
Polygenic Risk for Schizophrenia Has Sex-Specific Effects on Brain Activity during Memory Processing in Healthy Individuals


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Working Memory Task
2.3. Episodic Memory Task
2.4. MRI Acquisition
2.5. fMRI Analyses
2.6. Genotyping and Construction of Polygenic Risk Scores
2.7. Statistical Analyses
3. Results
3.1. Behavioral Results
3.2. Memory-Related Brain Activation
3.3. Sex-Specific Effects of Schizophrenia Genetics on Functional Brain Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The World Health Organization. The Global Burden of Disease: 2004 Update; WHO Press: Geneva, Switzerland, 2008. [Google Scholar]
- Young, J.W.; Geyer, M.A.; Diego, S. Developing treatments for cognitive deficits in schizophrenia: The challenge of translation. J. Psychopharmacol. 2015, 29, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Ragland, J.; Carter, C. Memory and Cognition in Schizophrenia. Mol. Psychiatry 2019, 24, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Fusar-Poli, P.; Deste, G.; Smierskova, R.; Barlati, S.; Yung, A.; Howes, O.; Stierglitz, R.-D.; Vita, A.; McGuire, P.; Borgwardt, S. Cognitive Functioning in Prodromal Psychosis. Arch. Gen. Psychiatry 2020, 69, 562–571. [Google Scholar] [CrossRef] [PubMed]
- White, T.; Schmidt, M.; Karatekin, C. Verbal and visuospatial working memory development and deficits in children and adolescents with schizophrenia. Early Interv. Psychiatry 2010, 4, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Gold, J.M.; Robinson, B.; Leonard, C.J.; Hahn, B.; Chen, S.; Mcmahon, R.P.; Luck, S.J. Selective Attention, Working Memory, and Executive Function as Potential Independent Sources of Cognitive Dysfunction in Schizophrenia. Schizophr. Bull. 2018, 44, 1227–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achim, A.; Lepage, M. Episodic memory-related activation in schizophrenia: Meta-analysis. Br. J. Psychiatry 2005, 187, 500–509. [Google Scholar] [CrossRef] [Green Version]
- Tsuang, M.; Lyons, M.; Faaone, S. Heterogeneity of Schizophrenia: Conceptual Models and Analytic Strategies. Br. J. Psychiatry 1990, 156, 17–26. [Google Scholar] [CrossRef]
- Sullivan, P.F.; Kendler, K.S.; Neale, M.C. Schizophrenia as a Complex Trait. Arch. Gen. Psychiatry 2003, 60, 1187–1192. [Google Scholar] [CrossRef] [Green Version]
- Ripke, S.; Neale, B.M.; Corvin, A.; Walters, J.T.R.; Farh, K.-H.; Lee, P.; Bulik-Sullivan, B.; Collier, D.A. Biological Insights From 108 Schizophrenia-Associated Genetic Loci. Nature 2014, 511, 421–427. [Google Scholar]
