
Genomic Predictions for Common Bunt, FHB, Stripe Rust, Leaf Rust, and Leaf Spotting Resistance in Spring Wheat

 ,
, 
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Germplasm and Phenotyping
2.2. DNA Extraction and Genotyping
2.3. Data Analyses
3. Results
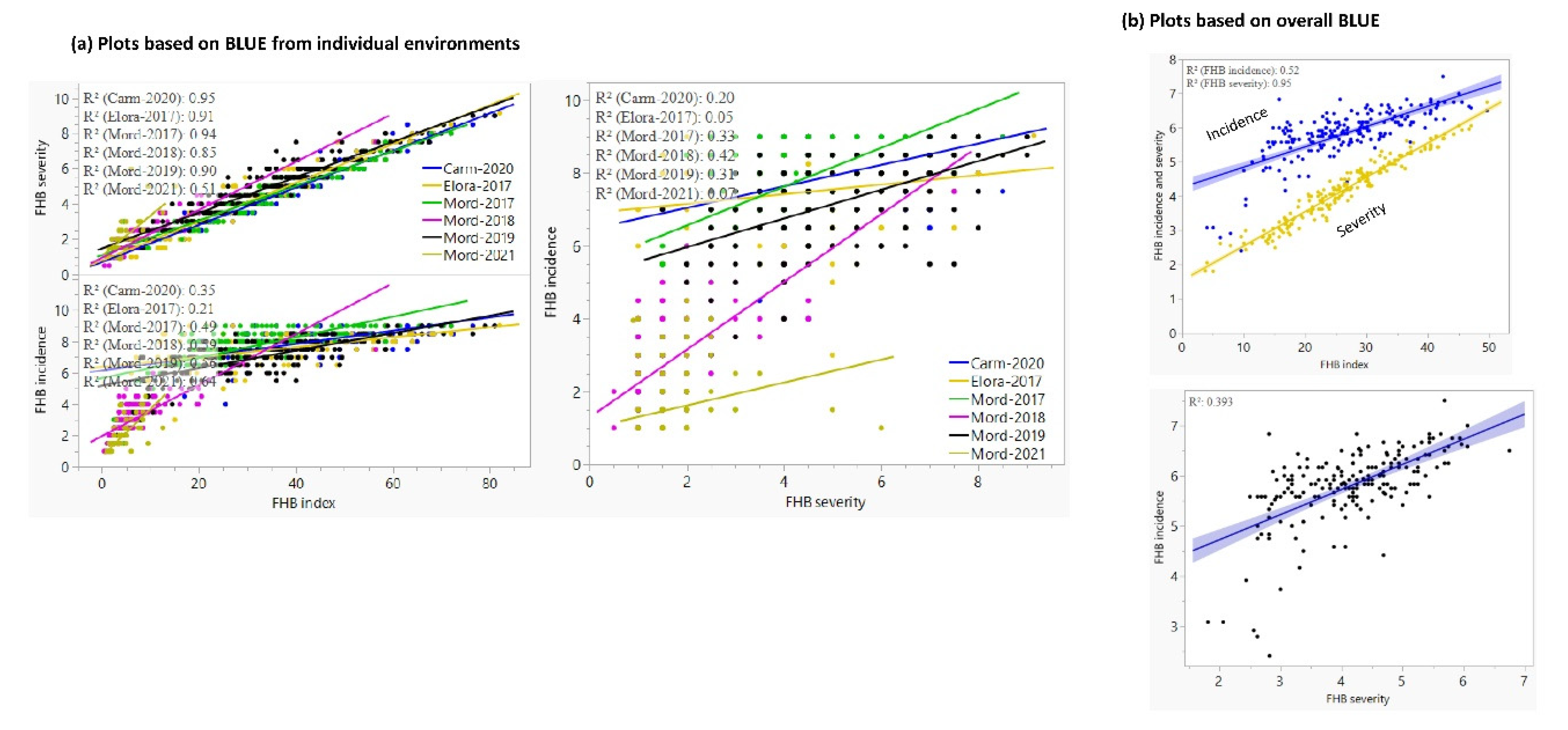
3.1. Reaction to Diseases
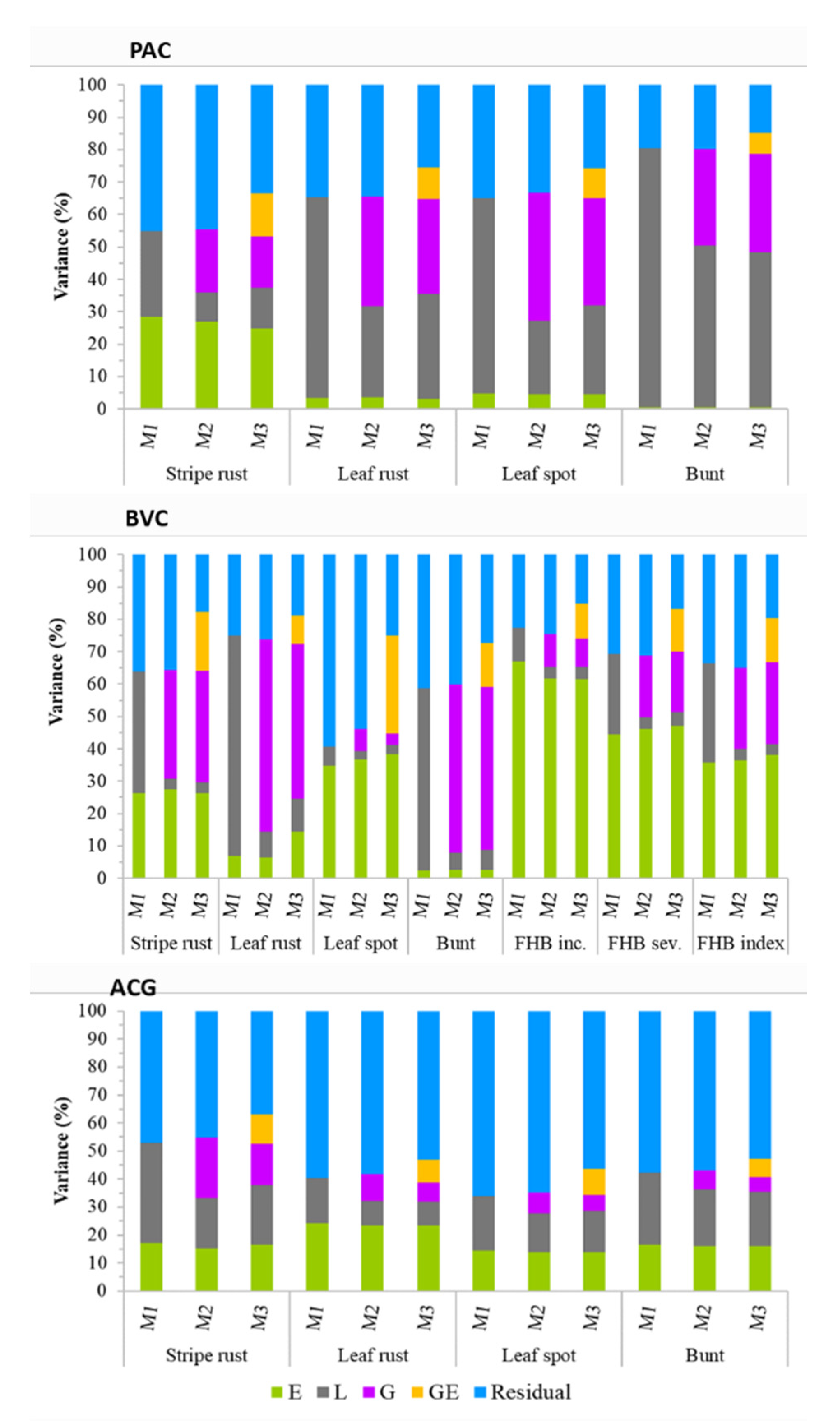
3.2. Partitioning of Variance Components
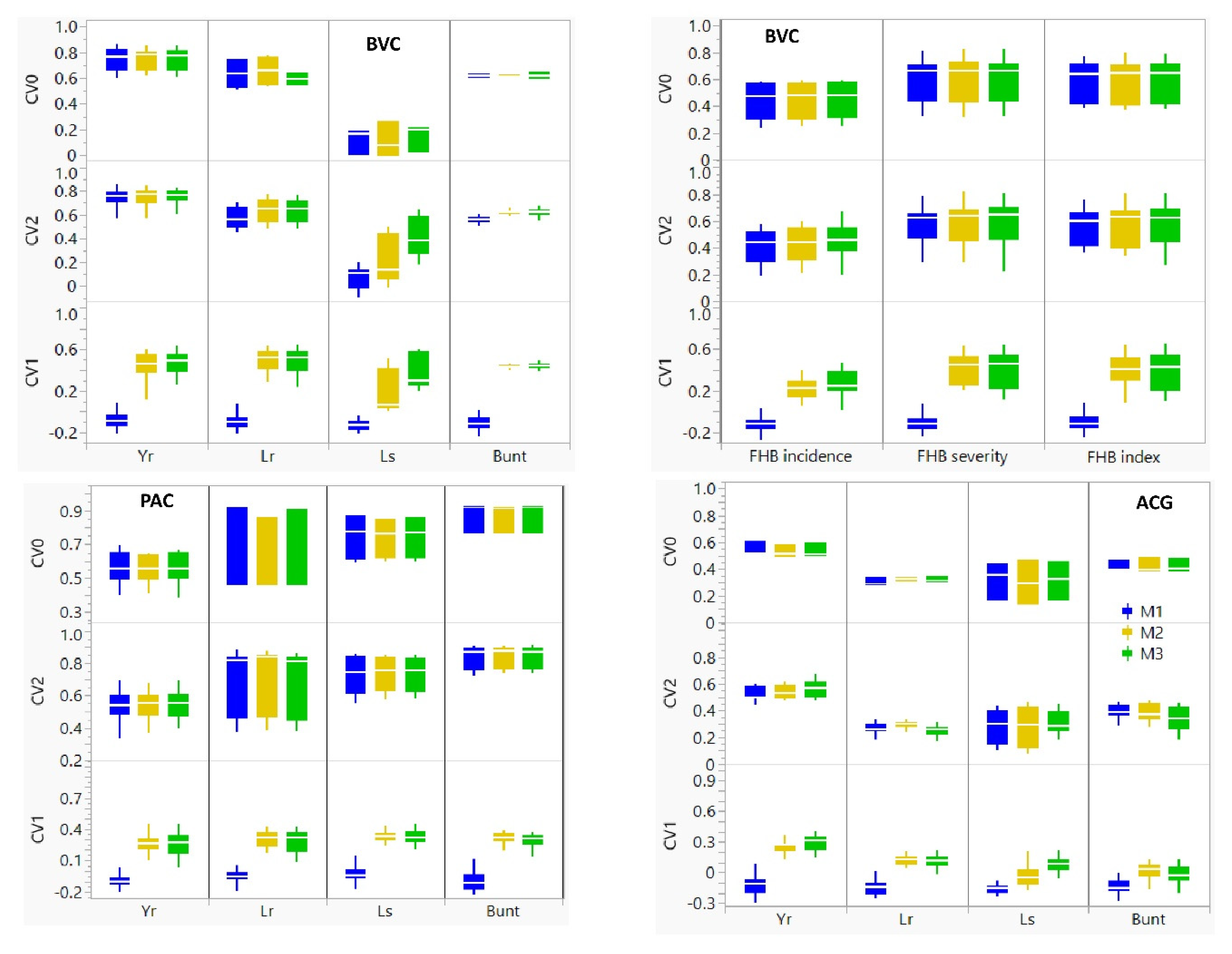
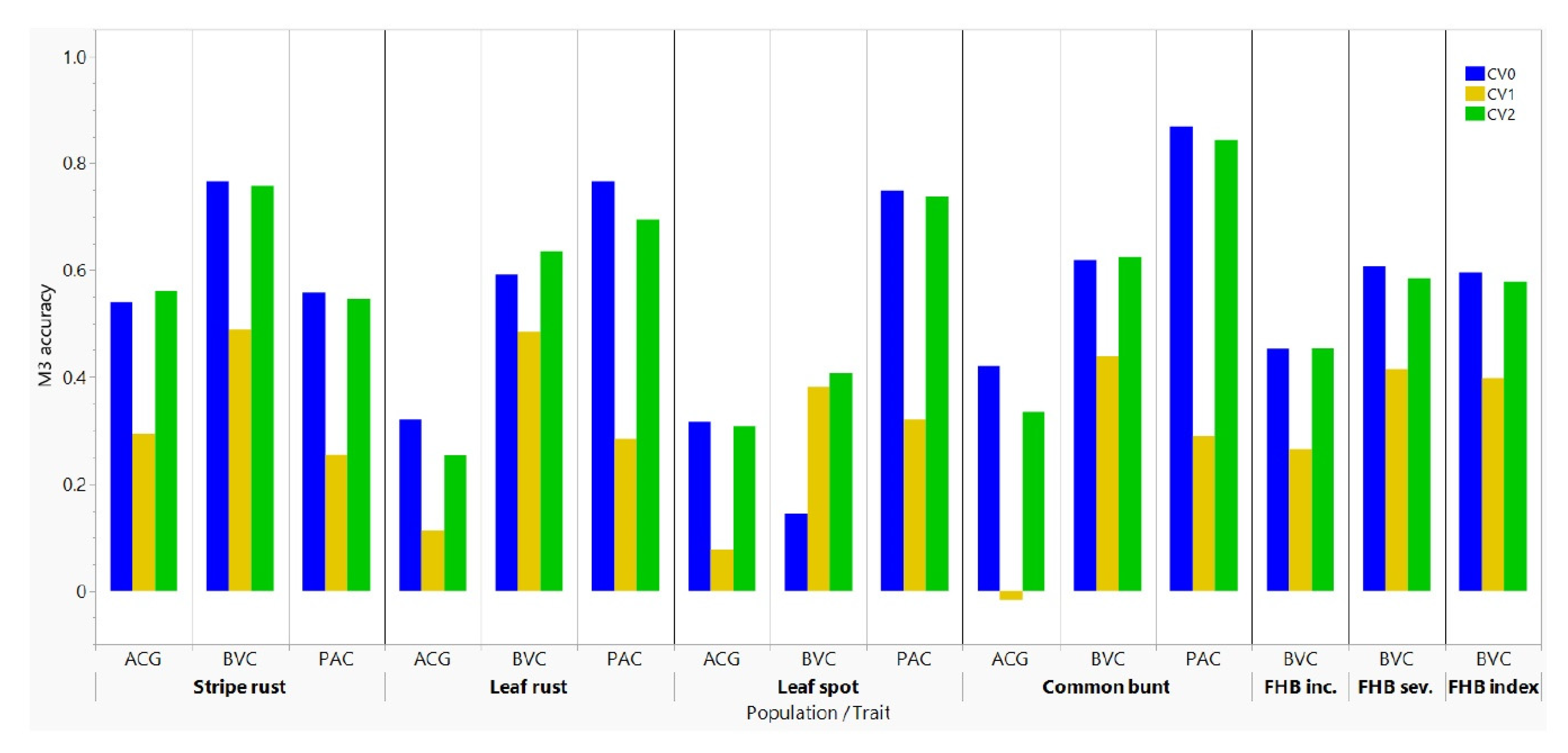
3.3. Comparison of Prediction Models
4. Discussion
4.1. Comparisons of Prediction Models
4.2. Prediction Accuracies among Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brar, G.S.; Fetch, T.; McCallum, B.D.; Hucl, P.J.; Kutcher, H.R. Virulence dynamics and breeding for resistance to stripe, stem, and leaf rust in Canada since 2000. Plant Dis. 2019, 103, 2981–2995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svarta, A.; Bimsteine, G. Winter wheat leaf diseases and several steps included in their integrated control: A review. In Proceedings of the Research for Rural Development, Jelgava, Latvia, 15–17 May 2019; pp. 55–62. [Google Scholar]
