
Cyanogenesis in the Sorghum Genus: From Genotype to Phenotype

 , , , , , ,
, , , , , ,  , and
, and 
Abstract
:1. Introduction
2. Results
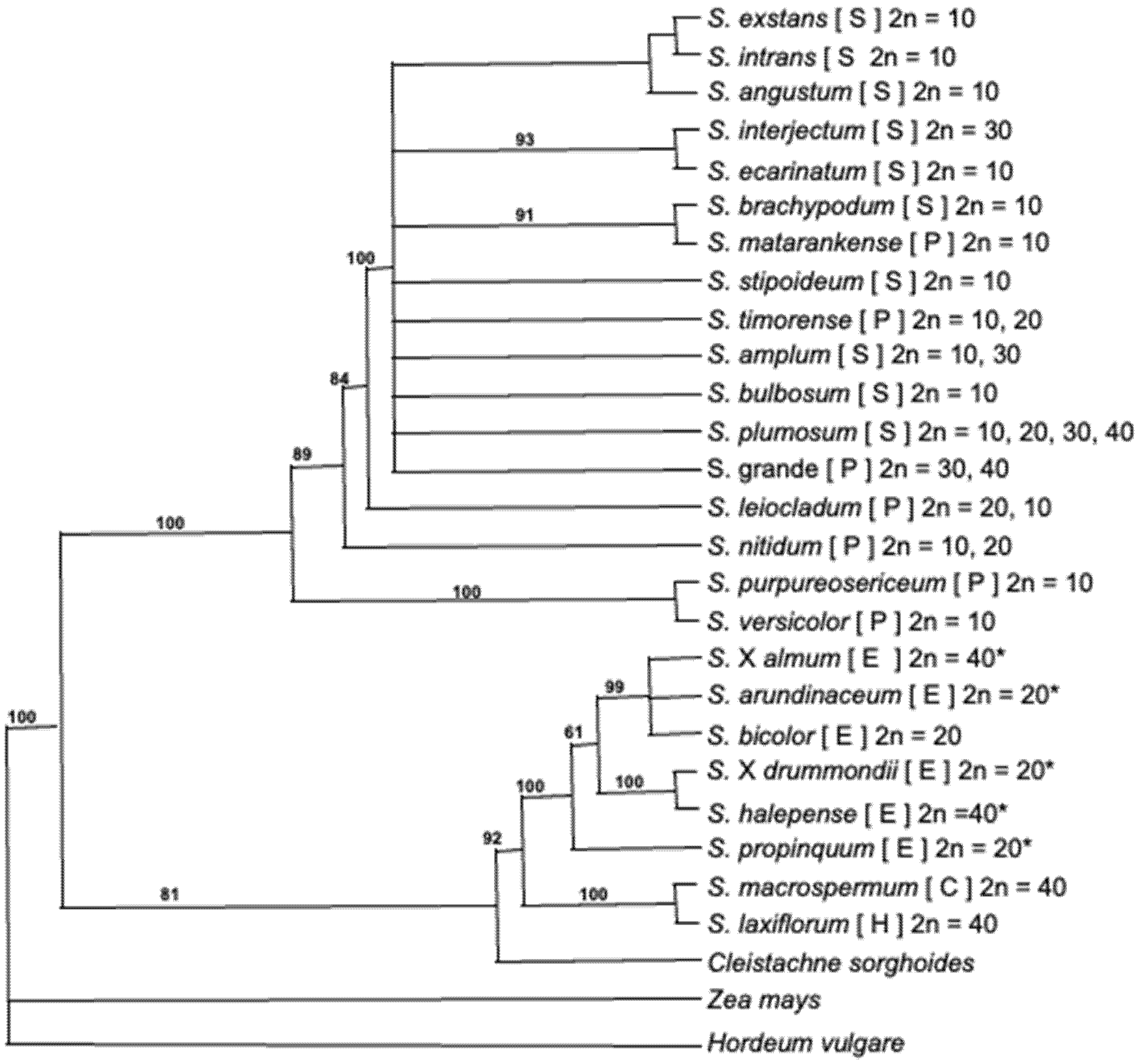
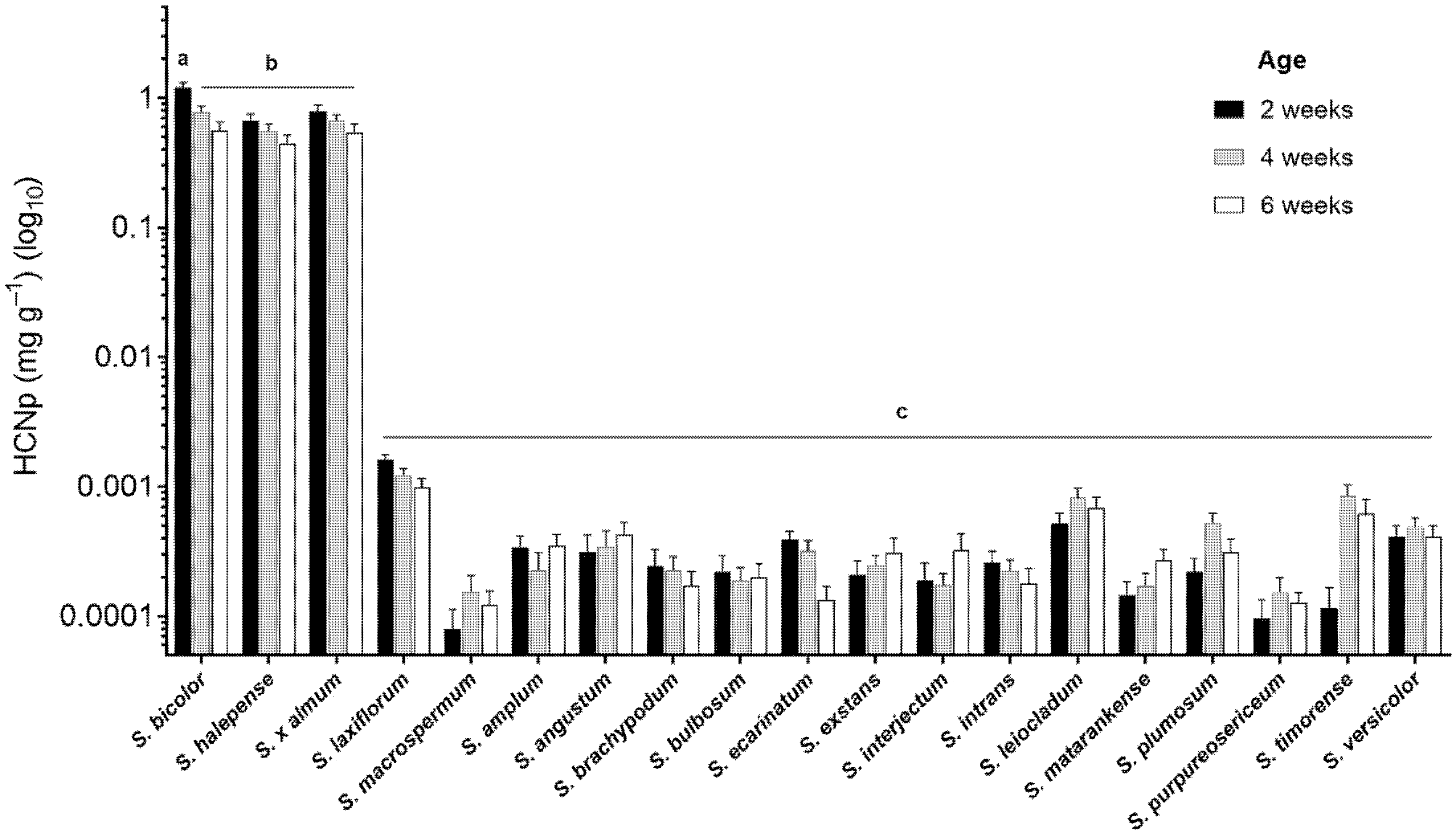
2.1. Hydrogen Cyanide Potential in the Wild Relatives of Sorghum
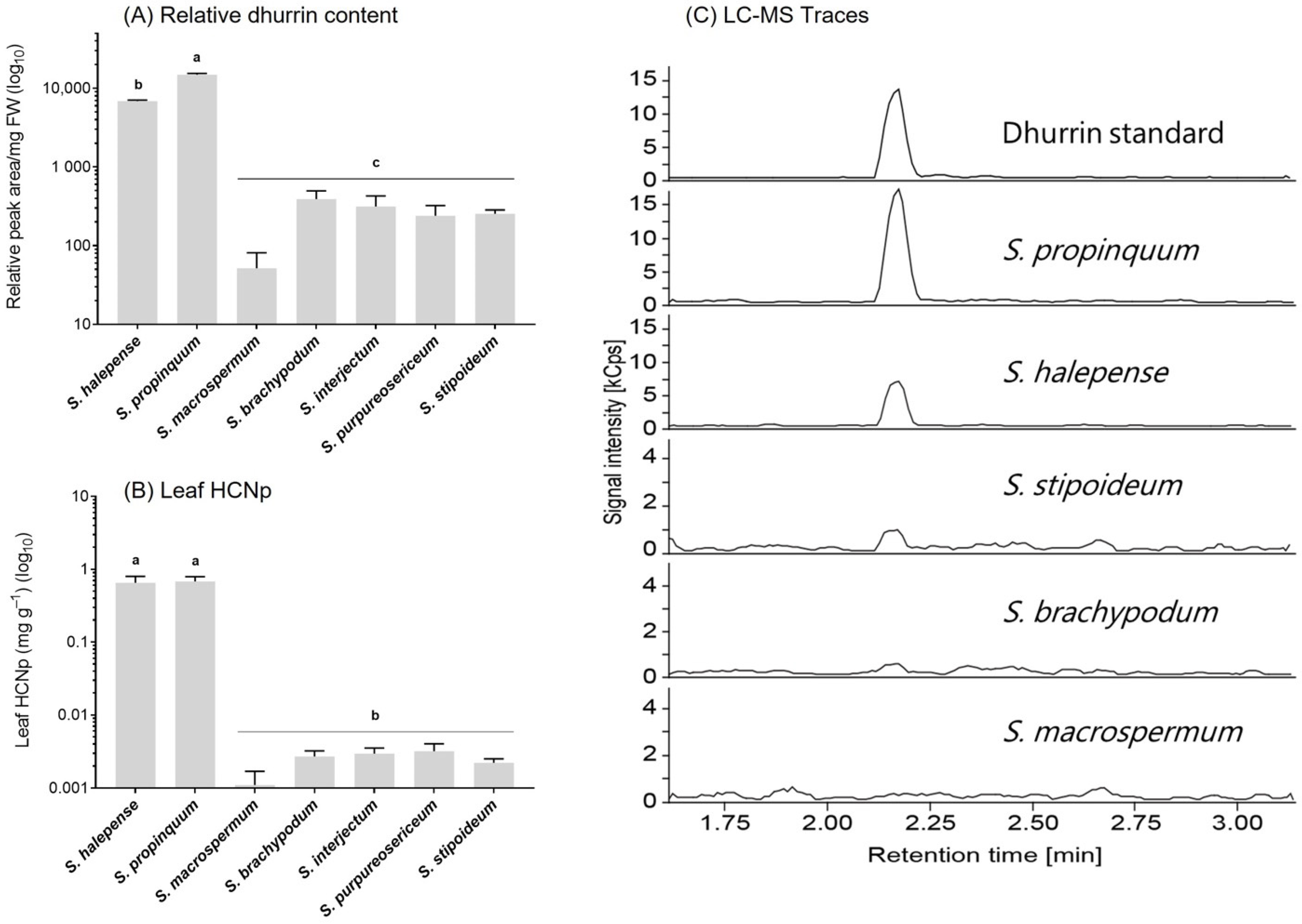
2.2. Relative Cyanogenic Glucoside Content
2.3. Variation within Cyanogenesis Related Genes in Sorghum Detected by Genome Sequencing
2.4. Variation within the Key Biosynthesis Gene, CYP79A1
3. Discussion
3.1. Phenotypic Variation of Cyanogenesis in Sorghum
3.2. Genomic Variation of Cyanogenesis in Sorghum
4. Materials and Methods
4.1. Plant Material and Growing Conditions
4.2. Hydrogen Cyanide Assays
4.3. LC-MS Analysis and Identification of Cyanogenic Glucoside(s)
4.4. Variant Analysis within Selected Genes Involved in Cyanogenesis and Related Pathways
4.5. Detailed Sequence Analysis of CYP79A1 across the Sorghum Phylogeny
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sage, R.F.; Zhu, X.-G. Exploiting the engine of C4 photosynthesis. J. Exp. Bot. 2011, 62, 2989–3000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tari, I.; Laskay, G.; Takács, Z.; Poór, P. Response of sorghum to abiotic stresses: A review. J. Agron. Crop Sci. 2013, 199, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Gleadow, R.M.; Møller, B.L. Cyanogenic glycosides: Synthesis, physiology, and phenotypic plasticity. Annu. Rev. Plant Biol. 2014, 65, 155–185. [Google Scholar] [CrossRef]
- Morant, A.V.; Jørgensen, K.; Jørgensen, C.; Paquette, S.M.; Sánchez-Pérez, R.; Møller, B.L.; Bak, S. β-Glucosidases as detonators of plant chemical defense. Phytochemistry 2008, 69, 1795–1813. [Google Scholar] [CrossRef]
- Heraud, P.; Cowan, M.F.; Marzec, K.M.; Møller, B.L.; Blomstedt, C.K.; Gleadow, R. Label-free Raman hyperspectral imaging analysis localizes the cyanogenic glucoside dhurrin to the cytoplasm in sorghum cells. Sci. Rep. 2018, 8, 2691. [Google Scholar] [CrossRef] [PubMed]
- Zagrobelny, M.; Bak, S.; Møller, B.L. Cyanogenesis in plants and arthropods. Phytochemistry 2008, 69, 1457–1468. [Google Scholar] [CrossRef]
- Gleadow, R.M.; Woodrow, I.E. Constraints on effectiveness of cyanogenic glycosides in herbivore defense. J. Chem. Ecol. 2002, 28, 1301–1313. [Google Scholar] [CrossRef] [PubMed]