- Purcell, S.; Wray, N.R.; Stone, J.L.; Visscher, P.M.; O’Donovan, M.C.; Sullivan, P.F.; Sklar, P. Common polygenic variation contributes to risk of schizophremia that overlaps with bipolar disease. Nature 2009, 460, 748–752. [Google Scholar]
- Choi, S.W.; Mak, T.S.H.; O’Reilly, P.F. Tutorial: A guide to performing polygenic risk score analyses. Nat. Protoc. 2020, 15, 2759–2772. [Google Scholar] [CrossRef]
- Dudbridge, F. Power and Predictive Accuracy of Polygenic Risk Scores. PLoS Genet. 2013, 9, e1003348. [Google Scholar] [CrossRef]
- Mallet, J.; Le Strat, Y.; Dubertret, C.; Gorwood, P. Polygenic Risk Scores Shed Light on the Relationship between Schizophrenia and Cognitive Functioning: Review and Meta-Analysis. J. Clin. Med. 2020, 9, 341. [Google Scholar] [CrossRef] [Green Version]
- Kępińska, A.P.; MacCabe, J.H.; Cadar, D.; Steptoe, A.; Murray, R.M.; Ajnakina, O. Schizophrenia polygenic risk predicts general cognitive deficit but not cognitive decline in healthy older adults. Transl. Psychiatry 2020, 10, 422. [Google Scholar] [CrossRef]
- Koch, E.; Nyberg, L.; Lundquist, A.; Pudas, S.; Adolfsson, R.; Kauppi, K. Sex-specific effects of polygenic risk for schizophrenia on lifespan cognitive functioning in healthy individuals. Transl. Psychiatry 2021, 11, 520. [Google Scholar] [CrossRef]
- Häfner, H.; Maurer, K.; Löffler, W.; Riecher-Rössler, A. The influence of age and sex on the onset of early course of schizophrenia. Br. J. Psychiatry 1993, 162, 80–86. [Google Scholar] [CrossRef]
- Moriarty, P.J.; Lieber, D.; Bennett, A.; White, L.; Parrella, M.; Harvey, P.D.; Davis, K.L. Gender differences in poor outcome patients with lifelong schizophrenia. Schizophr. Bull. 2001, 27, 103–113. [Google Scholar] [CrossRef]
- Rubin, L.; Haas, G.; Keshavan, M.; Sweeney, J.; Maki, P. Sex Difference in Cognitive Response to Antipsychotic Treatment in First Episode Schizophrenia. Neuropsychopharmacology 2008, 33, 290–297. [Google Scholar] [CrossRef]
- Castle, D.J.; Wessely, S.; Murray, R.M. Sex and schizophrenia: Effects of diagnostic stringency, and associations with premorbid variables. Br. J. Psychiatry 1993, 162, 658–664. [Google Scholar] [CrossRef]
- Kraepelin, E. Dementia Praecox and Paraphrenia; Robert, E. Krieger Publishing Co.: Huntington, NY, USA, 1971. [Google Scholar]
- Jongsma, H.E.; Turner, C.; Kirkbride, J.B.; Jones, P.B. International incidence of psychotic disorders, 2002–2017: A systematic review and meta-analysis. Lancet Public Health 2019, 4, e229–e244. [Google Scholar] [CrossRef] [Green Version]
