] | |
| Maremmana | 18.8% | 2008 | [26] | |
| Romagnola | 13.0% | |||
| Podolian cattle | 11.7% | |||
| Marchigiana | 11.7% | |||
| Chianina | 1.4% | |||
| Limousine | 12.3% | 2008 | [26] | |
| blonde d´Aquitaine | 7.9% | |||
| Charolaise | 1.2% | |||
| Rubia Gallega | 21.9% | 2008 | [26] | |
| Retinta | 16.1% | |||
| Czech Simmental | 27.08% | 2009 | [27] | |
| Andalusian breeds: Negra Andaluza | 19.45% | 2013 | [43] | |
| Berrenda en Negro | 28.9%−32.6% | |||
| Criollo | 12.3% | 2015 | [44] | |
| Swiss American | 7.5% | |||
| Braunvieh (Swiss Brown) | 1.4% | |||
| Holstein | 0.4% | |||
| rob(1;21) rob(23;26) rob(24;26) rob(26;29) | Holstein | 6.4% together | 2016 | [41] |
| rob(3;16) | Montbéliarde | Referred for the first time in one animal (bull) | 2018 | [38] |
| rob(13;23) | Ukrainian Red-and-Motley | 1.9% | 2019 | [45] |
3.3. Reciprocal Translocations in Cattle
3.4. Approaches in Translocations Detection
4. Cattle Chromosomal Aberrations in Genotoxicity Studies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Philips, C.J.C. Principles of Cattle Production, 2nd ed.; CABI Publishing: Cambridge, UK, 2010. [Google Scholar]
- Scheu, A.; Powell, A.; Bollongino, R.; Vigne, J.D.; Tresset, A.; Cakirlar, C.; Benecke, N.; Burger, J. The genetic prehistory of domesticated cattle from their origin to the spread across Europe. BMC Genet. 2015, 16, 54. [Google Scholar] [CrossRef] [Green Version]
- Deb, R.; Singh, U.; Kumar, S.; Sharma, A. Analysis of Cattle Genome. A Tool Manual, 1st ed.; Satish Serial Publishing House: Delhi, India, 2013. [Google Scholar]
- Elsik, C.G.; Tellam, R.L.; Worley, K.C. The genome sequence of Taurine cattle. A window to ruminant biology and evolution. Science 2009, 324, 522–528. [Google Scholar] [PubMed] [Green Version]
- Childers, A.; Reese, J.T.; Sundaram, J.P.; Vile, D.C.; Dickens, C.M.; Childs, K.L.; Salih, H.; Bennett, A.K.; Hagen, D.E.; Adelson, D.L.; et al. Bovine genome database: Integrated tools for genome annotation and discovery. Nucleic Acid Res. 2011, 39, D830–D834. [Google Scholar] [CrossRef]
- Stothard, P.; Liao, X.; Arantes, A.S.; De Pauw, M.; Coros, C.; Plastow, G.S.; Sargolzaei, M.; Crowley, J.J.; Basarab, J.A.; Schenkel, F.; et al. A large and diverse collection of bovine sequences from Canadian Cattle Genome Project. Gigascience 2015, 4, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagen, D.E.; Unni, D.R.; Tayal, A.; Burns, G.W.; Elsik, C.G. Bovine Genome Database: Tools for mining the Bos taurus genome. Methods Mol. Biol. 2018, 1757, 211–249. [Google Scholar] [PubMed]
- Gustavsson, I. From Giessen to Toulouse: 20 years in domestic animal cytogenetics. Genet. Sel. Evol. 1991, 23 (Suppl. S1), 9–17. [Google Scholar] [CrossRef]
- Basrur, P.K.; Stranzinger, G. Veterinary cytogenetics: Past and perspective. Cytogenet. Genome Res. 2008, 120, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Di Berardino, D. Tools of the trade: Diagnostics and research in domestic animal cytogenetics. J. Appl. Genet. 2008, 49, 357–366. [Google Scholar] [CrossRef]
