
Calcineurin Gamma Catalytic Subunit PPP3CC Inhibition by miR-200c-3p Affects Apoptosis in Epithelial Ovarian Cancer

 ,
,  ,
,  ,
,  ,
,  , , ,
, , ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Cell Lines
2.2. MiR-200c-3p Expression in Public Databases and mRNA in EOC Patients through CCLE and TCGA
2.3. RNA Extraction and RT-qPCR
2.4. Oligonucleotide Transfections
2.5. Western Blot Analysis
2.6. Clonogenic and Migration Assays
2.7. Apoptosis Assay
2.8. Statistical Analysis
3. Results
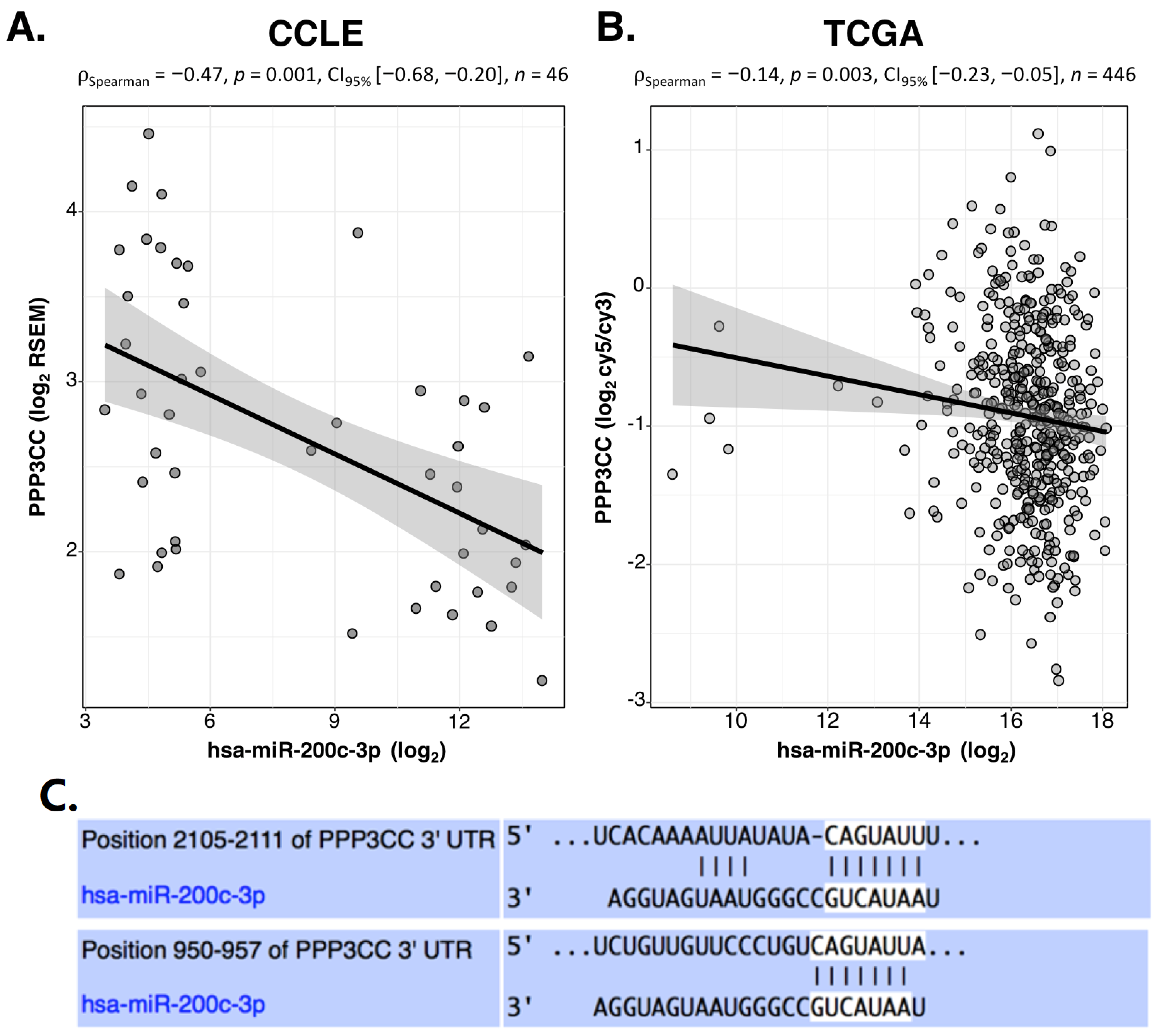
3.1. Inverse Correlation Analysis of miR-200c-3p and mRNAs in OC Cell Lines by Mining CCLE and TCGA Data
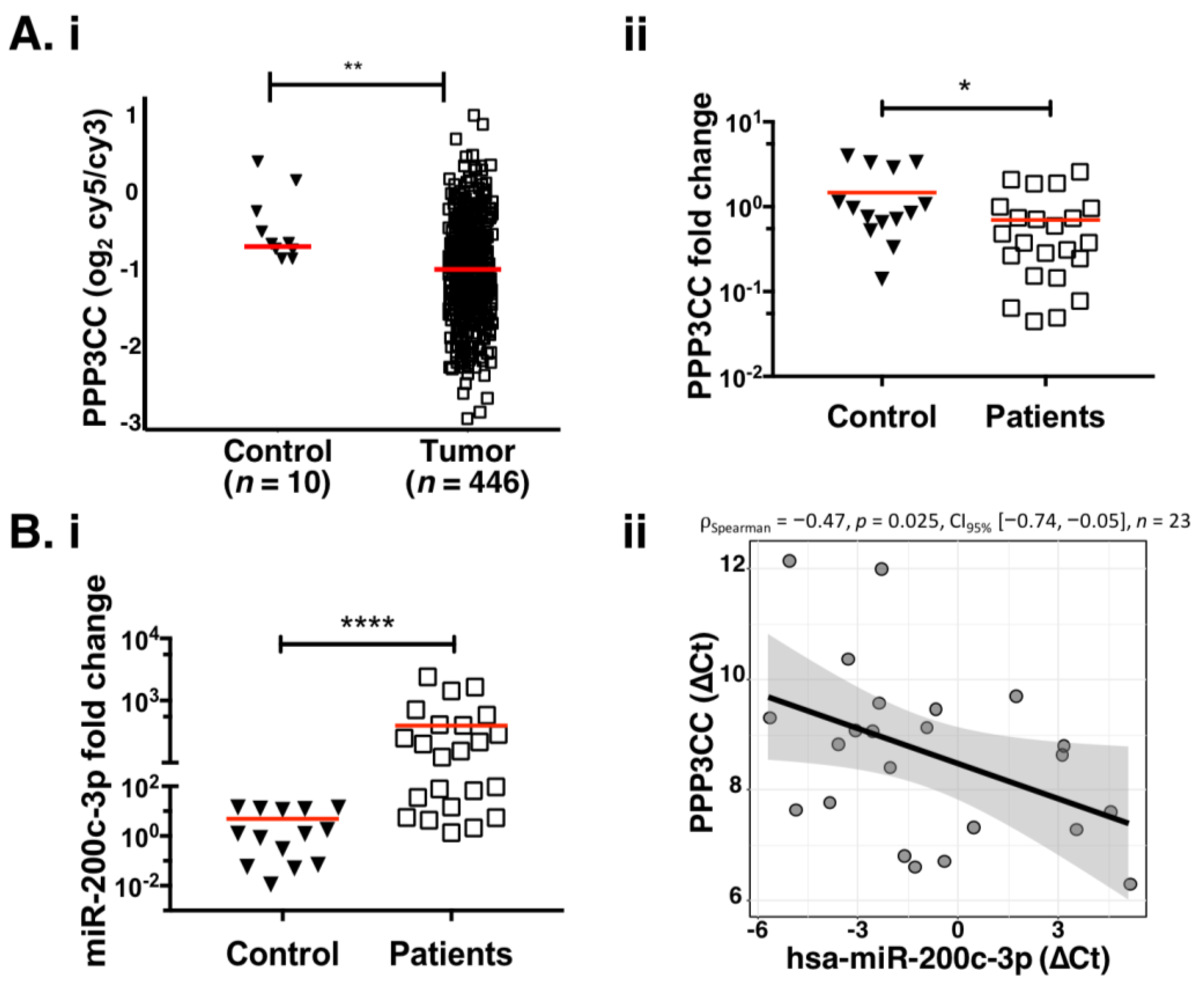
3.2. MiR-200c-3p Is Inversely Correlated with PPP3CC in EOC Patients
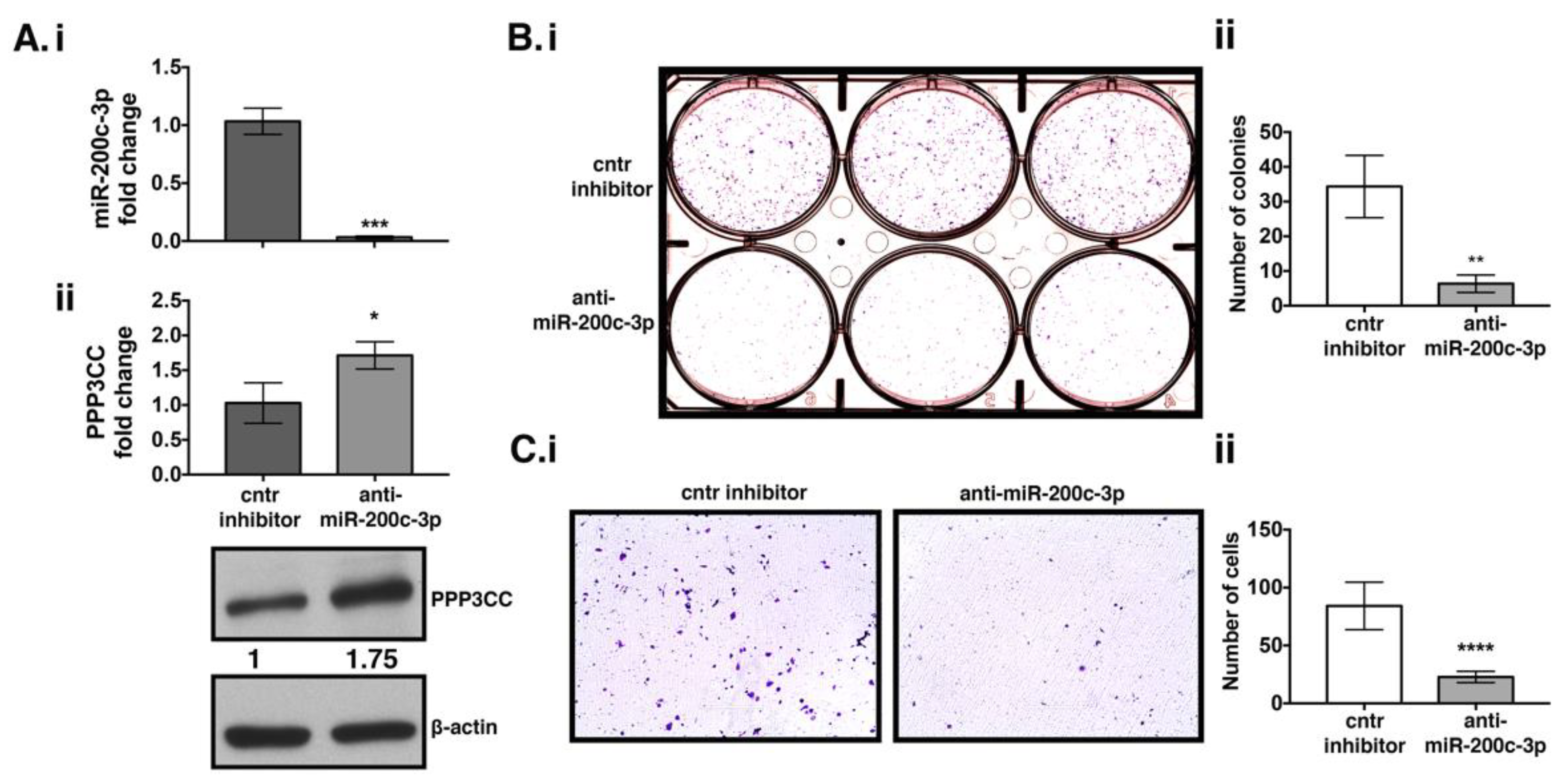
3.3. MiR-200c-3p Downregulation Decreases Proliferation and Migration in EOC Cells
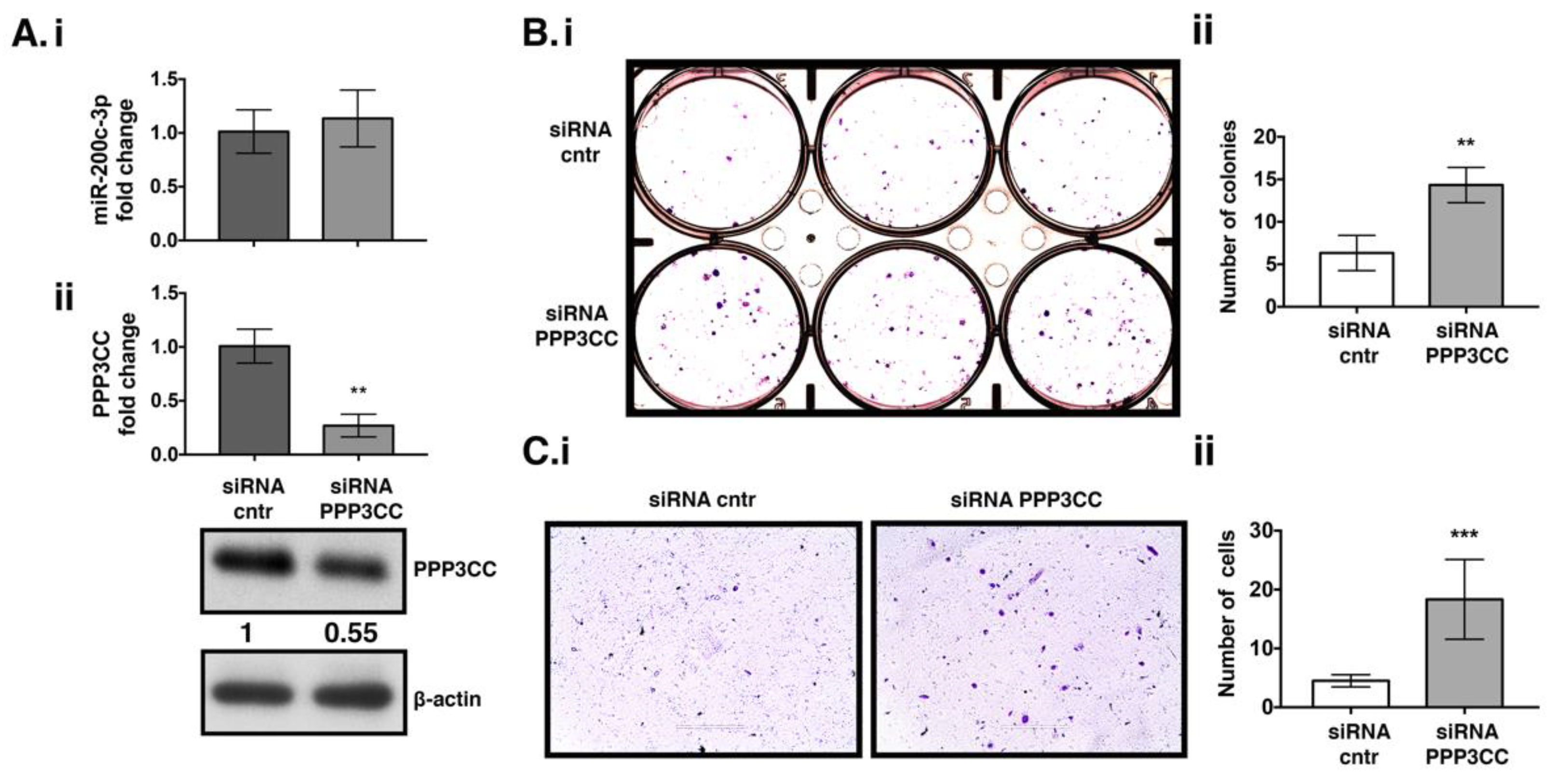
3.4. PPP3CC Knock Down in UWB 1.289 + BRCA1 Cell Line Induced Proliferation and Metastasis
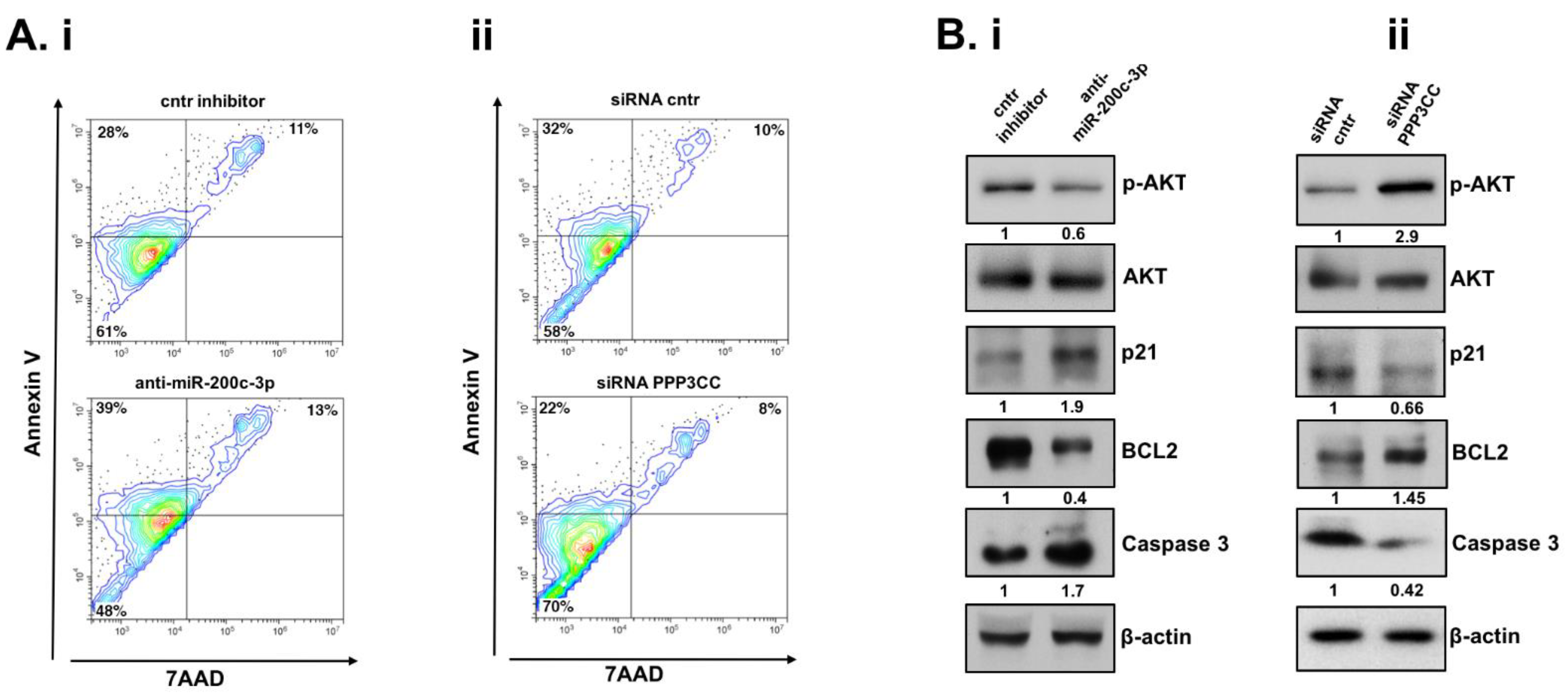
3.5. Inhibition of miR-200c-3p and PPP3CC, in EOC Transfected Cells, Has Opposite Effects in Apoptosis and AKT Signaling Pathway in EOC Transfected Cells
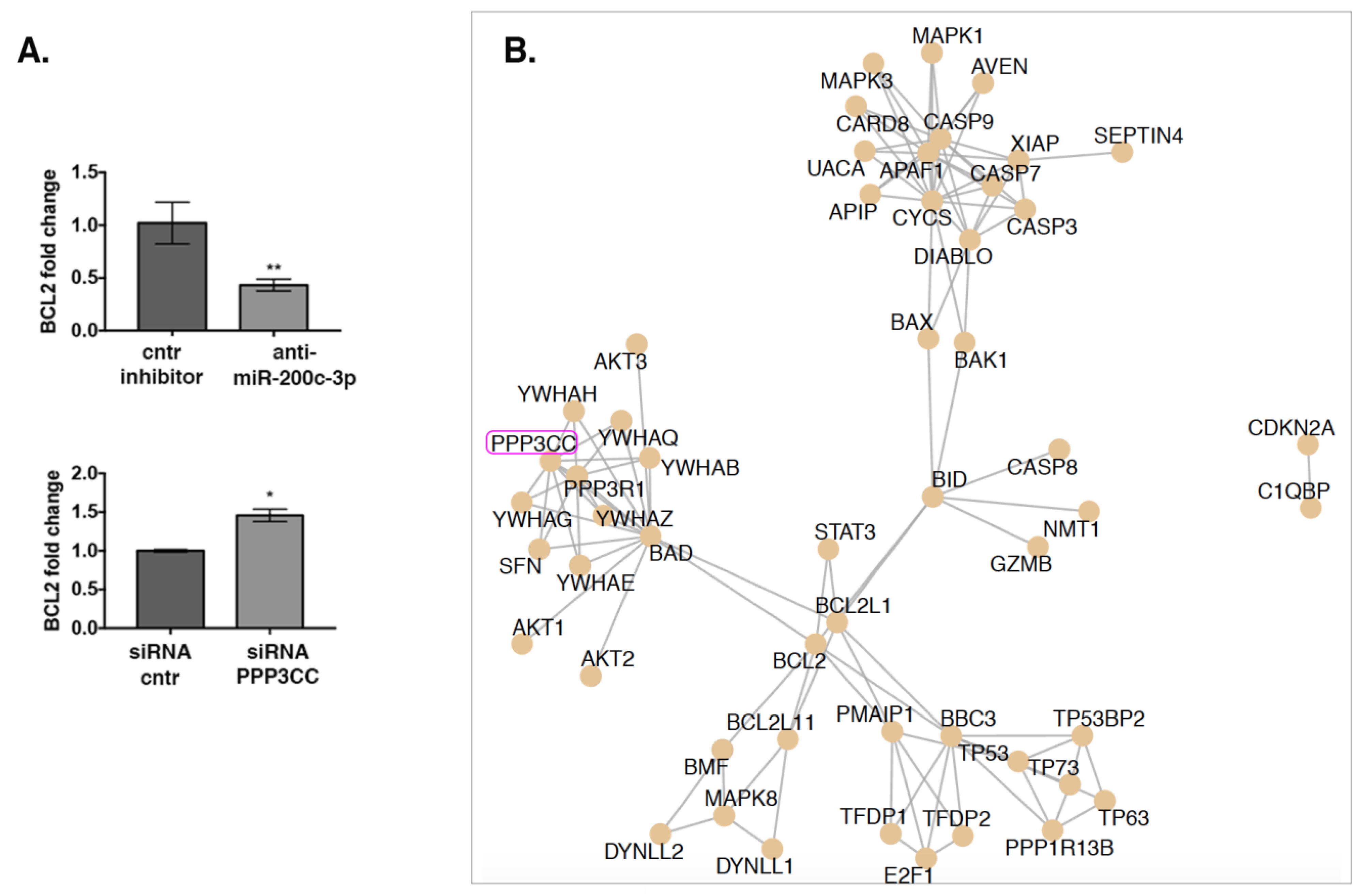
3.6. Network Analysis and Experimental Validation Reveals a Downstream Regulatory Activity of PPP3CC on Apoptosis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lheureux, S.; Braunstein, M.; Oza, A.M. Epithelial ovarian cancer: Evolution of management in the era of precision medicine. CA A Cancer J. Clin. 2019, 69, 280–304. [Google Scholar] [CrossRef] [Green Version]
- Torre, L.A.; Trabert, B.; DeSantis, C.E.; Miller, K.D.; Samimi, G.; Runowicz, C.D.; Gaudet, M.M.; Jemal, A.; Siegel, R.L. Ovarian cancer statistics. CA Cancer J. Clin. 2018, 68, 284–296. [Google Scholar] [CrossRef]