- Tattersall, D.B.; Bak, S.; Jones, P.R.; Olsen, C.E.; Nielsen, J.K.; Hansen, M.L.; Høj, P.B.; Møller, B.L. Resistance to an herbivore through engineered cyanogenic glucoside synthesis. Science 2001, 293, 1826–1828. [Google Scholar] [CrossRef] [PubMed]
- Ballhorn, D.J.; Kautz, S.; Heil, M.; Hegeman, A.D. Cyanogenesis of wild lima bean (Phaseolus lunatus L.) Is an efficient direct defence in nature. PLoS ONE 2009, 4, e5450. [Google Scholar] [CrossRef]
- Wheeler, J.L.; Mulcahy, C.; Walcott, J.J.; Rapp, G.G. Factors affecting the hydrogen cyanide potential of forage sorghum. Aust. J. Agric. Res. 1990, 41, 1093–1100. [Google Scholar] [CrossRef]
- Andersen, M.D.; Busk, P.K.; Svendsen, I.; Møller, B.L. Cytochromes P-450 from cassava (Manihot esculenta Crantz) catalyzing the first steps in the biosynthesis of the cyanogenic glucosides linamarin and lotaustralin: Cloning, functional expression in pichia pastoris, and substrate specificity of the isolated recombinant enzymes. J. Biol. Chem. 2000, 275, 1966–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlert, M.; Jagd, L.M.; Braumann, I.; Dockter, C.; Crocoll, C.; Motawia, M.S.; Møller, B.L.; Lyngkjær, M.F. Deletion of biosynthetic genes, specific SNP patterns and differences in transcript accumulation cause variation in hydroxynitrile glucoside content in barley cultivars. Sci. Rep. 2019, 9, 5730. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Pérez, R.; Pavan, S.; Mazzeo, R.; Moldovan, C.; Cigliano, R.A.; Del Cueto, J.; Ricciardi, F.; Lotti, C.; Ricciardi, L.; Dicenta, F.; et al. Mutation of a bHLH transcription factor allowed almond domestication. Science 2019, 364, 1095–1098. [Google Scholar] [CrossRef] [PubMed]
- Hansen, C.C.; Sørensen, M.; Veiga, T.A.M.; Zibrandtsen, J.F.S.; Heskes, A.M.; Olsen, C.E.; Boughton, B.A.; Møller, B.L.; Neilson, E.H.J. Reconfigured cyanogenic glucoside biosynthesis in Eucalyptus cladocalyx involves a cytochrome P450, CYP706C55. Plant Physiol. 2018, 178, 1081–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thodberg, S.; Sørensen, M.; Bellucci, M.; Crocoll, C.; Bendtsen, A.K.; Nelson, D.R.; Motawia, M.S.; Møller, B.L.; Neilson, E.H.J. A flavin-dependent monooxygenase catalyzes the initial step in cyanogenic glycoside synthesis in ferns. Commun. Biol. 2020, 3, 507. [Google Scholar] [CrossRef]
- Gleadow, R.; Pegg, A.; Blomstedt, C.K. Resilience of cassava (Manihot esculenta Crantz) to salinity: Implications for food security in low-lying regions. J. Exp. Bot. 2016, 67, 5403–5413. [Google Scholar] [CrossRef] [Green Version]