- Cribiu, E.P.; Di Berardino, D.; Di Meo, G.P.; Eggen, A.; Gallagher, D.S.; Gustavsson, I.; Hayes, H.; Iannuzzi, L.; Popescu, C.P.; Rubes, J.; et al. International System for Chromosomal Nomenclature of Domestic Bovids (ISCNDB, 2000). Cytogenet. Cell Genet. 2001, 92, 283–299. [Google Scholar] [CrossRef]
- Rubes, J.; Kubickova, S.; Pagacova, E.; Cernohorska, H.; Di Berardino, D.; Antoninova, M.; Vahala, J.; Robinson, T.J. Phylogenomic study of spiral horned antelope by cross species chromosome painting. Chromosome Res. 2008, 16, 935–947. [Google Scholar] [CrossRef]
- Iannuzzi, L.; King, W.A.; Di Berardino, D. Chromosome evolution in domestic bovids as revealed by chromosome banding and FISH-mapping techniques. Cytogenet. Genome Res. 2009, 126, 49–62. [Google Scholar] [CrossRef]
- Amancio, A.P.; Duarte, S.S.M.; Silva, R.C.; da Cruz, A.S.; Silva, D.C.; da Silva, C.C.; da Cruz, A.D. Banded karyotype of Nelore cattle (Bos taurus indicus Linnaeus, 1758). Comp. Cytogen. 2019, 13, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Raudsepp, T.; Chowdhary, B.P. Chromosome aberrations and fertility disorders in domestic animals. Annu. Rev. Anim. Biosci. 2016, 4, 15–43. [Google Scholar] [CrossRef] [PubMed]
- Szczerbal, I.; Switonski, M. Chromosome Abnormalities in Domestic Animals as Causes of Disorders of Sexual Development or Impaired Fertility. 2016. Available online: https://.intechopen.com/chapters/49736 (accessed on 23 May 2021).
- King, W. Chromosome variation in the embryos of domestic animals. Cytogenet. Genome Res. 2008, 120, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Garrick, D.; Ruvinsky, A. The Genetics of Cattle, 2nd ed.; CABI Publishing: Cambridge, UK, 2015. [Google Scholar]
- Griffiths, A.J.F.; Miller, J.H.; Suzuki, D.T.; Lewontin, R.C.; Gelbart, W.M. Mechanisms of gene imbalance. In An Introduction to Genetic Analysis, 7th ed.; W. H. Freeman: New York, NY, USA, 2000. Available online: https://www.ncbi.nlm.nih.gov/books/NBK21967 (accessed on 8 March 2021).
- Iannuzzi, L.; Di Meo, G.P.; Leifsson, P.S.; Eggen, A.; Christensen, K. A case of trisomy 28 in cattle revealed by both banding and FISH-mapping techniques. Hereditas 2001, 134, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, F.W. Introduction to Veterinary Genetics, 3rd ed.; Wiley-Blackwell: Oxford, UK, 2010. [Google Scholar]
- Häfliger, I.M.; Seefried, F.; Drögemüller, C. Trisomy 29 in a stillborn Swiss Original Braunvieh calf. Anim. Genet. 2020, 51, 483–484. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, A.; Parma, P.; Iannuzzi, L. Chromosome Abnormalities and Fertility in Domestic Bovids: A Review. Animals 2021, 11, 802. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, A.S.; Genualdo, V.; Perucatti, A.; Pauciullo, A.; Varricchio, G.; Incarnato, D. Fatal outcome in a newborn calf associated with partial trisomy 25q and partial monosomy 11q, 60, XX, der (11)t(11;25)(q11;q14 similar to 21). Cytogenet. Genome Res. 2015, 146, 222–229. [Google Scholar] [CrossRef]
- Larkin, D.M.; Farré, M. Cytogenetics and Chromosome Maps. In The Genetics of Cattle, 2nd ed.; Garrick, D., Ruvinsky, A., Eds.; CABI Publishing: Cambridge, UK, 2015; pp. 103–123. [Google Scholar]