- Seidman, L.J.; Goldstein, J.M.; Goodman, J.M.; Koren, D.; Turner, W.M.; Faraone, S.V.; Tsuang, M.T. Sex differences in olfactory identification and Wisconsin card sorting performance in schizophrenia: Relationship to attention and verbal ability. Biol. Psychiatry 1997, 42, 104–115. [Google Scholar] [CrossRef]
- Goldstein, J.M.; Seidman, L.J.; Goodman, J.M.; Koren, D.; Lee, H.; Weintraub, S.; Tsuang, M.T. Are There Sex Differences in Neuropsychological Functions Among Patients With Schizophrenia? Am. J. Psychiatry 1998, 155, 1358–1364. [Google Scholar] [CrossRef] [PubMed]
- Sota, T.L.; Heinrichs, R.W. Sex differences in verbal memory in schizophrenia patients treated with “typical” neuroleptics. Schizophr. Res. 2003, 62, 175–182. [Google Scholar] [CrossRef]
- Vaskinn, A.; Sundet, K.; Simonsen, C.; Hellvin, T.; Melle, I.; Andreassen, O.A. Sex Differences in Neuropsychological Performance and Social Functioning in Schizophrenia and Bipolar Disorder. Neuropsychology 2011, 25, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Huang, X.F.; Chen, D.C.; Xiu, M.H.; Hui, L.; Liu, H.; Kosten, T.R.; Zhang, X.Y. Gender differences in cognitive function of patients with chronic schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 39, 358–363. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Y.; Chen, D.C.; Xiu, M.H.; De Yang, F.; Haile, C.N.; Kosten, T.A.; Kosten, T.R. Gender differences in never-medicated first-episode schizophrenia and medicated chronic schizophrenia patients. J. Clin. Psychiatry 2012, 73, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Antonova, E.; Sharma, T.; Morris, R.; Kumari, V. The relationship between brain structure and neurocognition in schizophrenia: A selective review. Schizophr. Res. 2004, 70, 117–145. [Google Scholar] [CrossRef]
- Mendrek, A.; Mancini-Marïe, A. Sex/gender differences in the brain and cognition in schizophrenia. Neurosci. Biobehav. Rev. 2016, 67, 57–78. [Google Scholar] [CrossRef]
- Sweeney, J.A.; Maki, P.M. Effects of sex, menstrual cycle phase, and endogenous Hormones on Cognition in Schizophrenia. Schizophr. Res. 2016, 166, 269–275. [Google Scholar]
- Rasetti, R.; Weinberger, D.R. Intermediate phenotypes in psychiatric disorders. Curr. Opin. Genet. Dev. 2011, 21, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.A.; Scult, M.A.; Conley, E.D.; Chen, Q.; Weinberger, D.R.; Hariri, A.R. Effects of Schizophrenia Polygenic Risk Scores on Brain Activity and Performance during Working Memory Subprocesses in Healthy Young Adults. Schizophr. Bull. 2018, 44, 844–853. [Google Scholar] [CrossRef]