- Bredeson, J.V.; Lyons, J.B.; Prochnik, S.E.; Wu, G.A.; Ha, C.M.; Edsinger, E.; Grimwood, J.; Schmutz, J.; Rabbi, I.Y.; Egesi, C.; et al. Sequencing wild and cultivated cassava and related species reveals extensive interspecific hybridization and genetic diversity. Nat. Biotechnol. 2016, 34, 562–570. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.A. Why are so many food plants cyanogenic? Phytochemistry 1998, 47, 155–162. [Google Scholar] [CrossRef]
- McKey, D.; Cavagnaro, T.R.; Cliff, J.; Gleadow, R. Chemical ecology in coupled human and natural systems: People, manioc, multitrophic interactions and global change. Chemoecology 2010, 20, 109–133. [Google Scholar] [CrossRef]
- Franzmann, B.A.; Hardy, A.T.; Murray, D.A.H.; Henzell, R.G. Host-plant resistance and biopesticides: Ingredients for successful integrated pest management (IPM) in Australian sorghum production. Aust. J. Exp. Agric. 2008, 48, 1594–1600. [Google Scholar] [CrossRef]
- Sharma, H.C.; Franzmann, B.A.; Henzell, R.G. Mechanisms and diversity of resistance to sorghum midge, Stenodiplosis sorghicola in Sorghum bicolor. Euphytica 2002, 124, 1–12. [Google Scholar] [CrossRef]
- Fuller, D.Q.; Stevens, C.J. Sorghum Domestication and Diversification: A Current Archaeobotanical Perspective. In Plants and People in the African Past: Progress in African Archaeobotany; Mercuri, A.M., D’Andrea, A.C., Fornaciari, R., Höhn, A., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 427–452. [Google Scholar] [CrossRef]
- Dillon, S.L.; Shapter, F.M.; Henry, R.J.; Cordeiro, G.; Izquierdo, L.; Lee, L.S. Domestication to crop improvement: Genetic resources for Sorghum and Saccharum (Andropogoneae). Ann. Bot. 2007, 100, 975–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winchell, F.; Brass, M.; Manzo, A.; Beldados, A.; Perna, V.; Murphy, C.; Stevens, C.; Fuller, D.Q. On the origins and dissemination of domesticated sorghum and pearl millet across Africa and into India: A view from the Butana group of the far eastern Sahel. Afr. Archaeol. Rev. 2018, 35, 483–505. [Google Scholar] [CrossRef]
- Nielsen, L.J.; Stuart, P.; Pičmanová, M.; Rasmussen, S.; Olsen, C.E.; Harholt, J.; Møller, B.L.; Bjarnholt, N. Dhurrin metabolism in the developing grain of Sorghum bicolor (L.) Moench investigated by metabolite profiling and novel clustering analyses of time-resolved transcriptomic data. BMC Genom. 2016, 17, 1021. [Google Scholar] [CrossRef] [Green Version]
