
Using DNA Metabarcoding to Characterize the Prey Spectrum of Two Co-Occurring Themisto Amphipods in the Rapidly Changing Atlantic-Arctic Gateway Fram Strait
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
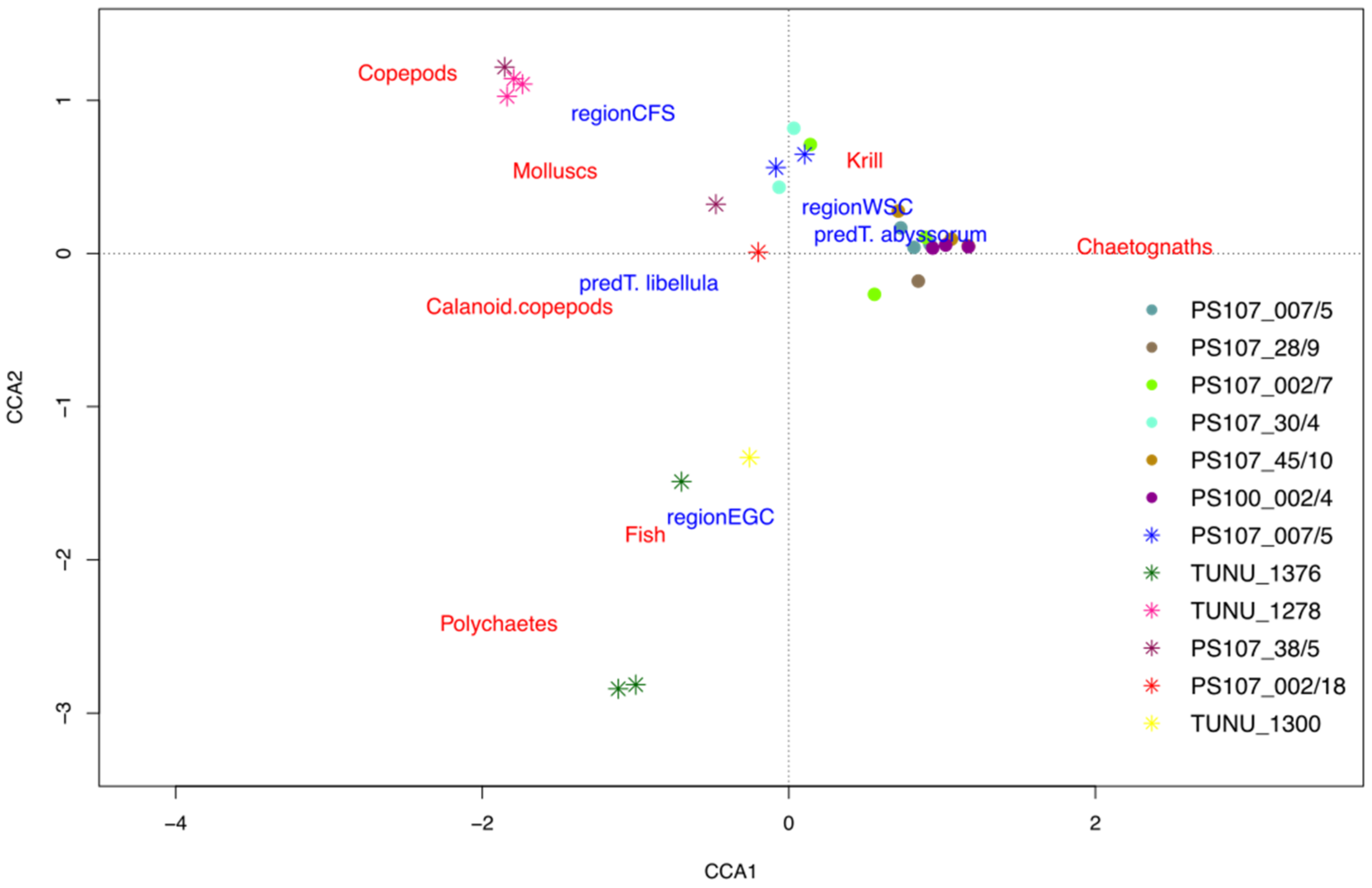
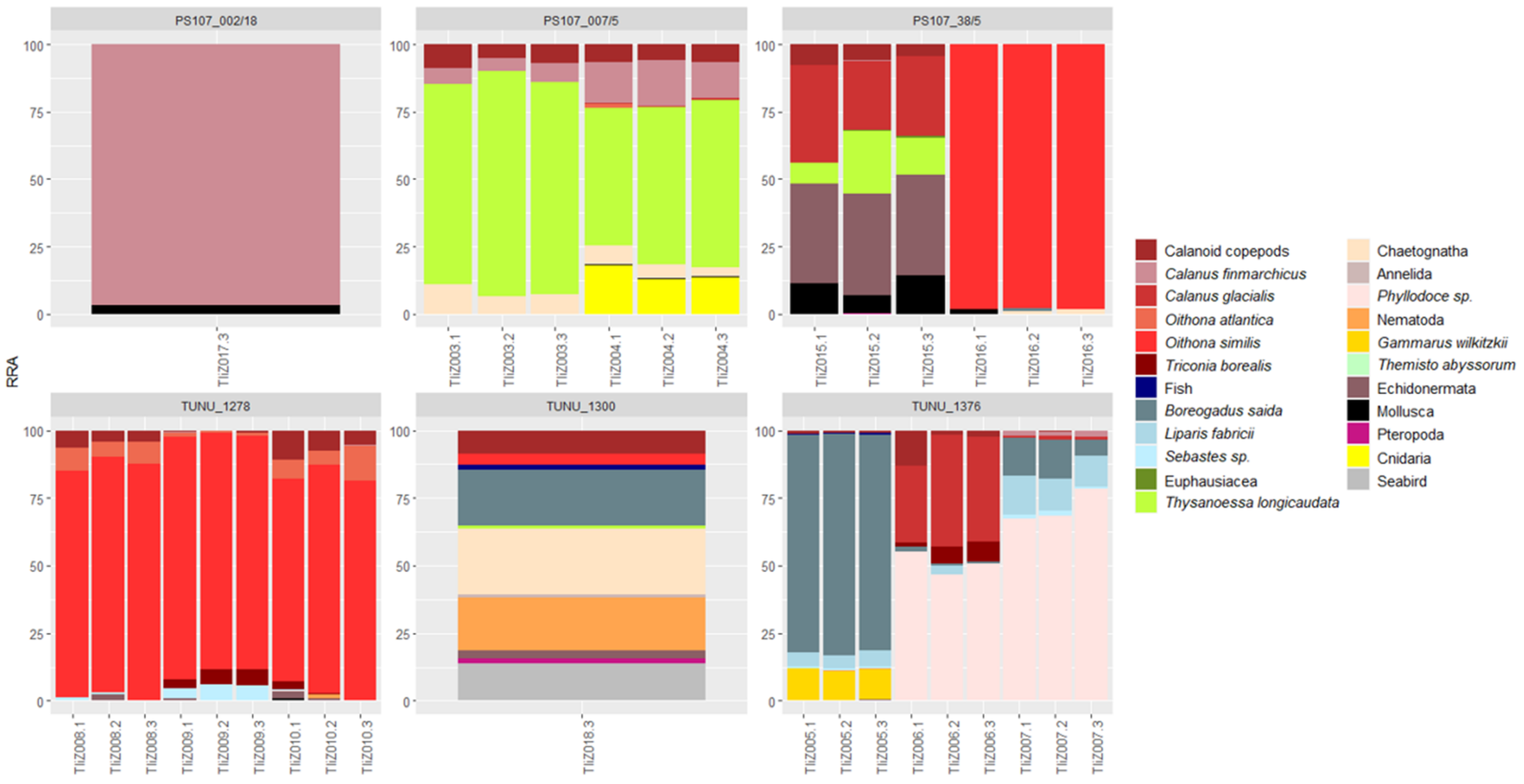
3.1. Interspecific Variation in the Diet Composition of the Two Themisto Species
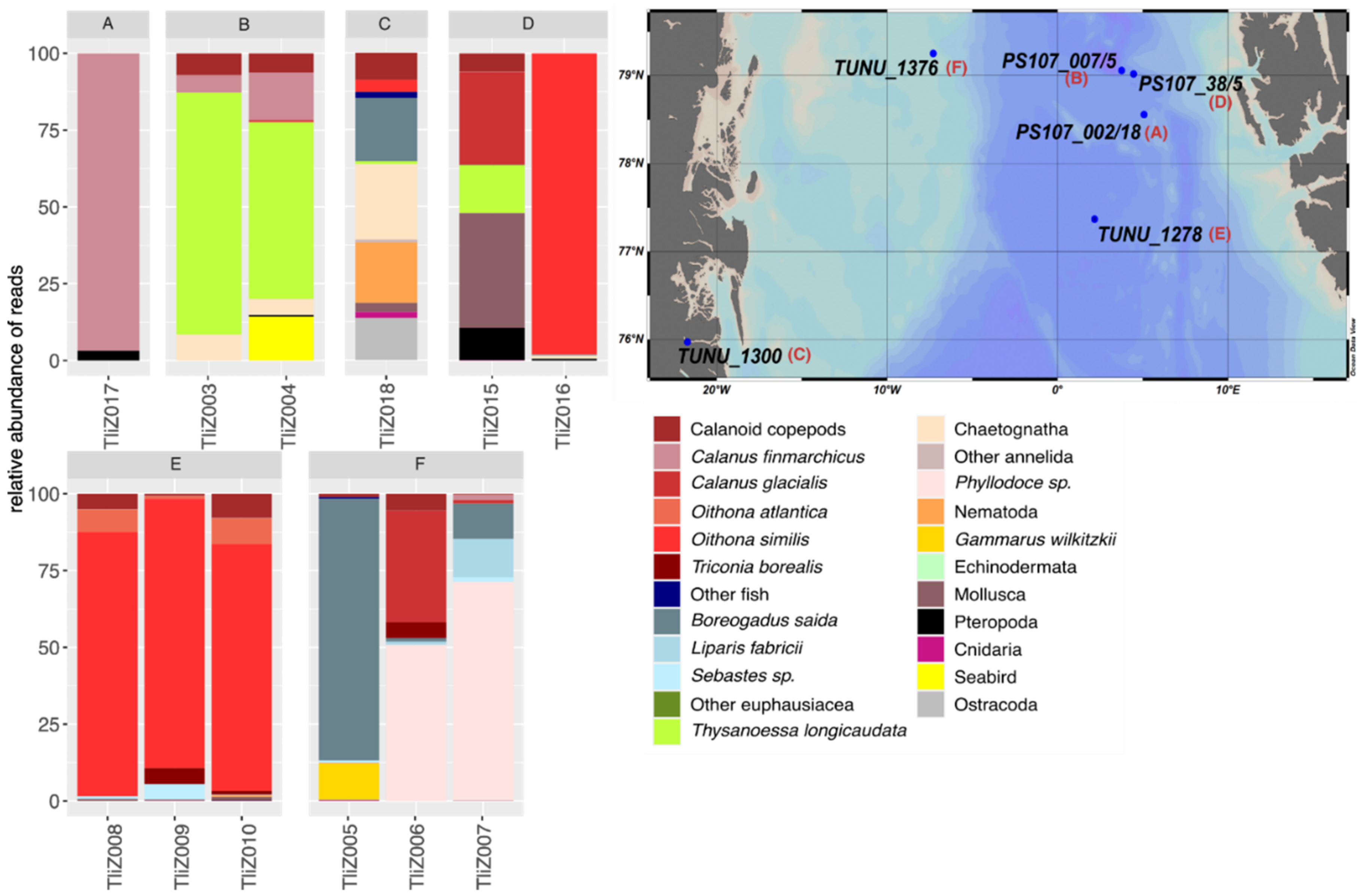
3.2. Intraspecific and Spatial Variation in Prey Composition of T. libellula
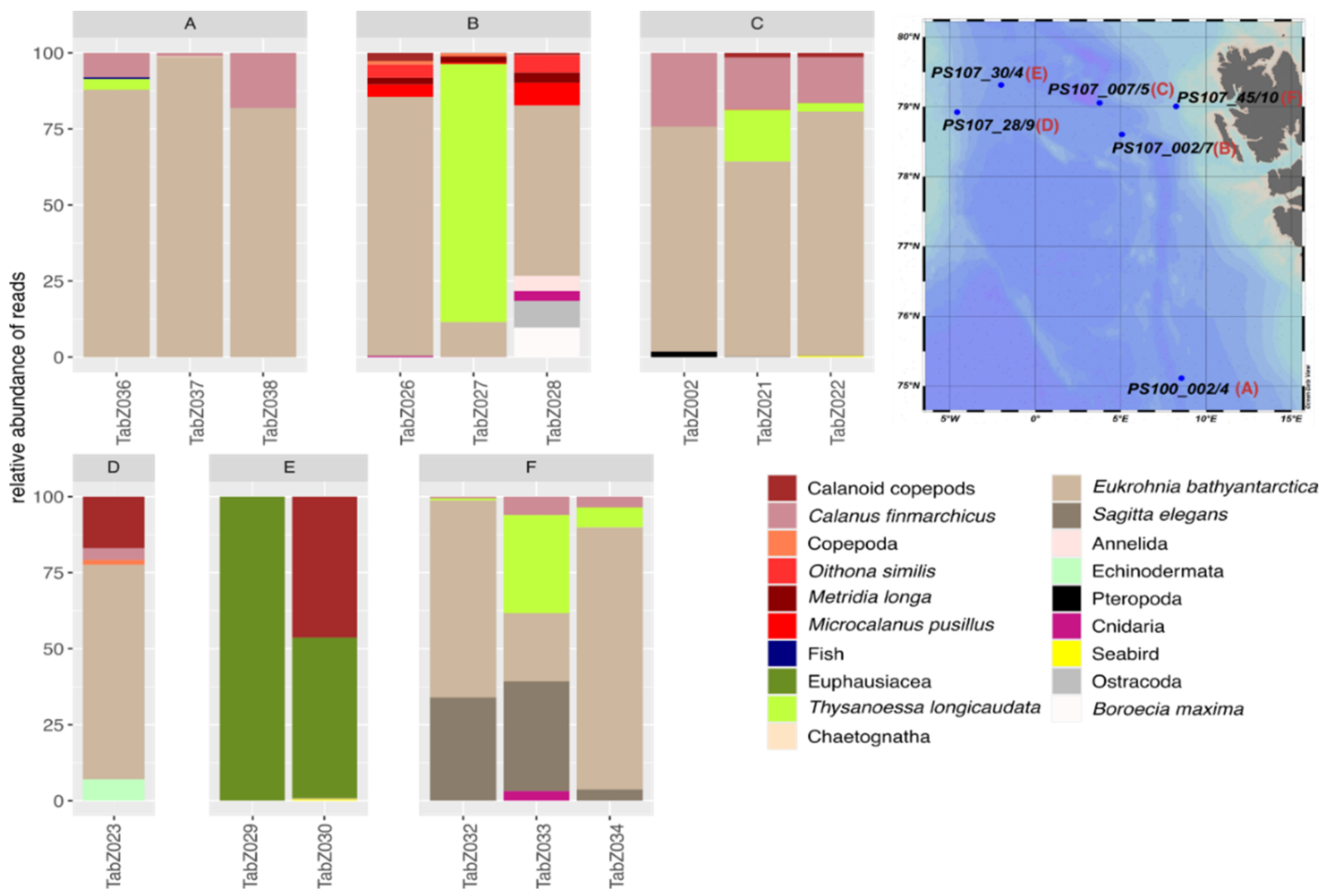
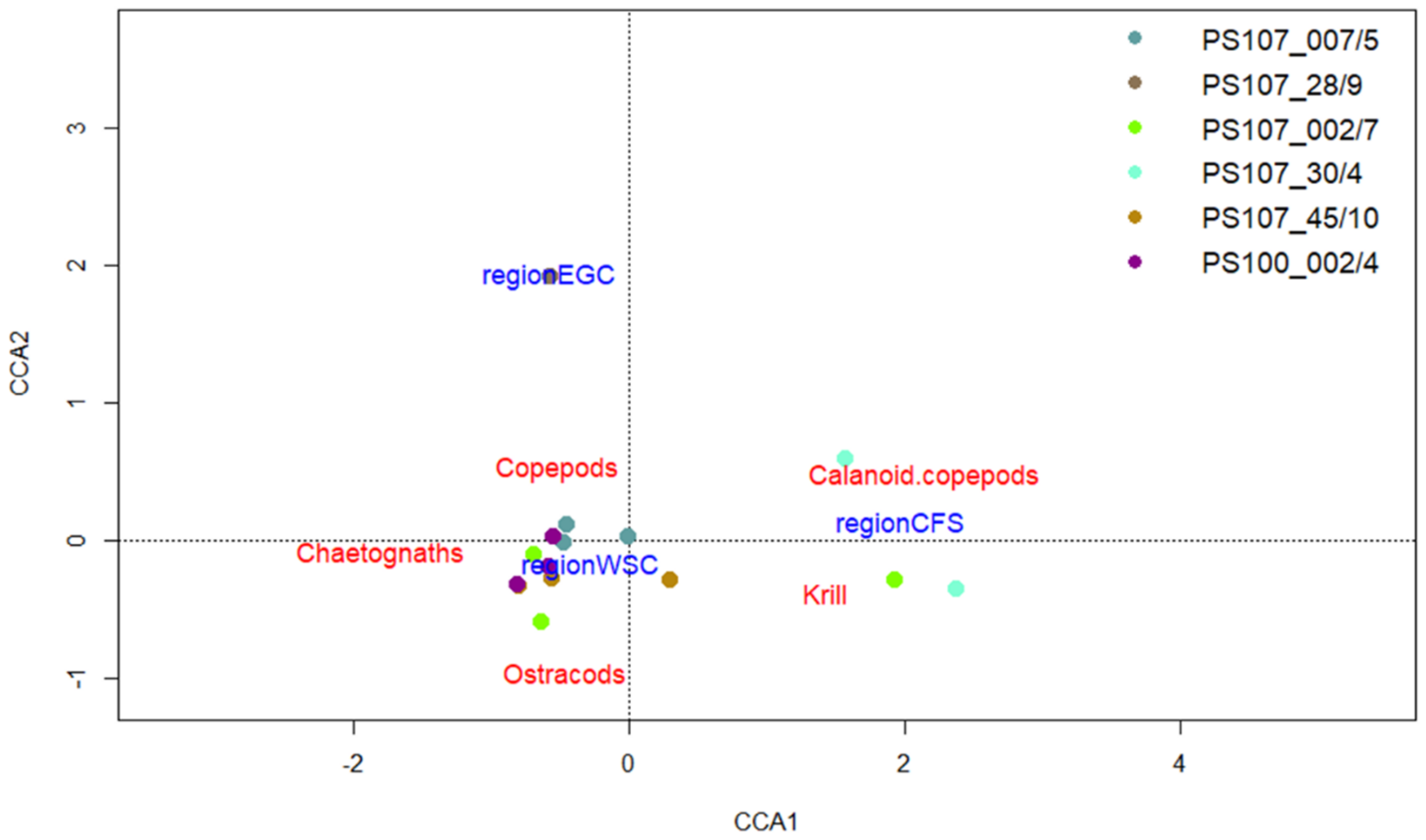
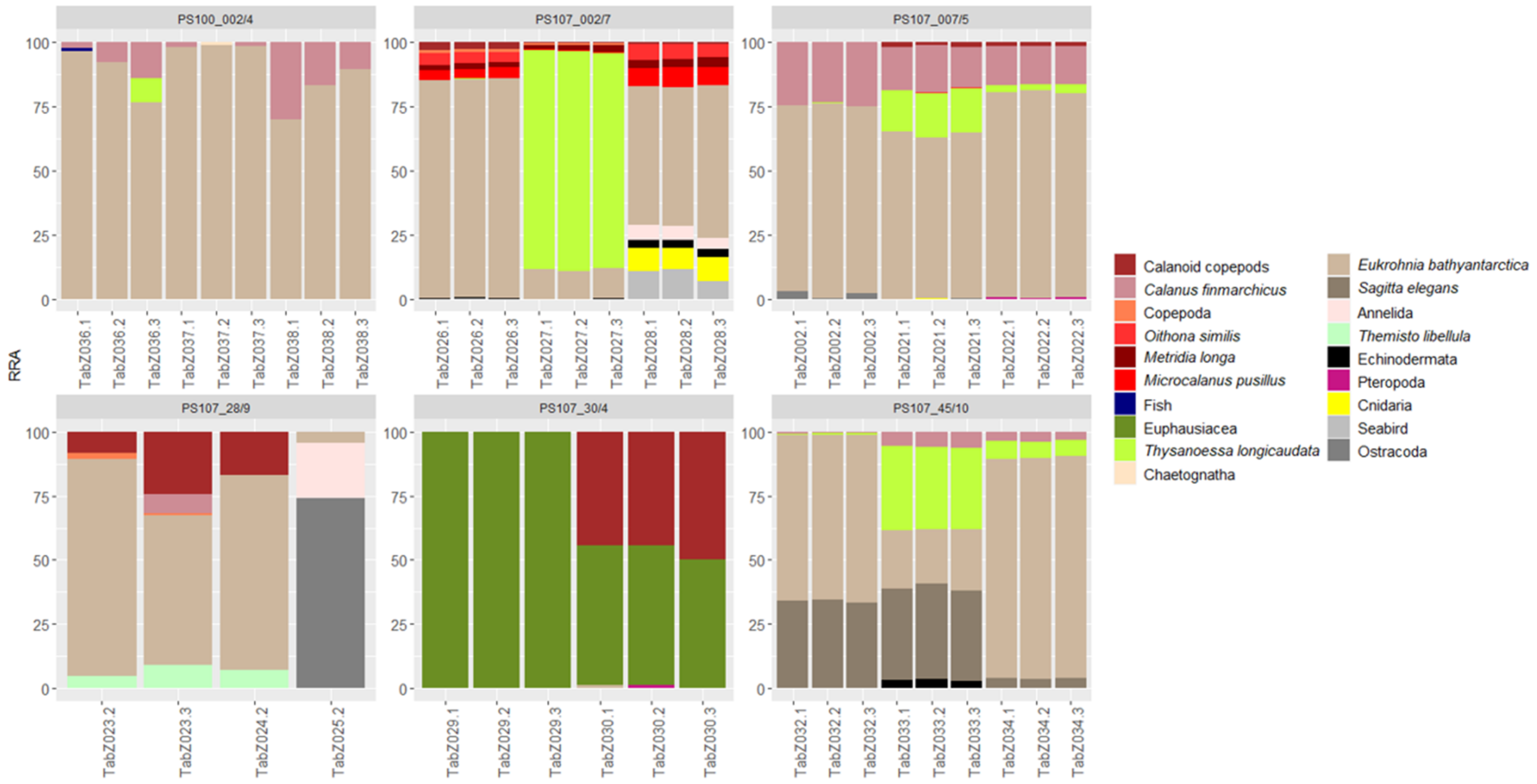
3.3. Intraspecific and Spatial Variation in Prey Composition of T. abyssorum
4. Discussion
4.1. Interspecific Variation in the Diet Composition of the Two Themisto Species
4.2. Intraspecific and Spatial Variation in Prey Composition of T. libellula
4.3. Intraspecific and Spatial Variation in Prey Composition of T. abyssorum
4.4. The Trophic Role of Themisto in a Changing Arctic Ocean
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Carmack, E.; Polyakov, I.; Padman, L.; Fer, I.; Hunke, E.; Hutchings, J.; Jackson, J.; Kelley, D.; Kwok, R.; Layton, C. Toward quantifying the increasing role of oceanic heat in sea ice loss in the new Arctic. Bull. Am. Meteorol. Soc. 2015, 96, 2079–2105. [Google Scholar] [CrossRef]