- Vescarelli, E.; Gerini, G.; Megiorni, F.; Anastasiadou, E.; Pontecorvi, P.; Solito, L.; De Vitis, C.; Camero, S.; Marchetti, C.; Mancini, R.; et al. MiR-200c sensitizes Olaparib-resistant ovarian cancer cells by targeting Neuropilin. J. Exp. Clin. Cancer Res. 2020, 39, 3. [Google Scholar] [CrossRef]
- Anastasiadou, E.; Messina, E.; Sanavia, T.; Mundo, L.; Farinella, F.; Lazzi, S.; Megiorni, F.; Ceccarelli, S.; Pontecorvi, P.; Marampon, F.; et al. MiR-200c-3p Contrasts PD-L1 Induction by Combinatorial Therapies and Slows Proliferation of Epithelial Ovarian Cancer through Downregulation of β-Catenin and c-Myc. Cells 2021, 10, 519. [Google Scholar] [CrossRef]
- Megiorni, F.; Camero, S.; Pontecorvi, P.; Camicia, L.; Marampon, F.; Ceccarelli, S.; Anastasiadou, E.; Bernabò, N.; Perniola, G.; Pizzuti, A.; et al. OTX015 Epi-Drug Exerts Antitumor Effects in Ovarian Cancer Cells by Blocking GNL3-Mediated Radioresistance Mechanisms: Cellular, Molecular and Computational Evidence. Cancers 2021, 13, 1519. [Google Scholar] [CrossRef]
- Lisio, M.A.; Fu, L.; Goyeneche, A.; Gao, Z.H.; Telleria, C. High-Grade Serous Ovarian Cancer: Basic Sciences, Clinical and Therapeutic Standpoints. Int. J. Mol. Sci. 2019, 20, 952. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [Green Version]
- Anastasiadou, E.; Faggioni, A.; Trivedi, P.; Slack, F.J. The Nefarious Nexus of Noncoding RNAs in Cancer. Int. J. Mol. Sci. 2018, 19, 2072. [Google Scholar] [CrossRef] [Green Version]
- Staicu, C.E.; Predescu, D.V.; Rusu, C.M.; Radu, B.M.; Cretoiu, D.; Suciu, N.; Crețoiu, S.M.; Voinea, S.C. Role of microRNAs as Clinical Cancer Biomarkers for Ovarian Cancer: A Short Overview. Cells 2020, 9, 169. [Google Scholar] [CrossRef] [Green Version]
- Zuberi, M.; Mir, R.; Das, J.; Ahmad, I.; Javid, J.; Yadav, P.; Masroor, M.; Ahmad, S.; Ray, P.C.; Saxena, A. Expression of serum miR-200a, miR-200b, and miR-200c as candidate biomarkers in epithelial ovarian cancer and their association with clinicopathological features. Clin. Transl. Oncol. 2015, 17, 779–787. [Google Scholar] [CrossRef]