- Figueroa, M.; Hammond-Kosack, K.E.; Solomon, P.S. A review of wheat diseases—a field perspective. Mol. Plant Pathol. 2018, 19, 1523–1536. [Google Scholar] [CrossRef] [PubMed]
- Aboukhaddour, R.; Fetch, T.; McCallum, B.D.; Harding, M.W.; Beres, B.L.; Graf, R.J. Wheat diseases on the prairies: A Canadian story. Plant Pathol. 2020, 69, 418–432. [Google Scholar] [CrossRef]
- McCallum, B.D.; Reimer, E.; McNabb, W.; Foster, A.; Rosa, S.; Xue, A. Physiologic specialization of Puccinia triticina, the causal agent of wheat leaf rust, in Canada in 2015–2019. Can. J. Plant Pathol. 2021, 33, 541–549. [Google Scholar] [CrossRef]
- McCallum, B.; Pearse, P.; Fetch, T., Jr.; Gaudet, D.; Tekauz, A.; Seto-Goh, P.; Xi, K.; Turkington, T. Stripe rust of wheat and barley in Manitoba, Saskatchewan and Alberta in 2006. Can. Plant Disease Surv. 2006, 86, 50–51. [Google Scholar]
- Peturson, B. Wheat rust epidemics in western Canada in 1953, 1954 and 1955. Can. J. Plant Sci. 1958, 38, 16–28. [Google Scholar] [CrossRef]
- Wegulo, S.; Byamukama, E. Rust Diseases of Wheat; University of Nebraska–Lincoln: Lincoln, NE, USA, 2012. [Google Scholar]