- Ducos, A.; Revay, T.; Kovacs, A.; Hidas, A.; Pinton, A.; Bonnet-Garnier, A.; Molteni, L.; Slota, E.; Switonski, M.; Arruga, M.V.; et al. Cytogenetic screening of livestock populations in Europe: An overview. Cytogenet. Genome Res. 2008, 120, 26–41. [Google Scholar] [CrossRef]
- Citek, J.; Rubes, J.; Hajkova, J. Robertsonian translocations, chimerism, and aneuploidy in cattle. J. Dairy Sci. 2009, 92, 3481–3483. [Google Scholar] [CrossRef]
- Uzar, T.; Szczerbal, I.; Serwanska-Leja, K.; Nowacka-Woszuk, J.; Gogulski, M.; Bugaj, S.; Switonski, M.; Komosa, M. Congenital Malformations in a Holstein-Fresian Calf with a Unique Mosaic Karyotype: A Case Report. Animals 2020, 10, 1615. [Google Scholar] [CrossRef] [PubMed]
- Harewood, L.; Fraser, P. The impact of chromosoal rearrangements on regulation of gene expression. Human Molec. Genet. 2014, 23, R76–R82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilch, E.S.; Morton, C.C. Historical and Clinical Perspectives on Chromosomal Translocations. In Chromosome Translocation. Advances in Experimental Medicine and Biology; Zhang, Y., Ed.; Springer: Singapore, 2018; Volume 1044. [Google Scholar]
- Switonski, M. 50 years of studies on bovine 1/29 Robertsonian translocation-from Giemsa staining to genomic analysis. Chromosome Res. 2014, 22, 395. [Google Scholar]
- Popescu, C.P.; Pech, A. Une bibliographie sur la translocation 1/29 de bovins dans le monde (1964–1990). Ann. Zootech. 1991, 40, 271–305. [Google Scholar] [CrossRef]
- Eldridge, F.E. High frequency of a Robertsonian translocation in a herd of British White cattle. Vet. Res. 1975, 97, 71–73. [Google Scholar] [CrossRef]
- Hari, J.J.; Franceschi, P.; Casabianca, F.; Bosher, J.; Popescu, C.P. Etude cytogénétique d’une population de bovins corses. Compte Rendu de l’Academie d’Agriculture de France 1984, 70, 8. [Google Scholar]
- Gustavsson, I.; Rockborn, G. Chromosome abnormality in three cases of lymphatic leukaemia in cattle. Nature 1964, 203, 990. [Google Scholar] [CrossRef] [PubMed]
- Gustavsson, I. Cytogenetics, distribution and phenotypic effects of a translocation in Swedish cattle. Hereditas 1969, 63, 68–169. [Google Scholar] [CrossRef]
- King, A.W. Ingemar Gustavsson (1938-2016). Cytogenet. Genome Res. 2017, 152, 167–168. [Google Scholar]
- Barasc, H.; Mouney-Bonnet, N.; Peigney, C.; Calgaro, A.; Revel, C.; Mary, N.; Ducos, A.; Pinton, A. Analysis of meitoc segregation pattern and interchromosomal effects in a bull heterozygous for a 3/16 Robertsonian translocation. Cytogenet. Genome Res. 2018, 4, 197–203. [Google Scholar] [CrossRef]
- De Lorenzi, L.; Genualdo, V.; Gimelli, S.; Rossi, E.; Perucatti, A.; Iannuzzi, A.; Zannotti, M.; Malagutti, L.; Molteni, L.; Iannuzzi, L.; et al. Genomic analysis of cattle rob (1;29). Chromosome Res. 2012, 20, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Joerg, H.; Garner, D.; Rieder, S.; Suwattana, D.; Stransinger, G. Molecular genetic characterization of Robertsonian translocations in cattle. J. Anim. Breed. Genet. 2001, 118, 371–377. [Google Scholar] [CrossRef]