- Walton, E.; Turner, J.; Gollub, R.L.; Manoach, D.S.; Yendiki, A.; Ho, B.C.; Sponheim, S.R.; Calhoun, V.D.; Ehrlich, S. Cumulative genetic risk and prefrontal activity in patients with schizophrenia. Schizophr. Bull. 2013, 39, 703–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattay, V.S.; Marenco, S.; Egan, M.F.; Weinberger, D.R. Complexity of Prefrontl Cortical Dysfunction in Schizophrenia: More Than Up or Down. Am. J. Psychiatry 2003, 160, 2209–2215. [Google Scholar]
- Potkin, S.G.; Turner, J.A.; Brown, G.G.; Mccarthy, G.; Greve, D.N.; Glover, G.H.; Manoach, D.S.; Belger, A.; Diaz, M.; Wible, C.G.; et al. Working memory and DLPFC inefficiency in schizophrenia: The FBIRN study. Schizophr. Bull. 2009, 35, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Walton, E.; Geisler, D.; Lee, P.H.; Hass, J.; Turner, J.A.; Liu, J.; Sponheim, S.R.; White, T.; Wassink, T.H.; Roessner, V.; et al. Prefrontal Inefficiency Is Associated with Polygenic Risk for Schizophrenia. Schizophr. Bull. 2014, 40, 1263–1271. [Google Scholar] [CrossRef] [Green Version]
- Aleman, A.; Hijman, R.; De Haan, E.H.F.; Kahn, R.S. Memory impairment in schizophrenia: A meta-analysis. Am. J. Psychiatry 1999, 156, 1358–1366. [Google Scholar] [PubMed]
- Green, M. What are the functional consequences of neurocognitive deficits in schizophrenia? Am. J. Psychiatry 1996, 153, 321–330. [Google Scholar] [PubMed]
- Rushe, T.M.; Woodruff, P.W.R.; Murray, R.M.; Morris, R.G. Episodic memory and learning in patients with chronic schizophrenia. Schizophr. Res. 1999, 35, 85–96. [Google Scholar] [CrossRef]
- Heinrichs, R.; Zakzanis, K. Neurocognitive deficit in schizophrenia: A quantitative review of the evidence. Neuropsychology 1998, 12, 426–445. [Google Scholar] [CrossRef]
- Dezhina, Z.; Ranlund, S.; Kyriakopoulos, M.; Williams, S.C.R.; Dima, D. A systematic review of associations between functional MRI activity and polygenic risk for schizophrenia and bipolar disorder. Brain Imaging Behav. 2019, 13, 862–877. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, J.A.; Mancini-Marïe, A.; Lakis, N.; Rinaldi, M.; Mendrek, A. Disturbed sexual dimorphism of brain activation during mental rotation in schizophrenia. Schizophr. Res. 2010, 122, 53–62. [Google Scholar] [CrossRef]
- Nyberg, L.; Boraxbekk, C.J.; Sörman, D.E.; Hansson, P.; Herlitz, A.; Kauppi, K.; Ljungberg, J.K.; Lövheim, H.; Lundquist, A.; Adolfsson, A.N.; et al. Biological and environmental predictors of heterogeneity in neurocognitive ageing: Evidence from Betula and other longitudinal studies. Ageing Res. Rev. 2020, 64, 101184. [Google Scholar] [CrossRef]
- Nyberg, L.; Andersson, M.; Kauppi, K.; Lundquist, A.; Persson, J.; Pudas, S.; Nilsson, L.G. Age-related and Genetic Modulation of Frontal Cortex. J. Cogn. Neurosci. 2014, 26, 746–754. [Google Scholar] [CrossRef] [Green Version]