- Nassar, N.M.A.; Fichtner, S.S. Hydrocyanic acid content in some wild Manihot (cassava) species. Can. J. Plant Sci. 1978, 58, 577–578. [Google Scholar] [CrossRef]
- Nassar, N.M.A. Wild cassava, Manihot spp.: Biology and potentialities for genetic improvement. Genet. Mol. Biol. 2000, 23, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Blaim, H.; Nowacki, E. Cyanogensis in Lotus and Trifolium species. Acta Agrobot. 1979, 32, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Blaise, S.; Carter, D.; Reynaud, J. Evolution and differentiation of Lotus corniculatus/Lotus alpinus populations from French South-Western Alps. I. Morphologic and cyanogenic variations. Evol. Trends Plants 1991, 5, 137–148. [Google Scholar]
- Band, L.; Heyn, C.C.; Plitmann, U. Distribution of cyanogenesis in Lotus (Leguminosae). Taxon 1981, 30, 601–608. [Google Scholar] [CrossRef]
- Dahler, J.M.; McConchie, C.; Turnbull, C.G.N. Quantification of cyanogenic glycosides in seedlings of three Macadamia (Proteaceae) species. Aust. J. Bot. 1995, 43, 619–628. [Google Scholar] [CrossRef]
- Selmar, D.; Lieberei, R.; Junqueira, N.; Biehl, B. Changes in cyanogenic glucoside content in seeds and seedlings of Hevea species. Phytochemistry 1991, 30, 2135–2140. [Google Scholar] [CrossRef]
- Dillon, S.L.; Lawrence, P.K.; Henry, R.J.; Price, H.J. Sorghum resolved as a distinct genus based on combined ITS1, ndhF and Adh1 analyses. Plant Syst. Evol. 2007, 268, 29–43. [Google Scholar] [CrossRef]
- Ananda, G.K.S.; Myrans, H.; Norton, S.L.; Gleadow, R.; Furtado, A.; Henry, R.J. Wild sorghum as a promising resource for crop improvement. Front. Plant Sci. 2020, 11, 1108. [Google Scholar] [CrossRef] [PubMed]
- Ananda, G.; Norton, S.; Blomstedt, C.; Furtado, A.; Møller, B.L.; Gleadow, R.; Henry, R. Phylogenetic relationships in the Sorghum genus based on sequencing of the chloroplast and nuclear genes. Plant Genome 2021, 14, e20123. [Google Scholar] [CrossRef]
- Lazarides, M.; Hacker, J.B.; Andrew, M.H. Taxonomy, cytology and ecology of indigenous Australian sorghums (Sorghum Moench: Andropogoneae: Poaceae). Aust. Syst. Bot. 1991, 4, 591–635. [Google Scholar] [CrossRef]
- Nicollier, G.F.; Pope, D.F.; Thompson, A.C. Biological activity of dhurrin and other compounds from Johnson grass (Sorghum halepense). J. Agric. Food Chem. 1983, 31, 744–748. [Google Scholar] [CrossRef]