- Arslan, K.; Ozdemir, F.; Ilgar, E.G.; Akyuz, B. Cytogenetic and molecular genetic screening in Holstein cattle breed which showing repeat breeding problems. J. Agric. Sci. Tarim Bilimleri Dergisi 2016, 22, 370–373. [Google Scholar]
- Iannuzzi, A.; Di Meo, G.P.; Caputi Jambrenghi, A.; Vonghia, G.; Iannuzzi, L.; Rangel-Figueiredo, F. Frequency and distribution of rob (1;29) in eight Portugese cattle breeds. Cytogenet. Genome Res. 2008, 120, 147–149. [Google Scholar] [CrossRef] [PubMed]
- Rodero-Serrano, E.; Demyda-Peyrás, S.; González-Martinez, A.; Rodero-Franganillo, A.; Moreno-Millán, M. The rob (1;29) chromosome translocation in endangered Andalusian cattle breeds. Livest. Sci. 2013, 158, 32–39. [Google Scholar] [CrossRef]
- Meza-López, C.; Lozano-Carbajal, B.; de la Colina-Flores, F.; Bañuelos-Valenzuela, R.; López-Carlos, M.A.; Echavarría-Cháirez, F.G.; Reveles-Torres, L.R. Prevalence of the Robertsonian (1;29) translocation in cattle in the state of Zacatecas, México. Rev. Mex. Cienc. Pecu 2015, 6, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Dzitsiuk, V.V.; Tipilo, H.T. Chromosomal anomalies in dairy cattle as reasons of impaired fertility. Agric. Sci. Pract. 2019, 6, 60. [Google Scholar] [CrossRef]
- Gholap, P.N.; Kale, D.S.; Sirothia, A.R. Genetic diseases in cattle: A review. Res. J. Anim. Vet. Fish. Sci. 2014, 2, 24–33. [Google Scholar]
- De Lorenzi, L.; De Giovanni, A.; Molteni, L.; Denis, C.; Eggen, A.; Parma, P. Characterization of balanced reciprocal translocation, rcp(9;11)(q27;q11) in cattle. Cytogenet. Genome Res. 2007, 119, 231–234. [Google Scholar] [CrossRef]
- De Lorenzi, L.; Morando, P.; Planas, J.; Zannotti, M.; Molteni, L.; Parma, P. Reciprocal translocations in cattle: Frequencies estimation. J. Anim. Breed. Genet. 2012, 129, 409–416. [Google Scholar] [CrossRef] [Green Version]
- De Lorenzi, L.; Kopecna, O.; Gimelli, S.; Cernohorska, H.; Zannotti, M.; Béna, F.; Molteni, L.; Rubes, J.; Parma, P. Reciprocal translocation t(4;7)(q14;q28) in cattle: Molecular characterization. Cytogenet. Genome Res. 2010, 129, 298–304. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzi, L.; Rossi, E.; Gimelli, S.; Parma, P. De novo reciprocal translocation t(5;6)(q13;q34) in cattle: Cytogenetic and molecular characterization. Cytogenet. Genome Res. 2014, 142, 95–100. [Google Scholar]
- De Lorenzi, L.; Iannuzzi, A.; Rossi, E.; Bonacina, S.; Parma, P. Centromere repositioning in cattle (Bos taurus) chromosome 17. Cytogenet. Genome Res. 2017, 151, 191–197. [Google Scholar] [CrossRef]
- Jennings, R.L.; Griffin, D.J.; O′Connor, R.E. A new approach for accurate detection of chromosome rearrangements that affect fertility in cattle. Animals 2020, 10, 114. [Google Scholar] [CrossRef] [Green Version]
- Switonski, M.; Andersson, M.; Nowacka-Woszuk, J.; Szczerbal, I.; Sosnowski, J.; Kopp, C.; Cernohorska, H.; Rubeš, J. Identification of a new reciprocal translocation in an AI bull by synaptonemal complex analysis, followed by chromosome painting. Cytogenet. Genome Res. 2008, 121, 245–248. [Google Scholar] [CrossRef]