- Gaudet, D.A.; Puchalski, B.L. Races of common bunt (Tilletia caries and T. foetida) of wheat in western Canada. Can. J. Plant Pathol. 1989, 11, 415–418. [Google Scholar] [CrossRef]
- Martens, J.W.; Seaman, W.L.; Atkinson, T.G. Diseases of Field Crops in Canada. An Illustrated Compendium; Canadian Phytopathological Society: Harrow, ON, Canada, 1984; p. 8 + 159. [Google Scholar]
- Tekauz, A.; McCallum, B.; Gilbert, J. Review: Fusarium head blight of barley in western Canada. Can. J. Plant Pathol. 2000, 22, 9–16. [Google Scholar] [CrossRef]
- Dhariwal, R.; Henriquez, M.A.; Hiebert, C.; McCartney, C.A.; Randhawa, H.S. Mapping of major Fusarium head blight resistance from Canadian wheat cv. AAC Tenacious. Int. J. Mol. Sci. 2020, 21, 4497. [Google Scholar] [CrossRef]
- Bai, G.; Shaner, G. Scab of wheat: Prospects for control. Plant Dis. 1994, 78, 760–766. [Google Scholar] [CrossRef]
- Gilbert, J.; Tekauz, A. Recent developments in research on fusarium head blight of wheat in Canada. Can. J. Plant Pathol. 2000, 22, 1–8. [Google Scholar] [CrossRef]
- Rod, K.S.; Bradley, C.A.; Van Sanford, D.A.; Knott, C.A. Integrating Management Practices to Decrease Deoxynivalenol Contamination in Soft Red Winter Wheat. Front. Plant Sci. 2020, 11, 1158. [Google Scholar] [CrossRef] [PubMed]