- Gray, E.; Rice, J.S.; Wattenbarger, D.; Benson, J.A.; Hester, A.J.; Loyd, R.C.; Greene, B.M. Hydrocyanic Acid Potential of Sorghum Plants Grown in Tennessee; Agricultural Experiment Station, The University of Tennessee: Knoxville, TN, USA, 1968; pp. 1–48. [Google Scholar]
- Myrans, H.; Vandegeer, R.K.; Henry, R.J.; Gleadow, R.M. Nitrogen availability and allocation in sorghum and its wild relatives: Divergent roles for cyanogenic glucosides. J. Plant Physiol. 2021, 258–259, 153393. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.F.; Blomstedt, C.K.; Møller, B.L.; Henry, R.J.; Gleadow, R.M. Variation in production of cyanogenic glucosides during early plant development: A comparison of wild and domesticated sorghum. Phytochemistry 2021, 184, 112645. [Google Scholar] [CrossRef]
- Cowan, M.F.; Blomstedt, C.K.; Norton, S.L.; Henry, R.J.; Møller, B.L.; Gleadow, R. Crop wild relatives as a genetic resource for generating low-cyanide, drought-tolerant Sorghum. Environ. Exp. Bot. 2020, 169, 103884. [Google Scholar] [CrossRef]
- Price, H.J.; Dillon, S.L.; Hodnett, G.; Rooney, W.L.; Ross, L.; Johnston, J.S. Genome evolution in the genus Sorghum (Poaceae). Ann. Bot. 2005, 95, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Liu, H.; Wen, J. Relationships between Sorghum bicolor (Poaceae) and its close relatives based on genomic in situ hybridization evidence. Turk. J. Bot. 2017, 41, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Winchell, F.; Stevens, C.J.; Murphy, C.; Champion, L.; Fuller, D. Evidence for sorghum domestication in fourth millennium BC Eastern Sudan: Spikelet morphology from ceramic impressions of the butana group. Curr. Anthropol. 2017, 58, 673–683. [Google Scholar] [CrossRef] [Green Version]
- De Wet, J.M.J.; Harlan, J.R. The origin and domestication of Sorghum bicolor. Econ. Bot. 1971, 25, 128–135. [Google Scholar] [CrossRef]
- Venkateswaran, K.; Elangovan, M.; Sivaraj, N. Origin, domestication and diffusion of Sorghum bicolor. In Breeding Sorghum for Diverse End Uses; Aruna, C., Visarada, K.B.R.S., Bhat, B.V., Tonapi, V.A., Eds.; Woodhead Publishing, Sawston, UK: Sawston, UK, 2019; pp. 15–31. [Google Scholar]
- Anon. Sorghum × almum Parodi: Hybrid between Sorghum halepense × S. bicolor, Poaceae. Available online: https://www.hort.purdue.edu/newcrop/duke_energy/Sorghum_Xalmum.html (accessed on 10 January 2022).