- Polyakov, V.I.; Walsh, J.E.; Kwok, R. Recent changes of Arctic multiyear sea ice coverage and the likely causes. Bull. Am. Meteorol. Soc. 2012, 93, 145–151. [Google Scholar] [CrossRef]
- Overland, E.J.; Wang, M. When will the summer Arctic be nearly sea ice free? Geophys. Res. Lett. 2013, 40, 2097–2101. [Google Scholar] [CrossRef]
- Barry, R.G.; Serreze, M.C.; Maslanik, J.A.; Preller, R.H. The Arctic sea ice-climate system: Observations and modeling. Rev. Geophys. 1993, 31, 397–422. [Google Scholar] [CrossRef]
- Lønne, O.; Gulliksen, B. On the distribution of sympagic macro-fauna in the seasonally ice covered Barents Sea. Polar Biol. 1991, 11, 457–469. [Google Scholar]
- Lønne, O.; Gulliksen, B. Sympagic macro-fauna from multiyear sea-ice near Svalbard. Polar Biol. 1991, 11, 471–477. [Google Scholar]
- Falk-Petersen, S.; Mayzaud, P.; Kattner, G.; Sargent, J.R. Lipids and life strategy of Arctic Calanus. Mar. Biol. Res. 2009, 5, 18–39. [Google Scholar] [CrossRef] [Green Version]