- Rieckmann, A.; Pudas, S.; Nyberg, L. Longitudinal Changes in Component Processes of Working Memory. Eneuro 2017, 4, ENEURO.0052-17.2017. [Google Scholar] [CrossRef] [Green Version]
- Salami, A.; Eriksson, J.; Nyberg, L. Opposing Effects of Aging on Large-Scale Brain Systems for Memory Encoding and Cognitive Control. J. Neurosci. 2012, 32, 10749–10757. [Google Scholar] [CrossRef] [Green Version]
- Pudas, S.; Josefsson, M.; Rieckmann, A.; Nyberg, L. Longitudinal Evidence for Increased Functional Response in Frontal Cortex for Older Adults with Hippocampal Atrophy and Memory Decline. Cereb. Cortex 2018, 28, 936–948. [Google Scholar] [CrossRef]
- Nyberg, L.; Andersson, M.; Lundquist, A.; Salami, A.; Wåhlin, A. Frontal Contribution to Hippocampal Hyperactivity During Memory Encoding in Aging. Front. Mol. Neurosci. 2019, 12, 229. [Google Scholar] [CrossRef]
- Ashburner, J. A fast diffeomorphic image registration algorithm. Neuroimage 2007, 38, 95–113. [Google Scholar] [CrossRef]
- The 1000 Genomes Project Consortium. A global reference for human genetic variation. Nature 2016, 526, 68–74. [Google Scholar]
- Howie, B.; Fuchsberger, C.; Stephens, M.; Marchini, J.; Abecasis, G.R. Fast and accurate genotype imputation in genome-wide association studies through pre-phasing. Nat. Genet. 2013, 44, 955–959. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Calafato, M.S.; Thgesen, J.H.; Ranlund, S.; Zartaloudi, E.; Cahn, W.; Crespo-Facorro, B.; Diéz-Revuelta, Á.; Di Forti, M.; Hall, M.H.; Iyegbe, C.; et al. Use of schizophrenia and bipolar disorder polygenic risk scores to identify psychotic disorders. Br. J. Psychiatry 2018, 213, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Coombes, B.J.; Ploner, A.; Bergen, S.E.; Biernacka, J.M. A principal component approach to improve association testing with polygenic risk scores. Genet. Epidemiol. 2020, 44, 676–686. [Google Scholar] [CrossRef]
- Lee, J.J.; Wedow, R.; Okbay, A.; Kong, E.; Maghzian, O.; Zacher, M.; Nguyen-Viet, T.A.; Bowers, P.; Sidorenko, J.; Karlsson Linnér, R.R.; et al. Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nat. Genet. 2018, 50, 1112–1121. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, J.; Vogel, E.K.; Lansner, A.; Bergström, F.; Nyberg, L. Neurocognitive Architecture of Working Memory. Neuron 2015, 88, 33–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manoach, D.S.; Press, D.Z.; Thangaraj, V.; Searl, M.M.; Goff, D.C.; Halpern, E.; Saper, C.B.; Warach, S. Schizophrenic Subjects Activate Dorsolateral Prefrontal Cortex during a Working Memory Task, as Measured by fMRI. Biol. Psychiatry 1999, 45, 1128–1137. [Google Scholar] [CrossRef]