- Meng, X.; Müller, V.; Milde-Langosch, K.; Trillsch, F.; Pantel, K.; Schwarzenbach, H. Diagnostic and prognostic relevance of circulating exosomal miR-373, miR-200a, miR-200b and miR-200c in patients with epithelial ovarian cancer. Oncotarget 2016, 7, 16923–16935. [Google Scholar] [CrossRef] [Green Version]
- Sulaiman, S.A.; Ab Mutalib, N.-S.; Jamal, R. miR-200c Regulation of Metastases in Ovarian Cancer: Potential Role in Epithelial and Mesenchymal Transition. Front. Pharmacol. 2016, 7, 271. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Rao, A.; Hogan, P.G. Interaction of calcineurin with substrates and targeting proteins. Trends Cell Biol. 2011, 21, 91–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, J.; Cyert, M.S. Identifying New Substrates and Functions for an Old Enzyme: Calcineurin. Cold Spring Harb. Perspect. Biol. 2020, 12, a035436. [Google Scholar] [CrossRef] [PubMed]
- Xin, B.; Ji, K.Q.; Liu, Y.S.; Zhao, X.D. Higher expression of calcineurin predicts poor prognosis in unique subtype of ovarian cancer. J. Ovarian Res. 2019, 12, 75. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, W.; Xu, W.; Du, M.; Ma, G.; Liu, H.; Chu, H.; Tong, N.A.; Wang, M.; Shao, J.; et al. Genetic Variations in the 3′-untranslated Regions of Genes Involved in the Cell Cycle and Apoptosis Pathways Affect Bladder Cancer Risk. Cancer Genom. Proteom. 2018, 15, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Hornstein, M.; Hoffmann, M.J.; Alexa, A.; Yamanaka, M.; Müller, M.; Jung, V.; Rahnenführer, J.; Schulz, W.A. Protein phosphatase and TRAIL receptor genes as new candidate tumor genes on chromosome 8p in prostate cancer. Cancer Genom. Proteom. 2008, 5, 123–136. [Google Scholar]
- DelloRusso, C.; Welcsh, P.L.; Wang, W.; Garcia, R.L.; King, M.C.; Swisher, E.M. Functional characterization of a novel BRCA1-null ovarian cancer cell line in response to ionizing radiation. Mol. Cancer Res. 2007, 5, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Ghandi, M.; Huang, F.W.; Jané-Valbuena, J.; Kryukov, G.V.; Lo, C.C.; McDonald, E.R., 3rd; Barretina, J.; Gelfand, E.T.; Bielski, C.M.; Li, H.; et al. Next-generation characterization of the Cancer Cell Line Encyclopedia. Nature 2019, 569, 503–508. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Vescarelli, E.; Pilloni, A.; Dominici, F.; Pontecorvi, P.; Angeloni, A.; Polimeni, A.; Ceccarelli, S.; Marchese, C. Autophagy activation is required for myofibroblast differentiation during healing of oral mucosa. J. Clin. Periodontol. 2017, 44, 1039–1050. [Google Scholar] [CrossRef]
- Jeong, J.H.; Park, S.J.; Dickinson, S.I.; Luo, J.L. A Constitutive Intrinsic Inflammatory Signaling Circuit Composed of miR-196b, Meis2, PPP3CC, and p65 Drives Prostate Cancer Castration Resistance. Mol. Cell 2017, 65, 154–167. [Google Scholar] [CrossRef] [Green Version]
- Bellacosa, A.; Kumar, C.C.; Di Cristofano, A.; Testa, J.R. Activation of AKT kinases in cancer: Implications for therapeutic targeting. Adv. Cancer Res. 2005, 94, 29–86. [Google Scholar] [CrossRef]
- Jänicke, R.U.; Sohn, D.; Essmann, F.; Schulze-Osthoff, K. The multiple battles fought by anti-apoptotic p21. Cell Cycle 2007, 6, 407–413. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; He, Q.Y. ReactomePA: An R/Bioconductor package for reactome pathway analysis and visualization. Mol. Biosyst. 2016, 12, 477–479. [Google Scholar] [CrossRef]
- Cittelly, D.M.; Dimitrova, I.; Howe, E.N.; Cochrane, D.R.; Jean, A.; Spoelstra, N.S.; Post, M.D.; Lu, X.; Broaddus, R.R.; Spillman, M.A.; et al. Restoration of miR-200c to ovarian cancer reduces tumor burden and increases sensitivity to paclitaxel. Mol. Cancer Ther. 2012, 11, 2556–2565. [Google Scholar] [CrossRef] [Green Version]
- Cao, Q.; Lu, K.; Dai, S.; Hu, Y.; Fan, W. Clinicopathological and prognostic implications of the miR-200 family in patients with epithelial ovarian cancer. Int. J. Clin. Exp. Pathol. 2014, 7, 2392–2401. [Google Scholar]