- Kohlbach, D.; Graeve, M.; Lange, A.B.; David, C.; Peeken, I.; Flores, H. The importance of ice algae-produced carbon in the central Arctic Ocean ecosystem: Food web relationships revealed by lipid and stable isotope analyses. Limnol. Oceanogr. 2016, 61, 2027–2044. [Google Scholar] [CrossRef]
- Søreide, J.E.; Carroll, M.L.; Hop, H.; Ambrose, W.G., Jr.; Hegseth, E.N.; Falk-Petersen, S. Sympagic-pelagic-benthic coupling in Arctic and Atlantic waters around Svalbard revealed by stable isotopic and fatty acid tracers. Mar. Biol. Res. 2013, 9, 831–850. [Google Scholar] [CrossRef]
- Søreide, J.E.; Hop, H.; Carroll, M.L.; Falk-Petersen, S.; Hegseth, E.N. Seasonal food web structures and sympagic–pelagic coupling in the European Arctic revealed by stable isotopes and a two-source food web model. Prog. Oceanogr. 2006, 71, 59–87. [Google Scholar] [CrossRef]
- Haug, T.; Bogstad, B.; Chierci, M.; Gjøsæter, H.; Hallfredsson, E.H.; Høines, A.S.; Hoel, A.H.; Ingvaldsen, R.B.; Jørgensen, L.L.; Knutsen, T. Future harvest of living resources in the Arctic Ocean north of the Nordic and Barents Seas: A review of possibilities and constraints. Fish. Res. 2017, 188, 38–57. [Google Scholar] [CrossRef]
- Søreide, J.E.; Leu, E.V.A.; Berge, J.; Graeve, M.; Falk-Petersen, S. Timing of blooms, algal food quality and Calanus glacialis reproduction and growth in a changing Arctic. Glob. Chang. Biol. 2010, 16, 3154–3163. [Google Scholar] [CrossRef]