- Van Veelen, N.M.J.; Vink, M.; Ramsey, N.F.; Kahn, R.S. Left dorsolateral prefrontal cortex dysfunction in medication-naive schizophrenia. Schizophr. Res. 2010, 123, 22–29. [Google Scholar] [CrossRef]
- Guimond, S.; Hawco, C.; Lepage, M. Prefrontal activity and impaired memory encoding strategies in schizophrenia. J. Psychiatr. Res. 2017, 91, 64–73. [Google Scholar] [CrossRef]
- Lepage, M.; Pelletier, M.; Achim, A.; Montoya, A.; Menear, M.; Lal, S. Parietal cortex and episodic memory retrieval in schizophrenia. Psychiatry Res. Neuroimaging 2010, 182, 191–199. [Google Scholar] [CrossRef]
- Uncapher, M.; Wagner, A.D. Posterior Parietal Cortex and Episodic Encoding: Insights from fMRI Subsequent Memory Effects and Dual Attention Theory. Neurobiol. Learn. Mem. 2010, 91, 139–154. [Google Scholar] [CrossRef] [Green Version]
- Wagner, A.D.; Shannon, B.J.; Kahn, I.; Buckner, R.L. Parietal lobe contributions to episodic memory retrieval. Trends Cogn. Sci. 2005, 9, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, J.B.; Uncapher, M.R.; Wagner, A.D. Posterior parietal cortex and episodic retrieval: Convergent and divergent effects of attention and memory. Learn. Mem. 2009, 16, 343–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konishi, S.; Wheeler, M.E.; Donaldson, D.I.; Buckner, R.L. Neural Correlates of Episodic Retrieval Success. Neuroimage 2000, 286, 276–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbetta, M.; Shulman, G.L. Control of goal-directed and stimulus-driven attention in the brain. Nat. Rev. Neurosci. 2002, 3, 201–215. [Google Scholar] [CrossRef]
- Cabeza, R.; Ciaramelli, E.; Olson, I.R.; Moscovitch, M. The parietal cortex and episodic memory: An attentional account. Nat. Rev. Neurosci. 2009, 9, 613–625. [Google Scholar] [CrossRef] [Green Version]
- Peters, E.R.; Nunn, J.A.; Pickering, A.D.; Hemsley, D.R. Perceptual organization deficits in psychotic patients. Psychiatry Res. 2002, 110, 125–135. [Google Scholar] [CrossRef]
- Gold, J.M.; Fuller, R.L.; Robinson, B.M.; Braun, E.L.; Luck, S.J. Impaired top-down control of visual search in schizophrenia. Schizophr. Res. 2007, 94, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Fuller, R.L.; Luck, S.J.; Braun, E.L.; Robinson, B.M.; McMahon, R.P.; Gold, J.M. Impaired control of visual attention in schizophrenia. J. Abnorm. Psychol. 2006, 115, 266–275. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Baseline | Follow-Up | t/χ2 Value | p-Value | |
|---|---|---|---|---|
| Working Memory | ||||