- Norton, S.L.; Khoury, C.K.; Sosa, C.C.; Castañeda-Álvarez, N.P.; Achicanoy, H.A.; Sotelo, S. Priorities for enhancing the ex situ conservation and use of Australian crop wild relatives. Aust. J. Bot. 2017, 65, 638–645. [Google Scholar] [CrossRef]
- Myrans, H.; Diaz, M.V.; Khoury, C.K.; Carver, D.; Henry, R.J.; Gleadow, R. Modelled distributions and conservation priorities of wild sorghums (Sorghum Moench). Divers. Distrib. 2020, 26, 1727–1740. [Google Scholar] [CrossRef]
- Herms, D.A.; Mattson, W.J. The dilemma of plants—To grow or defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef] [Green Version]
- McKey, D. Adaptive patterns in alkaloid physiology. Am. Nat. 1974, 108, 305–320. [Google Scholar] [CrossRef]
- Busk, P.K.; Møller, B.L. Dhurrin synthesis in sorghum is regulated at the transcriptional level and induced by nitrogen fertilization in older plants. Plant Physiol. 2002, 129, 1222–1231. [Google Scholar] [CrossRef] [Green Version]
- Darbani, B.; Motawia, M.S.; Olsen, C.E.; Nour-Eldin, H.H.; Møller, B.L.; Rook, F. The biosynthetic gene cluster for the cyanogenic glucoside dhurrin in Sorghum bicolor contains its co-expressed vacuolar MATE transporter. Sci. Rep. 2016, 6, 37079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, C.M.; Burow, G.B.; Brown, P.J.; Thurber, C.; Xin, Z.; Burke, J.J. Natural variation in synthesis and catabolism genes influences dhurrin content in Sorghum. Plant Genome 2015, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjarnholt, N.; Neilson, E.H.J.; Crocoll, C.; Jorgensen, K.; Motawia, M.S.; Olsen, C.E.; Dixon, D.P.; Edwards, R.; Møller, B.L. Glutathione transferases catalyze recycling of auto-toxic cyanogenic glucosides in sorghum. Plant J. Cell Mol. Biol. 2018, 94, 1109–1125. [Google Scholar] [CrossRef] [Green Version]
- Yachdav, G.; Kloppmann, E.; Kajan, L.; Hecht, M.; Goldberg, T.; Hamp, T.; Hönigschmid, P.; Schafferhans, A.; Roos, M.; Bernhofer, M.; et al. PredictProtein—An open resource for online prediction of protein structural and functional features. Nucleic Acids Res. 2014, 42, W337–W343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, K.; Osmani, S.A.; Hamann, T.; Naur, P.; Møller, B.L. Homology modeling of the three membrane proteins of the dhurrin metabolon: Catalytic sites, membrane surface association and protein–protein interactions. Phytochemistry 2011, 72, 2113–2123. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Albacete, D.; Montefiori, M.; Kol, S.; Motawia, M.S.; Møller, B.L.; Olsen, L.; Nørholm, M.H.H. The CYP79A1 catalyzed conversion of tyrosine to (E)-p-hydroxyphenylacetaldoxime unravelled using an improved method for homology modeling. Phytochemistry 2017, 135, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Hasemann, C.A.; Kurumbail, R.G.; Boddupalli, S.S.; Peterson, J.A.; Deisenhofer, J. Structure and function of cytochromes P450:a comparative analysis of three crystal structures. Structure 1995, 3, 41–62. [Google Scholar] [CrossRef]
- Blomstedt, C.K.; Gleadow, R.M.; O’Donnell, N.; Naur, P.; Jensen, K.; Laursen, T.; Olsen, C.E.; Stuart, P.; Hamill, J.D.; Møller, B.L.; et al. A combined biochemical screen and TILLING approach identifies mutations in Sorghum bicolor L. Moench resulting in acyanogenic forage production. Plant Biotechnol. J. 2012, 10, 54–66. [Google Scholar] [CrossRef]
- Skelton, J.L. EMS Induced Mutations in Dhurrin Metabolism and Their Impacts on Sorghum Growth and Development; Purdue University: West Lafayette, IN, USA, 2014. [Google Scholar]
- Knoch, E.; Motawie, M.S.; Olsen, C.E.; Møller, B.L.; Lyngkjær, M.F. Biosynthesis of the leucine derived α-, β- and γ-hydroxynitrile glucosides in barley (Hordeum vulgare L.). Plant J. Cell Mol. Biol. 2016, 88, 247–256. [Google Scholar] [CrossRef]
- Kahn, R.; Fahrendorf, T.; Halkier, B.; Møller, B. Substrate specificity of the cytochrome P450 enzymes CYP79A1 and CYP71E1 involved in the biosynthesis of the cyanogenic glucoside dhurrin in Sorghum bicolor (L.) Moench. Arch. Biochem. Biophys. 1999, 363, 9–18. [Google Scholar] [CrossRef]
- Gleadow, R.M.; Veechies, A.C.; Woodrow, I.E. Cyanogenic Eucalyptus nobilis is polymorphic for both prunasin and specific β-glucosidases. Phytochemistry 2003, 63, 699–704. [Google Scholar] [CrossRef]
- O’Donnell, N.H.; Møller, B.L.; Neale, A.D.; Hamill, J.D.; Blomstedt, C.K.; Gleadow, R.M. Effects of PEG-induced osmotic stress on growth and dhurrin levels of forage sorghum. Plant Physiol. Biochem. 2013, 73, 83–92. [Google Scholar] [CrossRef]