- Ehrlich, J.; Schaafsma, F.L.; Bluhm, B.A.; Peeken, I.; Castellani, G.; Brandt, A.; Flores, H. Sympagic fauna in and under Arctic pack ice in the annual sea-ice system of the new Arctic. Front. Mar. Sci. 2020, 7, 452. [Google Scholar] [CrossRef]
- Wadhams, P. Sea ice thickness distribution in Fram Strait. Nature 1983, 305, 108–111. [Google Scholar] [CrossRef]
- Hop, H.; Falk-Petersen, S.; Svendsen, H.; Kwasniewski, S.; Pavlov, V.; Pavlova, O.; Søreide, J.E. Physical and biological characteristics of the pelagic system across Fram Strait to Kongsfjorden. Prog. Oceanogr. 2006, 71, 182–231. [Google Scholar] [CrossRef]
- Polyakov, I.V.; Pnyushkov, A.V.; Alkire, M.B.; Ashik, I.M.; Baumann, T.M.; Carmack, E.C.; Goszczko, I.; Guthrie, J.; Ivanov, V.V.; Kanzow, T. Greater role for Atlantic inflows on sea-ice loss in the Eurasian Basin of the Arctic Ocean. Science 2017, 356, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Basedow, S.L.; Sundfjord, A.; Von Appen, W.-J.; Halvorsen, E.; Kwasniewski, S.; Reigstad, M. Seasonal variation in transport of zooplankton into the Arctic basin through the Atlantic gateway, Fram Strait. Front. Mar. Sci. 2018, 5, 194. [Google Scholar] [CrossRef] [Green Version]