- Kim, Y.W.; Kim, E.Y.; Jeon, D.; Liu, J.L.; Kim, H.S.; Choi, J.W.; Ahn, W.S. Differential microRNA expression signatures and cell type-specific association with Taxol resistance in ovarian cancer cells. Drug Des. Dev. Ther. 2014, 8, 293–314. [Google Scholar] [CrossRef] [Green Version]
- Pekarsky, Y.; Croce, C.M. Is miR-29 an oncogene or tumor suppressor in CLL? Oncotarget 2010, 1, 224–227. [Google Scholar] [CrossRef]
- Croce, C.M. Causes and consequences of microRNA dysregulation in cancer. Nat. Rev. Genet. 2009, 10, 704–714. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, S.; Chen, B.; Fu, C.; Dang, Y.; Fang, P.; Wang, J.; Wang, N.; Liu, L. Repression of the expression of PPP3CC by ZEB1 confers activation of NF-κB and contributes to invasion and growth in glioma cells. Jpn. J. Clin. Oncol. 2018, 48, 175–183. [Google Scholar] [CrossRef]
- Quang, C.T.; Leboucher, S.; Passaro, D.; Fuhrmann, L.; Nourieh, M.; Vincent-Salomon, A.; Ghysdael, J. The calcineurin/NFAT pathway is activated in diagnostic breast cancer cases and is essential to survival and metastasis of mammary cancer cells. Cell Death Dis. 2015, 6, e1658. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Kong, B.; Yang, X.; Cui, B.; Wei, Y.; Yang, Q. Overexpression of CHP2 enhances tumor cell growth, invasion and metastasis in ovarian cancer. In Vivo 2007, 21, 593–598. [Google Scholar]
- Hamano, R.; Miyata, H.; Yamasaki, M.; Kurokawa, Y.; Hara, J.; Moon, J.H.; Nakajima, K.; Takiguchi, S.; Fujiwara, Y.; Mori, M.; et al. Overexpression of miR-200c induces chemoresistance in esophageal cancers mediated through activation of the Akt signaling pathway. Clin. Cancer Res. 2011, 17, 3029–3038. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Dong, Z.; Chen, Y.; Yang, L.; Lai, D. Enrichment of ovarian cancer stem-like cells is associated with epithelial to mesenchymal transition through an miRNA-activated AKT pathway. Cell Prolif. 2013, 46, 436–446. [Google Scholar] [CrossRef]