- Gleadow, R.M.; Ottman, M.J.; Kimball, B.A.; Wall, G.W.; Pinter, P.J.; LaMorte, R.L.; Leavitt, S.W. Drought-induced changes in nitrogen partitioning between cyanide and nitrate in leaves and stems of sorghum grown at elevated CO2 are age dependent. Field Crops Res. 2016, 185, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Blomstedt, C.K.; Rosati, V.C.; Møller, B.L.; Gleadow, R. Counting the costs: Nitrogen partitioning in Sorghum mutants. Funct. Plant Biol. 2018, 45, 705–718. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.E.; Gleadow, R.M.; Cavagnaro, T.R. Age versus stage: Does ontogeny modify the effect of phosphorus and arbuscular mycorrhizas on above- and below-ground defence in forage sorghum? Plant Cell Environ. 2014, 37, 929–942. [Google Scholar] [CrossRef]
- Gleadow, R.M.; McKinley, B.A.; Blomstedt, C.K.; Lamb, A.C.; Møller, B.L.; Mullet, J.E. Regulation of dhurrin pathway gene expression during Sorghum bicolor development. Planta 2021, 254, 119. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.J.; Chapin, F.S.; Mooney, H.A. Resource Limitation in Plants-An Economic Analogy. Annu. Rev. Ecol. Syst. 1985, 16, 363–392. [Google Scholar] [CrossRef]
- Wiedemuth, K.; Muller, J.; Kahlau, A.; Amme, S.; Mock, H.P.; Grzam, A.; Hell, R.; Egle, K.; Beschow, H.; Humbeck, K. Successive maturation and senescence of individual leaves during barley whole plant ontogeny reveals temporal and spatial regulation of photosynthetic function in conjunction with C and N metabolism. J. Plant Physiol. 2005, 162, 1226–1236. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.A. The role of trichomes in plant defense. Q. Rev. Biol. 1973, 48, 3–15. [Google Scholar] [CrossRef]
- Tian, D.; Tooker, J.; Peiffer, M.; Chung, S.H.; Felton, G.W. Role of trichomes in defense against herbivores: Comparison of herbivore response to woolly and hairless trichome mutants in tomato (Solanum lycopersicum). Planta 2012, 236, 1053–1066. [Google Scholar] [CrossRef] [PubMed]
- Johnson, H.B. Plant pubescence: An ecological perspective. Bot. Rev. 1975, 41, 233–258. [Google Scholar] [CrossRef]
- Pičmanová, M.; Neilson, E.H.; Motawia, M.S.; Olsen, C.E.; Agerbirk, N.; Gray, C.J.; Flitsch, S.; Meier, S.; Silvestro, D.; Jorgensen, K.; et al. A recycling pathway for cyanogenic glycosides evidenced by the comparative metabolic profiling in three cyanogenic plant species. Biochem. J. 2015, 469, 375–389. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, F.B.; Cho, S.K.; Olsen, C.E.; Yang, S.W.; Møller, B.L.; Jørgensen, K. Diurnal regulation of cyanogenic glucoside biosynthesis and endogenous turnover in cassava. Plant Direct 2018, 2, e00038. [Google Scholar] [CrossRef]
- Goodger, J.Q.D.; Capon, R.J.; Woodrow, I.E. Cyanogenic polymorphism in Eucalyptus polyanthemos Schauer subsp. vestita L. Johnson and K. Hill (Myrtaceae). Biochem. Syst. Ecol. 2002, 30, 617–630. [Google Scholar] [CrossRef]
- Gleadow, R.M.; Haburjak, J.; Dunn, J.E.; Conn, M.E.; Conn, E.E. Frequency and distribution of cyanogenic glycosides in Eucalyptus L’Herit. Phytochemistry 2008, 69, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Gleadow, R.M.; Woodrow, I.E. Polymorphism in cyanogenic glycoside content and cyanogenic β-glucosidase activity in natural populations of Eucalyptus cladocalyx. Aust. J. Plant Physiol. 2000, 27, 693–699. [Google Scholar] [CrossRef]
- Armstrong, H.E.; Armstrong Edward, F.; Horton, E. Herbage Studies. II. Variation in Lotus corniculatus and Trifolium repens (cyanophoric plants). Proc. R. Soc. Lond. Ser. B 1913, 86, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Aikman, K.; Bergman, D.; Ebinger, J.; Seigler, D. Variation of cyanogenesis in some plant species of the midwestern United States. Biochem. Syst. Ecol. 1996, 24, 637–645. [Google Scholar] [CrossRef]
- Jones, D.A. On the polymorphism of cyanogenesis in Lotus corniculatus L. Heredity 1977, 39, 27–44. [Google Scholar] [CrossRef] [Green Version]
- Hughes, M.A. The cyanogenic polymorphism in Trifolium repens L. (white clover). Heredity 1991, 66, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Olsen, K.M.; Sutherland, B.L.; Small, L.L. Molecular evolution of the Li/li chemical defence polymorphism in white clover (Trifolium repens L.). Mol. Ecol. 2007, 16, 4180–4193. [Google Scholar] [CrossRef]
- Kakes, P.; Hakvoort, H.W.J. On the origin of the cyanogenic polymorphism in Trifolium repens L. J. Evol. Biol. 1994, 7, 201–215. [Google Scholar] [CrossRef]
- Takos, A.M.; Knudsen, C.; Lai, D.; Kannangara, R.; Mikkelsen, L.; Motawia, M.S.; Olsen, C.E.; Sato, S.; Tabata, S.; Jørgensen, K.; et al. Genomic clustering of cyanogenic glucoside biosynthetic genes aids their identification in Lotus japonicus and suggests the repeated evolution of this chemical defence pathway. Plant J. Cell Mol. Biol. 2011, 68, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Thodberg, S.; Del Cueto, J.; Mazzeo, R.; Pavan, S.; Lotti, C.; Dicenta, F.; Neilson, E.H.J.; Møller, B.L.; Sánchez-Pérez, R. Elucidation of the amygdalin pathway reveals the metabolic basis of bitter and sweet almonds (Prunus dulcis). Plant Physiol. 2018, 178, 1096–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brozynska, M.; Furtado, A.; Henry, R.J. Genomics of crop wild relatives: Expanding the gene pool for crop improvement. Plant Biotechnol. J. 2016, 14, 1070–1085. [Google Scholar] [CrossRef] [PubMed]