- Polyakov, I.V.; Alkire, M.B.; Bluhm, B.A.; Brown, K.A.; Carmack, E.C.; Chierci, M.; Danielson, S.L.; Ellingsen, I.; Ershova, E.A.; Gårdfeldt, K. Borealization of the Arctic Ocean in response to anomalous advection from sub-Arctic seas. Front. Mar. Sci. 2020, 7, 491. [Google Scholar] [CrossRef]
- Andrews, A.J.; Christiansen, J.S.; Bhat, S.; Lynghammar, A.; Westgaard, J.-I.; Pampoulie, C.; Præbel, K. Boreal marine fauna from the Barents Sea disperse to Arctic Northeast Greenland. Sci. Rep. 2019, 9, 5799. [Google Scholar] [CrossRef] [Green Version]
- Longhurst, A.R. The structure and evolution of plankton communities. Prog. Oceanogr. 1985, 15, 1–35. [Google Scholar] [CrossRef]
- Bowman, T.E. The pelagic amphipod genus Parathemisto (Hyperiidea: Hyperiidae) in the north Pacific and adjacent Arctic Ocean. Proc. U.S. Natl. Mus. 1960, 112, 343–392. [Google Scholar] [CrossRef] [Green Version]
- Dalpadado, P.; Borkner, N.; Skjoldal, H.R. Distribution and Life History of Themisti (Amphipoda) spp., North of 73° N in the Barents Sea. 1994. Available online: https://imr.brage.unit.no/imr-xmlui/handle/11250/113407?locale-attribute=en (accessed on 1 March 2020).