| Age, Mean (SD) | 63.13 (13.46) | 64.94 (13.01) | −1.57 | 0.12 a |
| Sex, N females (males) | 181 (163) | 101 (111) | 1.11 | 0.29 b |
| RT, Maintenance | 1.35 (0.27) | 1.30 (0.26) | 2.26 | 0.02 a |
| RT, Manipulation | 1.45 (0.32) | 1.40 (0.32) | 1.57 | 0.12 a |
| Hits - FA, Maintenance | 7.93 (1.58) | 8.15 (1.33) | −1.73 | 0.08 a |
| Hits - FA, Manipulation | 6.42 (2.50) | 6.47 (2.36) | −0.25 | 0.80 a |
| Episodic Memory | ||||
| Age, Mean (SD) | 63.10 (13.25) | 65.01 (12.97) | −1.69 | 0.09 a |
| Sex, N females (males) | 184 (167) | 103 (113) | 1.02 | 0.31 b |
| RT, Retrieval, Mean (SD) | 2.62 (0.34) | 2.65 (0.44) | −0.89 | 0.37a |
| Hits, Retrieval, Mean (SD) | 14.29 (4.42) | 16.10 (4.00) | −5.00 | 8.02 × 10−7 a |
| Adjusted R2 | SE | t-Value (df) | p-Value | ||
|---|---|---|---|---|---|
| Working Memory | |||||
| RT, Maintenance | All | −0.0022 | 0.0135 | 0.248 (343) | 0.804 |
| Males | 0.0070 | 0.0222 | 1.524 (162) | 0.129 | |
| Females | 0.0019 | 0.0165 | −1.215 (180) | 0.226 | |
| RT, Manipulation | All | −0.0026 | 0.0176 | 0.111 (343) | 0.912 |
| Males | 0.0023 | 0.0284 | 1.174 (162) | 0.242 | |
| Females | −0.0009 | 0.0220 | −0.897 (180) | 0.371 | |
| Hits - FA, Maintenance | All | 0.0438 | 0.0866 | −1.088 (343) | 0.277 |
| Males | 0.0257 | 0.1361 | −2.357 (162) | 0.020 * | |
| Females | −0.0013 | 0.1116 | 0.866 (180) | 0.388 | |
| Hits - FA, Manipulation | All | 0.1964 | 0.1209 | −2.537 (343) | 0.012 * |
| Males | 0.0240 | 0.1746 | −2.489 (162) | 0.014* | |
| Females | 0.0020 | 0.1688 | −1.219 (180) | 0.224 | |
| Episodic Memory | |||||
| RT, Retrieval | All | 0.0031 | 0.0172 | 1.531 (350) | 0.127 |
| Males | 0.0186 | 0.0256 | 2.203 (166) | 0.029 * | |
| Females | −0.0040 | 0.0228 | 0.190 (183) | 0.849 | |
| Hits, Retrieval | All | 0.0034 | 0.2097 | −1.660 (350) | 0.098 |
| Males | 0.0319 | 0.3064 | −2.904 (166) | 0.004 * | |
| Females | −0.0034 | 0.2830 | 0.045 (183) | 0.964 | |
| Coordinates | Brain Region | t-Value (df) | p-Value | N | Follow-Up, Coordinates, p (N) | |
|---|---|---|---|---|---|---|
| Working Memory | ||||||
| Manip-Ctrl | ||||||
| Neg cor PRS | 4 −96 4 | No area | 3.87 (343) | 0.000064 | 31 | ns |
| −6 −6 14 | L Thalamus | 3.64 (343) | 0.000156 | 41 | ns | |
| 6 −62 −36 | CVL | 3.55 (343) | 0.000219 | 45 | ns | |
| 40 −86 −2 | R IOG | 3.50 (343) | 0.000268 | 17 | ns | |
| −46 −44 22 | L cerebral WM | 3.45 (343) | 0.000315 | 17 | ns | |
| Manip-Maint | ||||||
| Neg cor PRS | −44 40 30 | L DLPFC | 4.20 (343) | 0.000017 | 80 | ns |
| −52 22 −6 | No area | 3.66 (343) | 0.000145 | 60 | ns | |
| −36 −76 40 | L angular gyrus | 3.23 (343) | 0.000669 | 14 | ns | |