- Rubes, J.; Pinton, A.; Bonnet-Garnier, A.; Fillon, V.; Musilova, P.; Michalova, K.; Kubickova, S.; Ducos, A.; Yerle, M. Fluorscence in situ hybridization applied to domestic animal cytogenetics. Cytogenet. Genome Res. 2009, 126, 34–48. [Google Scholar]
- Yang, F.; Trifonov, V.; Ng, B.L.; Kosyakova, N.; Carter, N.P. Generation of paint probes by flow-sorted and microdissected chromosomes. In Fluorescence in situ hybridization (FISH). Application Guide, 1st ed.; Liehr, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 35–52. [Google Scholar]
- Dixon, S.C.; Miller, N.G.; Carter, N.P.; Tucker, E.M. Bivariate flow cytometry of farm animal chromosomes: A potential tool for gene mapping. Anim. Genet. 1992, 23, 203–210. [Google Scholar] [CrossRef]
- Kubickova, S.; Cernohorska, H.; Musilova, P.; Rubes, J. The use of laser microdissection for the preparation of chromosome-specific painting probes in farm animals. Chromosome Res. 2002, 10, 571–577. [Google Scholar]
- Fröhlich, J.; Kubickova, S.; Musilová, P.; Cernohorska, H.; Muskova, H.; Vodička, R.; Rubes, J. Karyotype relationships among selected deer species and cattle revealed by bovine FISH probes. PLoS ONE 2017, 12, e0187559. [Google Scholar] [CrossRef] [Green Version]
- Rezacova, O.; Kubickova, S.; Cernohorska, H.; Rubes, J. Comparison of spontaneous background genomic aberration frequencies among cattle, pig and humans using dual-coloured FISH. Chromosome Res. 2003, 11, 715–724. [Google Scholar]
- Pinton, A.; Ducos, A.; Yerle, M. Chromosomal rearrangements in cattle and pigs revealed by chromosome microdissection and chromosome painting. Genet. Sel. Evol. 2003, 35, 685–696. [Google Scholar] [CrossRef] [Green Version]
- De Lorenzi, L.; Molteni, L.; De Giovanni, A.; Parma, P. A new case of rob (14, 17) in cattle. Cytogenet. Genome Res. 2008, 120, 144–146. [Google Scholar] [CrossRef]
- De Lorenzi, L.; Molteni, L.; Denis, C.; Eggen, A.; Parma, P. A new case of centric fusion in cattle:rob (21;23). Anim. Genet. 2008, 39, 454–455. [Google Scholar] [CrossRef]
- Weise, A.; Mrasek, K.; Kosyakova, N.; Mkrtchyan, H.; Gross, M.; Klaschka, V.; Liehr, T. ISH probes derived from BACs, including microwave treatment for better FISH results. In Fluorescence in situ hybridization (FISH). Application Guide, 1st ed.; Liehr, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 53–60. [Google Scholar]
- Eggen, A.; Gautier, M.; Billaut, A.; Petit, E.; Hayes, H.; Laurent, P.; Urban, C. Construction and characterization of a bovine BAC library with four genome-equivalent coverage. Genet. Sel. Evol. 2001, 33, 543–548. [Google Scholar] [CrossRef]
- Huang, L.; Ma, F.; Chapman, A.; Lu, S.; Xie, X.S. Single-cell whole genome amplification and sequencing: Methodology and applications. Annu. Rev. Genomics Hum. Genet. 2015, 16, 79–102. [Google Scholar] [CrossRef] [Green Version]
- Dimitriadou, E.; Esteki, M.Z.; Vermeesch, J.R. Copy Number Variation Analysis by Array Analysis of Single Cells Following Whole Genome Amplification. In Whole Genome Amplification, Methods and Protocols; Kroneis, T., Ed.; Humana Press: Totowa, NJ, USA, 2015. [Google Scholar]