- Privat, M.; Aubel, C.; Arnould, S.; Communal, Y.; Ferrara, M.; Bignon, Y.J. AKT and p21 WAF1/CIP1 as potential genistein targets in BRCA1-mutant human breast cancer cell lines. Anticancer Res. 2010, 30, 2049–2054. [Google Scholar]
- Erin, N.; Bronson, S.K.; Billingsley, M.L. Calcium-dependent interaction of calcineurin with Bcl-2 in neuronal tissue. Neuroscience 2003, 117, 541–555. [Google Scholar] [CrossRef]
- Shibasaki, F.; McKeon, F. Calcineurin functions in Ca(2+)-activated cell death in mammalian cells. J. Cell Biol. 1995, 131, 735–743. [Google Scholar] [CrossRef]
- Anderson, N.S.; Turner, L.; Livingston, S.; Chen, R.; Nicosia, S.V.; Kruk, P.A. Bcl-2 expression is altered with ovarian tumor progression: An immunohistochemical evaluation. J. Ovarian Res. 2009, 2, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herod, J.J.; Eliopoulos, A.G.; Warwick, J.; Niedobitek, G.; Young, L.S.; Kerr, D.J. The prognostic significance of Bcl-2 and p53 expression in ovarian carcinoma. Cancer Res. 1996, 56, 2178–2184. [Google Scholar]
- Yuan, J.; Lan, H.; Jiang, X.; Zeng, D.; Xiao, S. Bcl-2 family: Novel insight into individualized therapy for ovarian cancer (Review). Int. J. Mol. Med. 2020, 46, 1255–1265. [Google Scholar] [CrossRef]
- O’Neill, C.J.; Deavers, M.T.; Malpica, A.; Foster, H.; McCluggage, W.G. An immunohistochemical comparison between low-grade and high-grade ovarian serous carcinomas: Significantly higher expression of p53, MIB1, BCL2, HER-2/neu, and C-KIT in high-grade neoplasms. Am. J. Surg. Pathol. 2005, 29, 1034–1041. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Crasta, J.A. An Immunohistochemical Comparison of P53 and Bcl-2 as Apoptotic and MIB1 as Proliferative Markers in Low-Grade and High-Grade Ovarian Serous Carcinomas. Int. J. Gynecol. Cancer 2010, 20, 537–541. [Google Scholar] [CrossRef]
- Kalogeraki, A.; Tamiolakis, D.; Matalliotaki, C.; Karvela-Kalogeraki, I.; Karvelas-Kalogerakis, M.; Segredakis, J.; Sinatkas, V.; Matalliotakis, I. The prognostic significance of p53, bcl2 and mib1 expressions related with other clinicopathological variables in serous ovarian carcinomas. A clinicopathological study in peritoneal fluids. Rev. Med. Chir Soc. Med. Nat. Iasi 2015, 119, 454–460. [Google Scholar]