- Havermans, C.; Auel, H.; Hagen, W.; Held, C.; Ensor, N.S.; Tarling, G.A. Predatory zooplankton on the move: Themisto amphipods in high-latitude marine pelagic food webs. Adv. Mar. Biol. 2019, 82, 51–92. [Google Scholar] [PubMed]
- Auel, H.; Harjes, M.; Da Rocha, R.; Stübing, D.; Hagen, W. Lipid biomarkers indicate different ecological niches and trophic relationships of the Arctic hyperiid amphipods Themisto abyssorum and T. libellula. Polar Biol. 2002, 25, 374–383. [Google Scholar] [CrossRef]
- Conservation of Arctic Flora and Fauna. State of the Arctic Marine Biodiversity Report; Conservation of Arctic Flora and Fauna: Akureyri, Iceland, 2017. [Google Scholar]
- Pakhomov, E.; Perissinotto, R. Trophodynamics of the hyperiid amphipod Themisto gaudichaudi in the South Georgia region during late austral summer. Mar. Ecol. Prog. Ser. 1996, 134, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Olsen, B.R.; Troedsson, C.; Hadziavdic, K.; Pedersen, R.B.; Rapp, H.T. A molecular gut content study of Themisto abyssorum (Amphipoda) from Arctic hydrothermal vent and cold seep systems. Mol. Ecol. 2014, 23, 3877–3889. [Google Scholar] [CrossRef]
- Arai, M.N. Pelagic Coelenterates and Eutrophication: A Review. Hydrobiologia 2001, 451, 69–87. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Hajibabaei, M.; Rieseberg, L.H. Environmental DNA; Wiley Online Library: Hoboken, NJ, USA, 2012; pp. 1789–1793. [Google Scholar]
- Urban, P.; Præbel, K.; Bhat, S.; Dierking, J.; Wangensteen, O.S. DNA metabarcoding reveals the importance of gelatinous zooplankton in the diet of Pandalus borealis, a keystone species in the Arctic. Mol. Ecol. 2022, 31, 1562–1576. [Google Scholar] [CrossRef]
- Knust, R. Polar research and supply vessel POLARSTERN operated by the Alfred-Wegener-Institute. J. Large Scale Res. Facil. 2017, 3, A119. [Google Scholar] [CrossRef]
- Auel, H.; Havermans, C.; Melo Franco Santos, R.; Prinz, N.; Jacinto, R.; Boavida, J. AMICA—Arctic Marginal Ice Zone Community Assessment: Biodiversity, productivity and trophic interactions in the marginal ice zone of Fram Strait under global change. In The Expedition PS100 of the Research Vessel POLARSTERN to the Fram Strait in 2016. Berichte zur Polar- und Meeresforschung; Kanzow, T., Ed.; Alfred Wegener Institute for Polar and Marine Research: Bremerhaven, Germany, 2016; pp. 125–132. [Google Scholar]
- Auel, H.; Havermans, C.; Zelm, R.; Köhler, E.; Scheel, M. Sensitivity of Arctic zooplankton to temperature change: From ecophysiological effects to community repsonse. In The Expedition PS107 of the Research Vessel POLARSTERN to the Arctic in 2017. Berichte zur Polar- und Meeresforschung; Schewe, I., Ed.; Alfred Wegener Institute for Polar and Marine Research: Bremerhaven, Germany, 2018; pp. 72–75. [Google Scholar]
- Qiagen. DNeasy Powersoil Kit Handbook-for the Isolation of Microbial Genomic DNA from all Soil Samples. 2017. Available online: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&ved=2ahUKEwjjnb2kror7AhW1g_0HHUj2AzQQFnoECBYQAQ&url=https%3A%2F%2Fwww.qiagen.com%2Fcn%2Fresources%2Fdownload.aspx%3Fid%3D5a0517a7-711d-4085-8a28-2bb25fab828a%26lang%3Den&usg=AOvVaw1Q_l8pjSpR7NeXPkmGYA3x (accessed on 3 June 2020).
- Siegenthaler, A.; Wangensteen, O.S.; Benvenuto, C.; Campos, J.; Mariani, S. DNA metabarcoding unveils multiscale trophic variation in a widespread coastal opportunist. Mol. Ecol. 2019, 28, 232–249. [Google Scholar] [CrossRef] [Green Version]
- Leray, M.; Yang, J.Y.; Meyer, C.P.; Mills, S.C.; Agudelo, N.; Ranwez, V.; Boehm, J.T.; Machida, R.J. A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity: Application for characterizing coral reef fish gut contents. Front. Zool. 2013, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wangensteen, O.S.; Palacín, C.; Guardiola, M.; Turon, X. DNA metabarcoding of littoral hard-bottom communities: High diversity and database gaps revealed by two molecular markers. PeerJ 2018, 6, e4705. [Google Scholar] [CrossRef]
- Geller, J.; Meyer, C.; Parker, M.; Hawk, H. Redesign of PCR primers for mitochondrial cytochrome c oxidase subunit I for marine invertebrates and application in all-taxa biotic surveys. Mol. Ecol. Resour. 2013, 13, 851–861. [Google Scholar] [CrossRef]
- Brown, J.; Pirrung, M.; McCue, L.A. FQC Dashboard: Integrates FastQC results into a web-based, interactive, and extensible FASTQ quality control tool. Bioinformatics 2017, 33, 3137–3139. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, 2584. [Google Scholar] [CrossRef] [Green Version]
- Mahé, F.; Rognes, T.; Quince, C.; De Vargas, C.; Dunthorn, M. Swarm v2: Highly-scalable and high-resolution amplicon clustering. PeerJ 2015, 3, e1420. [Google Scholar] [CrossRef] [Green Version]
- Boyer, F.; Mercier, C.; Bonin, A.; Le Bras, Y.; Taberlet, P.; Coissac, E. obitools: A unix-inspired software package for DNA metabarcoding. Mol. Ecol. Resour. 2016, 16, 176–182. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Deagle, B.E.; Thomas, A.C.; McInnes, J.C.; Clarke, L.J.; Vesterinen, E.J.; Clare, E.L.; Katzinel, T.R.; Eveson, J.P. Counting with DNA in metabarcoding studies: How should we convert sequence reads to dietary data? Mol. Ecol. 2019, 28, 391–406. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package; R Package Version 2.5-7. 2020. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 15 March 2021).
- Goral, F.; Schellenberg, J. goeveg: Function for Community Data and Ordinations; R Package Version 0.5.1. 2021. Available online: https://cran.r-project.org/web/packages/goeveg/goeveg.pdf (accessed on 15 March 2021).