- Schaafsma, A.W.; Tamburic-Ilincic, L. Effect of Seeding Rate and Seed Treatment Fungicides on Agronomic Performance, Fusarium Head Blight Symptoms, and DON Accumulation in Two Winter Wheats. Plant Dis. 2005, 89, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Shah, L.; Ali, A.; Yahya, M.; Zhu, Y.; Wang, S.; Si, H.; Rahman, H.; Ma, C. Integrated control of fusarium head blight and deoxynivalenol mycotoxin in wheat. Plant Pathol. 2018, 67, 532–548. [Google Scholar] [CrossRef]
- Lu, Q.; Szabo-Hever, A.; Bjørnstad, Å.; Lillemo, M.; Semagn, K.; Mesterhazy, A.; Ji, F.; Shi, J.; Skinnes, H. Two major resistance quantitative trait loci are required to counteract the increased susceptibility to Fusarium head blight of the Rht-D1b dwarfing gene in wheat. Crop Sci. 2011, 51, 2430–2438. [Google Scholar] [CrossRef]
- Gosman, N.; Steed, A.; Simmonds, J.; Leverington-Waite, M.; Wang, Y.; Snape, J.; Nicholson, P. Susceptibility to Fusarium head blight is associated with the Rht-D1b semi-dwarfing allele in wheat. Theor. Appl. Genet. 2008, 116, 1145–1153. [Google Scholar] [CrossRef]