- Weiwad, M.; Edlich, F.; Erdmann, F.; Jarczowski, F.; Kilka, S.; Dorn, M.; Pechstein, A.; Fischer, G. A reassessment of the inhibitory capacity of human FKBP38 on calcineurin. FEBS Lett. 2005, 579, 1591–1596. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Nylander, K.D.; Yan, C.; Schor, N.F. Role of caspase 3-dependent Bcl-2 cleavage in potentiation of apoptosis by Bcl-2. Mol. Pharmacol. 2002, 61, 142–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, J.; Yang, H.; Cui, T.; Pan, P.; Kabir, N.; Chen, D.; Ma, J.; Chen, X.; Chen, Y.; Yang, Y. Combined treatment with sorafenib and silibinin synergistically targets both HCC cells and cancer stem cells by enhanced inhibition of the phosphorylation of STAT3/ERK/AKT. Eur. J. Pharmacol. 2018, 832, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Li, Z.; Dai, C.; Zhao, D.; Wang, Y.; Ma, C.; Liu, C. Chrysophanol inhibits proliferation and induces apoptosis through NF-κB/cyclin D1 and NF-κB/Bcl-2 signaling cascade in breast cancer cell lines. Mol. Med. Rep. 2018, 17, 4376–4382. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Zhao, B.; Chang, H.; Xiao, M.; Wu, Y.; Liu, Y. Paclitaxel suppresses proliferation and induces apoptosis through regulation of ROS and the AKT/MAPK signaling pathway in canine mammary gland tumor cells. Mol. Med. Rep. 2018, 17, 8289–8299. [Google Scholar] [CrossRef] [Green Version]
- Jahani-Asl, A.; Basak, A.; Tsang, B.K. Caspase-3-mediated cleavage of Akt: Involvement of non-consensus sites and influence of phosphorylation. FEBS Lett. 2007, 581, 2883–2888. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Liu, D.; Songyang, Z. The role of Asp-462 in regulating Akt activity. J. Biol. Chem. 2002, 277, 35561–35566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Héron-Milhavet, L.; Franckhauser, C.; Rana, V.; Berthenet, C.; Fisher, D.; Hemmings, B.A.; Fernandez, A.; Lamb, N.J. Only Akt1 is required for proliferation, while Akt2 promotes cell cycle exit through p21 binding. Mol. Cell. Biol. 2006, 26, 8267–8280. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.C.; Lin, K.T.; Chen, C.H.; Lee, S.A.; Lee, P.Y.; Liu, Y.W.; Kuo, Y.L.; Wang, F.S.; Lai, J.M.; Huang, C.Y. Using an in situ proximity ligation assay to systematically profile endogenous protein-protein interactions in a pathway network. J. Proteome Res. 2014, 13, 5339–5346. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anastasiadou, E.; Messina, E.; Sanavia, T.; Labruna, V.; Ceccarelli, S.; Megiorni, F.; Gerini, G.; Pontecorvi, P.; Camero, S.; Perniola, G.; et al. Calcineurin Gamma Catalytic Subunit PPP3CC Inhibition by miR-200c-3p Affects Apoptosis in Epithelial Ovarian Cancer. Genes 2021, 12, 1400. https://doi.org/10.3390/genes12091400
Anastasiadou E, Messina E, Sanavia T, Labruna V, Ceccarelli S, Megiorni F, Gerini G, Pontecorvi P, Camero S, Perniola G, et al. Calcineurin Gamma Catalytic Subunit PPP3CC Inhibition by miR-200c-3p Affects Apoptosis in Epithelial Ovarian Cancer. Genes. 2021; 12(9):1400. https://doi.org/10.3390/genes12091400
Chicago/Turabian StyleAnastasiadou, Eleni, Elena Messina, Tiziana Sanavia, Vittorio Labruna, Simona Ceccarelli, Francesca Megiorni, Giulia Gerini, Paola Pontecorvi, Simona Camero, Giorgia Perniola, and et al. 2021. "Calcineurin Gamma Catalytic Subunit PPP3CC Inhibition by miR-200c-3p Affects Apoptosis in Epithelial Ovarian Cancer" Genes 12, no. 9: 1400. https://doi.org/10.3390/genes12091400
APA StyleAnastasiadou, E., Messina, E., Sanavia, T., Labruna, V., Ceccarelli, S., Megiorni, F., Gerini, G., Pontecorvi, P., Camero, S., Perniola, G., Venneri, M. A., Trivedi, P., Lenzi, A., & Marchese, C. (2021). Calcineurin Gamma Catalytic Subunit PPP3CC Inhibition by miR-200c-3p Affects Apoptosis in Epithelial Ovarian Cancer. Genes, 12(9), 1400. https://doi.org/10.3390/genes12091400