| −2 −26 −8 | No area | 3.21 (343) | 0.000737 | 10 | ns | |
| Episodic memory | ||||||
| Enc-Ctrl | ||||||
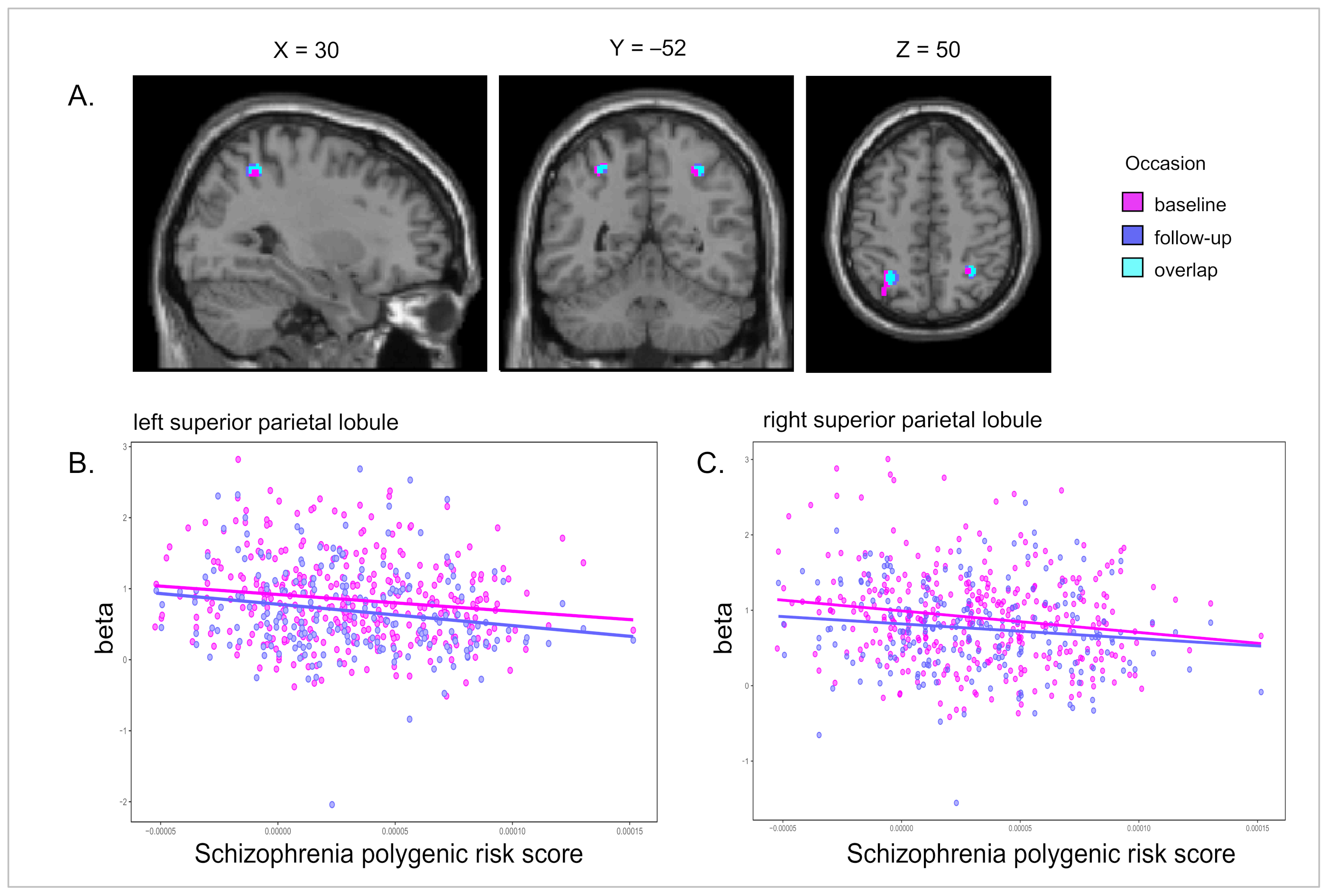
| Neg cor PRS | −30 −52 56 | L SPL | 4.04 (350) | 0.000034 | 137 | −28 −54 52, 0.004600 (44) |
| 34 −78 32 | No area | 3.36 (350) | 0.000430 | 19 | ns | |
| 30 −48 52 | R SPL | 3.35 (350) | 0.000454 | 28 | 34 −46 52, 0.010790 (26) | |
| Retr-Ctrl | ||||||
| Neg cor PRS | 54 −72 6 | R IOG | 3.80 (350) | 0.000085 | 47 | ns |
| −28 −56 52 | L SPL | 3.62 (350) | 0.000171 | 95 | −26 −54 56, 0.004511 (80) | |
| 0 −96 16 | No area | 3.59 (350) | 0.000191 | 59 | ns | |
| 30 −52 50 | R SPL | 3.50 (350) | 0.000268 | 32 | 34 −50 52, 0.013565 (45) | |
| 20 −78 52 | No area | 3.35 (350) | 0.000449 | 17 | ns | |
| −20 −82 42 | No area | 3.29 (350) | 0.000551 | 30 | ns | |
| Pos cor PRS | 34 −12 −28 | R HC | 3.41 (350) | 0.000361 | 14 | ns |
| Sex | Coordinates | Brain Region | t-Value (df) | p-Value | N Voxels with p < 0.001 | |
|---|---|---|---|---|---|---|
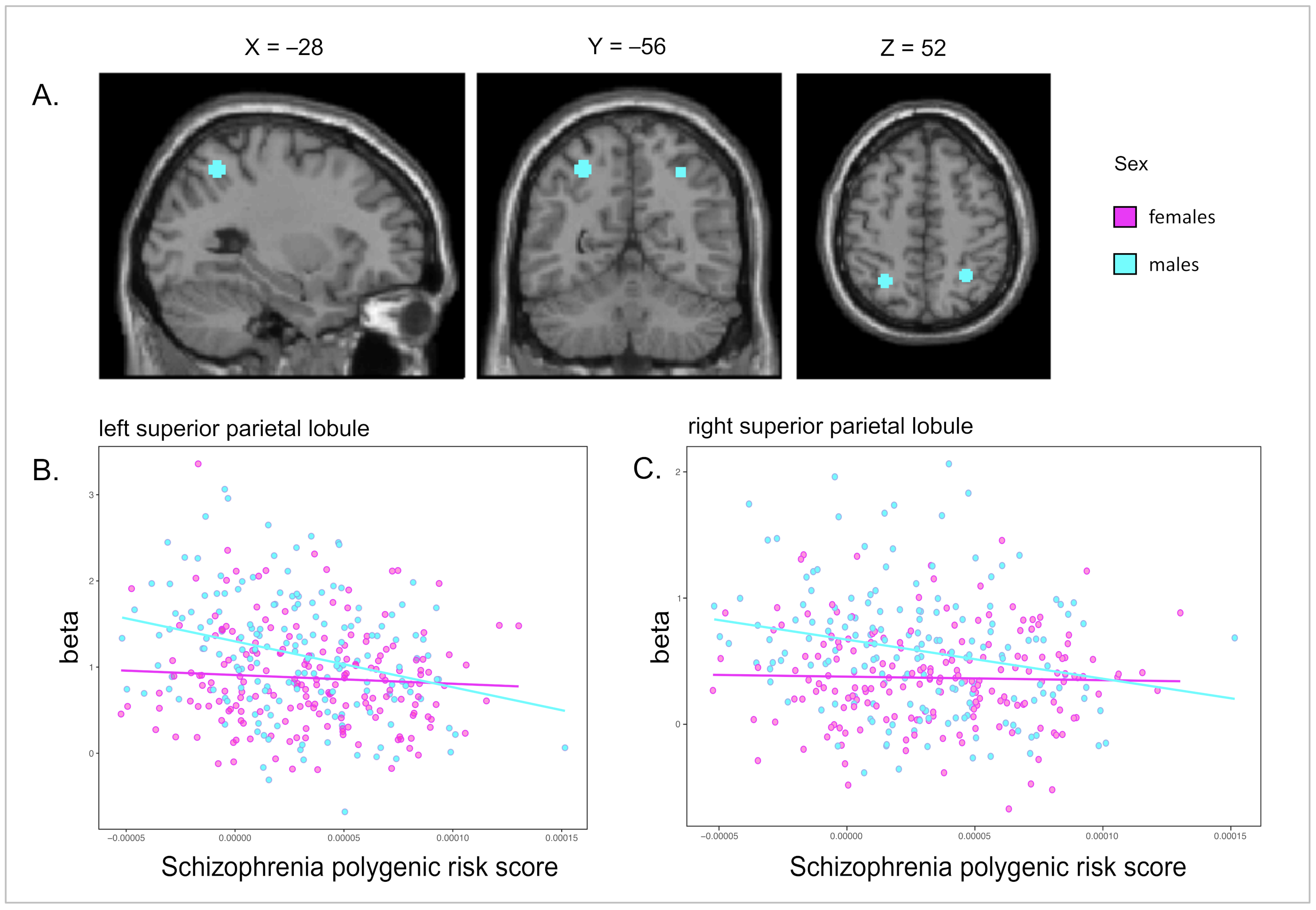
| Enc-Ctrl | Males | −30 −54 56 | L SPL | 3.99 (166) | 0.00005 * | 81 of 81 |
| 34 −50 54 | R SPL | 3.36 (166) | 0.00048 * | 75 of 81 | ||
| Females | −26 −54 56 | L SPL | 2.09 (183) | 0.01893 | 0 of 81 | |
| 30 −52 50 | R SPL | 2.08 (183) | 0.01927 | 0 of 81 | ||
| Retr-Ctrl | Males | −28 −56 52 | L SPL | 3.92 (166) | 0.00007 * | 81 of 81 |
| 32 −50 54 | R SPL | 3.39 (166) | 0.00044 * | 81 of 81 | ||
| Females | −30 −60 50 | L SPL | 1.53 (183) | 0.06169 | 0 of 81 | |
| 30 −54 50 | R SPL | 1.96 (183) | 0.02547 | 0 of 81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koch, E.; Nyberg, L.; Lundquist, A.; Kauppi, K. Polygenic Risk for Schizophrenia Has Sex-Specific Effects on Brain Activity during Memory Processing in Healthy Individuals. Genes 2022, 13, 412. https://doi.org/10.3390/genes13030412
Koch E, Nyberg L, Lundquist A, Kauppi K. Polygenic Risk for Schizophrenia Has Sex-Specific Effects on Brain Activity during Memory Processing in Healthy Individuals. Genes. 2022; 13(3):412. https://doi.org/10.3390/genes13030412
Chicago/Turabian StyleKoch, Elise, Lars Nyberg, Anders Lundquist, and Karolina Kauppi. 2022. "Polygenic Risk for Schizophrenia Has Sex-Specific Effects on Brain Activity during Memory Processing in Healthy Individuals" Genes 13, no. 3: 412. https://doi.org/10.3390/genes13030412
APA StyleKoch, E., Nyberg, L., Lundquist, A., & Kauppi, K. (2022). Polygenic Risk for Schizophrenia Has Sex-Specific Effects on Brain Activity during Memory Processing in Healthy Individuals. Genes, 13(3), 412. https://doi.org/10.3390/genes13030412