- Hawken, R.J.; Cavanagh, J.A.L.; Meadows, J.R.S.; Khatkar, M.S.; Husaini, Y.; Zenger, K.R.; McClintock, S.; McClintock, A.E.; Raadsma, H.W. Technical note: Whole-genome amplification of DNA extracted from cattle semen samples. J. Dairy Sci. 2006, 89, 2217–2222. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, D.; Zhang, J.; Mao, Y.; Kong, L.; Zhang, Y.; Liang, B.; Sun, X.; Xu, C. BasePhasing: A highly efficient approach preimplantation genetic haplotyping in clinical application of balanced translocation carriers. BMC Med. Genom. 2019, 12, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubeš, J.; Borkovec, L.; Hořínová, Z.; Urbanová, J.; Proroková, I.; Kulíková, L. Cytogenetic monitoring of farm animals under conditions of environmental pollution. Mutat. Res. 1992, 283, 199–210. [Google Scholar] [CrossRef]
- Oruc, H.H. Fungicides and their effects on animals. In Fungicides; Carisse, O., Ed.; InTech: Rijeka, Croatia, 2010. [Google Scholar]
- Guitart, R.; Croubels, S.; Caloni, F.; Sachana, M.; Davanzo, F.; Vandenbroucke, V.; Berny, P. Animal Poisoning in Europe Part I: Farm livestock and poultry. Vet. J. 2010, 183, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.F.; Gil, F.; Lacasana, M. Toxicological interactions of pesticide mixtures: An update. Arch. Toxicol. 2017, 91, 3211–3223. [Google Scholar] [CrossRef] [PubMed]
- Rubes, J.; Pokorna, Z.; Borkovec, L.; Urbanova, J.; Strnadova, V. Dairy cattle as a bioindicator of exposure to genotoxic substances in a heavy polluted area in northern Bohemia. Mutat. Res. 1997, 391, 57–70. [Google Scholar] [CrossRef]
- Favetta, L.; Villagómez, D.; Iannuzzi, L.; Di Meo, G.; Webb, A.; Crain, S.; King, W. Disorders of sexual development and abnormal early development in domestic food-producing mammals: The role of chromosome abnormalities, environment and stress factors. Sex Dev. 2012, 6, 18–32. [Google Scholar] [CrossRef]
- Nakamura, A.J.; Suzuki, M.; Redon, C.E.; Kuwahara, Y.; Yamashiro, H.; Abe, Y.; Takahashi, S.; Fukuda, T.; Isogai, E.; Bonner, W.M.; et al. The Causal Relationship between DNA Damage Induction in Bovine Lymphocytes and the Fukushima Nuclear Power Plant Accident. Radiat. Res. 2017, 187, 630–636. [Google Scholar] [CrossRef] [Green Version]
- Lioi, M.; Scarfi, M.; Santoro, A.; Barbieri, R.; Zeni, O.; Di Berardino, D.; Ursini, M. Genotoxicity and oxidative stress induced by pesticide exposure in bovine lymphocyte cultures in vitro. Mutat. Res. 1998, 403, 13–20. [Google Scholar] [CrossRef]
- Rossi, C.; Danielli, P.P.; Ronchi, B. In vitro effects of the organochlorine pesticide β-hexachlorocyclohexane on bovine peripheral blood mononuclear cells. Ital. J. Anim. Sci. 2014, 13, 620–626. [Google Scholar] [CrossRef]