- Gleadow, R.; Bjarnholt, N.; Jørgensen, K.; Fox, J.; Miller, R.M. Detection, identification and quantitative measurement of cyanogenic glycosides. In Research Methods in Plant Science. Vol. 1. Soil Allelochemicals; International Allelopathy Foundation, Studium Press LLC: Houston, TX, USA, 2012; Volume 1, pp. 283–310. [Google Scholar]
- Montini, L.; Crocoll, C.; Gleadow, R.M.; Motawia, M.S.; Janfelt, C.; Bjarnholt, N. Matrix-Assisted Laser Desorption/Ionization-Mass Spectrometry Imaging of Metabolites during Sorghum Germination. Plant Physiol. 2020, 183, 925–942. [Google Scholar] [CrossRef] [PubMed]
- Møller, B.L.; Olsen, C.E.; Motawia, M.S. General and stereocontrolled approach to the chemical synthesis of naturally occurring cyanogenic glucosides. J. Nat. Prod. 2016, 79, 1198–1202. [Google Scholar] [CrossRef] [Green Version]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The ensembl variant effect predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [Green Version]
- Allen, G.C.; Flores-Vergara, M.A.; Krasynanski, S.; Kumar, S.; Thompson, W.F. A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nat. Protoc. 2006, 1, 2320–2325. [Google Scholar] [CrossRef]
- Halkier, B.A.; Møller, B.L. Biosynthesis of the cyanogenic glucoside dhurrin in seedlings of Sorghum bicolor (L) Moench and partial purification of the enzyme system involved. Plant Physiol. 1989, 90, 1552–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sorghum Species | ||||||
|---|---|---|---|---|---|---|
| Gene | Lax | Mac | Bra | Lei | Mat | Pur |
| CYP79A1 | 285 | 146 | 258 | 406 | 338 | 235 |
| CYP71E1 | 130 | 306 | 270 | 232 | 232 | 157 |
| UGT85B1 | 157 | 104 | 99 | 155 | 177 | 154 |
| POR | 295 | 331 | 322 | 297 | 233 | 234 |
| NIT4A | 167 | 237 | 248 | 198 | 181 | 149 |
| NIT4B1 | 321 | 403 | 493 | 187 | 344 | 356 |
| NIT4B2 | 186 | 330 | 314 | 224 | 174 | 191 |
| GST1 | 116 | 54 | 150 | 42 | 106 | 109 |
| GST1B | 134 | 138 | 162 | 132 | 138 | 116 |
| GST3 | 41 | 81 | 69 | 61 | 52 | 58 |
| CAS C1 | 148 | 98 | 247 | 119 | 123 | 132 |
| CAS26 | 125 | 250 | 205 | 156 | 139 | 147 |
| MATE | 222 | 188 | 165 | 188 | 177 | 155 |
| HNL | 342 | 251 | 528 | 437 | 297 | 386 |
| DHR1 | 216 | 329 | 477 | 130 | 206 | 354 |
| DHR2 | 183 | 332 | 429 | 86 | 100 | 110 |
| ACC | 27 | 63 | 70 | 34 | 33 | 75 |
| CM7 | 109 | 265 | 231 | 119 | 119 | 136 |
| Species | Source of Sequence Data | Coding Sequence (bp) | Peptide Sequence (aa) | Intron Length (bp) | |
|---|---|---|---|---|---|
| S. bicolor | database | 1677 | 559 | 97 | Phytozome |
| S. amplum | PCR | 1665 | 555 | 87 | |
| S. angustum | PCR | 1653 | 551 | 94 | |
| S. brachypodum | genome | 1677 | |||
| S. brachypodum | PCR | 1662 | 554 | 99 | |
| S. bulbosum | PCR | 1671 | 557 | 92 | |
| S. ecarinatum | PCR | 1656 | 552 | 82 | |
| S. exstans | PCR | 977 | 338 | - | Partial only |
| S. interjectum | PCR | 1665 | 555 | 81 | |
| S. intrans | PCR | 1665 | 555 | 105 | |
| S. laxiflorum | genome | 1677 | |||
| S. laxiflorum | PCR | 1662 | 554 | 123 | |
| S. leiocladum | genome | 1677 | |||
| S. macrospermum | genome | 1677 | |||
| S. matarankense | genome | 1677 | |||
| S. plumosum | PCR | 1668 | 556 | 195 | |
| S. propinquum | - | No tissue | |||
| S. purpureosericeum | genome | 1677 | |||
| S. purpureosericeum | PCR | 1650 | 550 | 163 | |
| S. stipoideum | - | No amplification | |||
| S. timorense | - | No amplification | |||
| S. versicolor | PCR | 1662 | 554 | 177 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cowan, M.; Møller, B.L.; Norton, S.; Knudsen, C.; Crocoll, C.; Furtado, A.; Henry, R.; Blomstedt, C.; Gleadow, R.M. Cyanogenesis in the Sorghum Genus: From Genotype to Phenotype. Genes 2022, 13, 140. https://doi.org/10.3390/genes13010140
Cowan M, Møller BL, Norton S, Knudsen C, Crocoll C, Furtado A, Henry R, Blomstedt C, Gleadow RM. Cyanogenesis in the Sorghum Genus: From Genotype to Phenotype. Genes. 2022; 13(1):140. https://doi.org/10.3390/genes13010140
Chicago/Turabian StyleCowan, Max, Birger Lindberg Møller, Sally Norton, Camilla Knudsen, Christoph Crocoll, Agnelo Furtado, Robert Henry, Cecilia Blomstedt, and Roslyn M. Gleadow. 2022. "Cyanogenesis in the Sorghum Genus: From Genotype to Phenotype" Genes 13, no. 1: 140. https://doi.org/10.3390/genes13010140
APA StyleCowan, M., Møller, B. L., Norton, S., Knudsen, C., Crocoll, C., Furtado, A., Henry, R., Blomstedt, C., & Gleadow, R. M. (2022). Cyanogenesis in the Sorghum Genus: From Genotype to Phenotype. Genes, 13(1), 140. https://doi.org/10.3390/genes13010140