- Dalpadado, P.; Yamaguchi, A.; Ellertsen, B.; Johannessen, S. Trophic interactions of macro-zooplankton (krill and amphipods) in the Marginal Ice Zone of the Barents Sea. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 2266–2274. [Google Scholar] [CrossRef]
- Kraft, A.; Berge, J.; Varpe, Ø.; Falk-Petersen, S. Feeding in Arctic darkness: Mid-winter diet of the pelagic amphipods Themisto abyssorum and T. libellula. Mar. Biol. 2013, 160, 241–248. [Google Scholar] [CrossRef]
- Dalpadado, P.; Borkner, N.; Bogstad, B.; Mehl, S. Distribution of Themisto (Amphipoda) spp. in the Barents Sea and predator-prey interactions. ICES J. Mar. Sci. 2001, 58, 876–895. [Google Scholar] [CrossRef]
- Vinogradov, G.M. Deep-sea near-bottom swarms of pelagic amphipods Themisto: Observations from submersibles. Sarsia 1999, 84, 465–467. [Google Scholar] [CrossRef]
- Berge, J.; Nahrgang, J. The Atlantic spiny lumpsucker Eumicrotremus spinosus: Life history traits and the seemingly unlikely interaction with the pelagic amphipod Themisto libellula. Pol. Polar Res. 2013, 34, 279–287. [Google Scholar] [CrossRef]
- Kruse, S. Population structure and reproduction of Eukrohnia bathypelagica and Eukrohnia bathyantarctica in the Lazarev Sea, Southern Ocean. Polar Biol. 2009, 32, 1377–1387. [Google Scholar] [CrossRef]
- Zheng, L.; He, J.; Lin, Y.; Cao, W.; Zhang, W. 16S rRNA is a better choice than COI for DNA barcoding hydrozoans in the coastal waters of China. Acta Oceanol. Sin. 2014, 33, 55–76. [Google Scholar] [CrossRef]
- Günther, B.; Fromentin, J.-M.; Metral, L.; Arnaud-Haond, S. Metabarcoding confirms the opportunistic foraging behaviour of Atlantic bluefin tuna and reveals the importance of gelatinous prey. PeerJ 2021, 9, e11757. [Google Scholar] [CrossRef]
- Günther, B.; Knebelsberger, T.; Neumann, H.; Laakmann, S.; Martínez Arbizu, P. Metabarcoding of marine environmental DNA based on mitochondrial and nuclear genes. Sci. Rep. 2018, 8, 14822. [Google Scholar] [CrossRef] [Green Version]
- Clarke, L.J.; Beard, J.M.; Swadling, K.M.; Deagle, B.E. Effect of marker choice and thermal cycling protocol on zooplankton DNA metabarcoding studies. Ecol. Evol. 2017, 7, 873–883. [Google Scholar] [CrossRef]
- Tang, C.Q.; Leasi, F.; Obertegger, U.; Kieneke, A.; Barraclough, T.G.; Fontaneto, D. The widely used small subunit 18S rDNA molecule greatly underestimates true diversity in biodiversity surveys of the meiofauna. Proc. Natl. Acad. Sci. USA 2012, 109, 16208–16212. [Google Scholar] [CrossRef] [Green Version]
- Tempestini, A.; Fortier, L.; Pinchuk, A.; Dufresne, F. Molecular phylogeny of the genus Themisto (Guérin, 1925)(Amphipoda: Hyperiidae) in the northern hemisphere. J. Crustac. Biol. 2017, 37, 732–742. [Google Scholar] [CrossRef]
- Williamson, D. Note on Themisto gracilipes Norman; Report of Marine Biological Station at Port Erin, Isel of Man; 1950; p. 62. [Google Scholar]
- Fetzer, I.; Deubel, H. Effect of river run-off on the distribution of marine invertebrate larvae in the southern Kara Sea (Russian Arctic). J. Mar. Syst. 2006, 60, 98–114. [Google Scholar] [CrossRef]
- Sejr, M.K.; Jensen, K.T.; Rysgaard, S. Macrozoobenthic community structure in a high-arctic East Greenland fjord. Polar Biol. 2000, 23, 792–801. [Google Scholar] [CrossRef]