- Chwarzbacherová, V.; Wnuk, M.; Lewinska, A.; Potocki, L.; Zebrowski, J.; Koziorowski, M.; Holečková, B.; Šiviková, K.; Dianovský, J. Evaluation of cytotoxic and genotoxic activity of fungicide formulation Tango Super in bovine lymphocytes. Environ. Poll. 2017, 220, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Sivikova, K.; Holeckova, B.; Schwarzbacherova, V.; Galdikova, M.; Dianovsky, J. Potential chromosome damage, cell-cycle kinetics and apoptosis induced by epoxiconazole in bovine peripheral lymphocytes in vitro. Chemosphere 2018, 193, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Ferré, D.M.; Ludueña, H.R.; Romano, R.R.; Gorla, N.B.M. Evaluation of the genotoxic potential of cypermetrin, chlorpyrifos and their subsequent mixture, on cultured bovine lymphocytes. Chemosphere 2020, 243, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Holeckova, B.; Sivikova, K.; Dianovsky, J.; Galdikova, M. Efect of triazole pesticide formulation on bovine culture cells. J. Environ. Sci. Health B 2013, 48, 1080–1088. [Google Scholar] [CrossRef] [PubMed]
- Šiviková, K.; Dianovský, J.; Holečková, B.; Galdíková, M.; Kolesárová, V. Assessment of cytogenetic damage in bovine peripheral lymphocytes exposed to in vitro tebuconazole-based fungicide. Chemosphere 2013, 92, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Castro, C.C.; Luoma, A.M.; Adams, E.J. Coevolution of T cell receptors with MHC and non-MHC ligands. Immunol. Rev. 2015, 267, 30–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drazovska, M.; Sivikova, K.; Holeckova, B.; Dianovsky, J.; Galdikova, M.; Schwarzbacherova, V. Evaluation of potential genotoxic/cytotoxic effects induced by epoxiconazole and fenpropimorph-based fungicide in bovine lymphocytes in vitro. J. Environ. Sci. Health B 2016, 51, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Galdikova, M.; Sivikova, K.; Holeckova, B.; Dianovsky, J.; Drazovska, M.; Schwarzbacherova, V. The effect of thiacloprid formulation on DNA/chromosome damage and changes in GST activity in bovine peripheral lymphocytes. J. Environ. Sci. Health B 2016, 50, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Marshall, R.; Obe, G. Application of chromosome painting to clastogenicity testing in vitro. Environ. Mol. Mutagen. 1998, 32, 212–222. [Google Scholar] [CrossRef]
- Baumgartner, A. Comparative genomic hybridization (CGH) in genotoxicology. In Genotoxicity Assessment. Methods and Protocols; Dhawan, A., Bajpajee, M., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 245–256. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holečková, B.; Schwarzbacherová, V.; Galdíková, M.; Koleničová, S.; Halušková, J.; Staničová, J.; Verebová, V.; Jutková, A. Chromosomal Aberrations in Cattle. Genes 2021, 12, 1330. https://doi.org/10.3390/genes12091330
Holečková B, Schwarzbacherová V, Galdíková M, Koleničová S, Halušková J, Staničová J, Verebová V, Jutková A. Chromosomal Aberrations in Cattle. Genes. 2021; 12(9):1330. https://doi.org/10.3390/genes12091330
Chicago/Turabian StyleHolečková, Beáta, Viera Schwarzbacherová, Martina Galdíková, Simona Koleničová, Jana Halušková, Jana Staničová, Valéria Verebová, and Annamária Jutková. 2021. "Chromosomal Aberrations in Cattle" Genes 12, no. 9: 1330. https://doi.org/10.3390/genes12091330
APA StyleHolečková, B., Schwarzbacherová, V., Galdíková, M., Koleničová, S., Halušková, J., Staničová, J., Verebová, V., & Jutková, A. (2021). Chromosomal Aberrations in Cattle. Genes, 12(9), 1330. https://doi.org/10.3390/genes12091330