- Bodil, B.A.; Ambrose, W.G.; Bergmann, M.; Clough, L.M.; Gebruk, A.V.; Hasemann, C.; Iken, K.; Klages, M.; MacDonald, I.R.; Renaud, P.E. Diversity of the arctic deep-sea benthos. Mar. Biodivers. 2011, 41, 87–107. [Google Scholar] [CrossRef]
- Hirche, H.-J.; Kosobokova, K. Distribution of Calanus finmarchicus in the northern North Atlantic and Arctic Ocean—expatriation and potential colonization. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2007, 54, 2729–2747. [Google Scholar] [CrossRef]
- Freer, J.; Daase, M.; Tarling, G.A. Modelling the biogeographic boundary shift of Calanus finmarchicus reveals drivers of Arctic Atlantification by subarctic zooplankton. Glob. Chang. Biol. 2021, 28, 429–440. [Google Scholar]
- Huserbråten, M.B.O.; Eriksen, E.; Gjøsæter, H.; Vikebø, F. Polar cod in jeopardy under the retreating Arctic sea ice. Commun. Biol. 2019, 2, 407. [Google Scholar] [CrossRef] [Green Version]
- Nahrgang, J.; Varpe, Ø.; Korshunova, E.; Murzina, S.; Hallanger, I.G.; Vieweg, I.; Berge, J. Gender specific reproductive strategies of an Arctic key species (Boreogadus saida) and implications of climate change. PLoS ONE 2014, 9, e98452. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
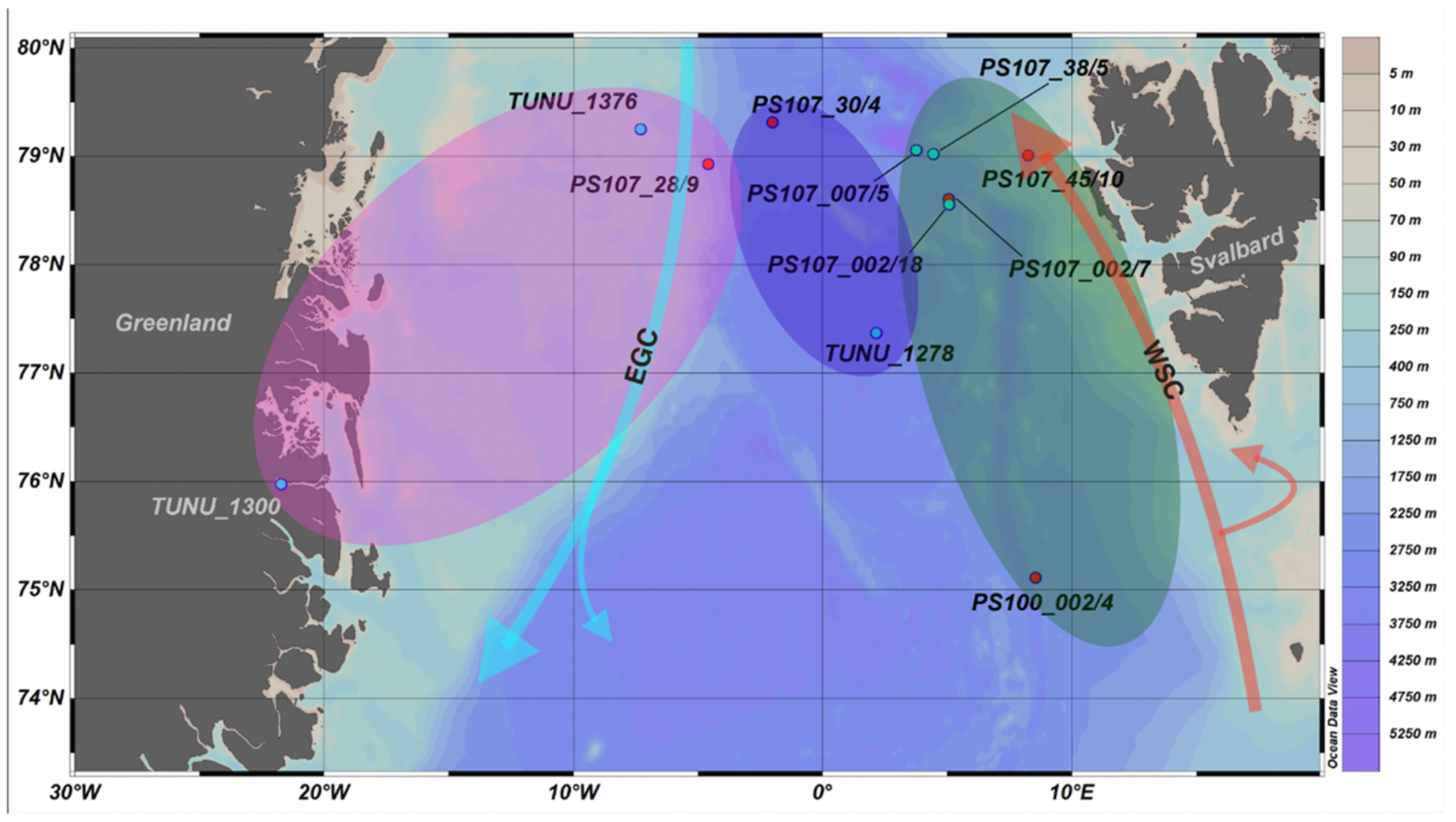
| Station | Species | Size [mm] | Latitude [°N] | Longitude | Gear | Depth [m] | Date |
|---|---|---|---|---|---|---|---|
| PS107_007/5 | T. libellula | 10–20 | 79.06 | 3.75 E | BO | 450 | 29 July 2017 |
| TUNU_1376 | 10–20 | 79.25 | 7.31 W | PT | 20 | 24 September 2017 | |
| TUNU_1278 | 10–20 | 77.37 | 2.15 E | PT | 31 | 15 September 2017 | |
| TUNU_1300 | >20 | 75.97 | 21.72 W | CT | 230 | 18 September 2017 | |
| PS107_002/18 | 10–20 | 78.56 | 5.06 E | BO | 60 | 26 July 2017 | |
| PS107_38/5 | 10–20 | 79.02 | 4.45 E | BO | 60 | 11 August 2017 | |
| PS107_007/5 | T. abyssorum | <10 | 79.06 | 3.75 E | BO | 450 | 29 July 2017 |
| PS107_28/9 | < 10 | 78.93 | 4.58 W | BO | 450 | 5 August 2017 | |
| PS107_002/7 | <10 | 78.61 | 5.06 E | MN | NA | 26 July 2017 | |
| PS107_30/4 | 10–20 | 79.32 | 2.01 W | BO | 450 | 6 August 2017 | |
| PS107_45/10 | <10 | 79.01 | 8.23 E | BO | 450 | 14 August 2017 | |
| PS100_002/4 | <10 | 75.11 | 8.54 E | BO | NA | 20 July 2016 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dischereit, A.; Wangensteen, O.S.; Præbel, K.; Auel, H.; Havermans, C. Using DNA Metabarcoding to Characterize the Prey Spectrum of Two Co-Occurring Themisto Amphipods in the Rapidly Changing Atlantic-Arctic Gateway Fram Strait. Genes 2022, 13, 2035. https://doi.org/10.3390/genes13112035
Dischereit A, Wangensteen OS, Præbel K, Auel H, Havermans C. Using DNA Metabarcoding to Characterize the Prey Spectrum of Two Co-Occurring Themisto Amphipods in the Rapidly Changing Atlantic-Arctic Gateway Fram Strait. Genes. 2022; 13(11):2035. https://doi.org/10.3390/genes13112035
Chicago/Turabian StyleDischereit, Annkathrin, Owen S. Wangensteen, Kim Præbel, Holger Auel, and Charlotte Havermans. 2022. "Using DNA Metabarcoding to Characterize the Prey Spectrum of Two Co-Occurring Themisto Amphipods in the Rapidly Changing Atlantic-Arctic Gateway Fram Strait" Genes 13, no. 11: 2035. https://doi.org/10.3390/genes13112035
APA StyleDischereit, A., Wangensteen, O. S., Præbel, K., Auel, H., & Havermans, C. (2022). Using DNA Metabarcoding to Characterize the Prey Spectrum of Two Co-Occurring Themisto Amphipods in the Rapidly Changing Atlantic-Arctic Gateway Fram Strait. Genes, 13(11), 2035. https://doi.org/10.3390/genes13112035