- He, X.; Singh, P.K.; Dreisigacker, S.; Singh, S.; Lillemo, M.; Duveiller, E. Dwarfing genes Rht-B1b and Rht-D1b are associated with both type I FHB susceptibility and low anther extrusion in two bread wheat populations. PLoS ONE 2016, 11, e0162499. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, M.R.; Fox, S.L.; Hucl, P.; Singh, A.K. Leaf spotting reaction of spring common, durum and spelt wheat, and Kamut under organic management in western Canada. Can. J. Plant Sci. 2014, 94, 929–935. [Google Scholar] [CrossRef]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef]
- Heffner, E.L.; Lorenz, A.J.; Jannink, J.L.; Sorrells, M.E. Plant breeding with genomic selection: Gain per unit time and cost. Crop Sci. 2010, 50, 1681–1690. [Google Scholar] [CrossRef]
- Bassi, F.M.; Bentley, A.R.; Charmet, G.; Ortiz, R.; Crossa, J. Breeding schemes for the implementation of genomic selection in wheat (Triticum spp.). Plant Sci. 2016, 242, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Cobb, J.N.; Juma, R.U.; Biswas, P.S.; Arbelaez, J.D.; Rutkoski, J.; Atlin, G.; Hagen, T.; Quinn, M.; Ng, E.H. Enhancing the rate of genetic gain in public-sector plant breeding programs: Lessons from the breeder’s equation. Theor. Appl. Genet. 2019, 132, 627–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, H.; Wang, R.; Yuan, Y.; Anderson, J.; Pumphrey, M.; Zhang, Z.; Chen, J. Evaluation of the potential for genomic selection to improve spring wheat resistance to fusarium head blight in the pacific northwest. Front. Plant Sci. 2018, 9, 911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muleta, K.T.; Bulli, P.; Zhang, Z.; Chen, X.; Pumphrey, M. Unlocking diversity in germplasm collections via genomic selection: A case study based on quantitative adult plant resistance to stripe rust in spring wheat. Plant Genome 2017, 10, plantgenome2016.2012.0124. [Google Scholar] [CrossRef] [Green Version]
- Tehseen, M.M.; Kehel, Z.; Sansaloni, C.P.; Lopes, M.D.S.; Amri, A.; Kurtulus, E.; Nazari, K. Comparison of genomic prediction methods for yellow, stem, and leaf rust resistance in wheat landraces from afghanistan. Plants 2021, 10, 558. [Google Scholar] [CrossRef]
- Ornella, L.; Sukhwinder, S.; Perez, P.; Burgueno, J.; Singh, R.; Tapia, E.; Bhavani, S.; Dreisigacker, S.; Braun, H.J.; Mathews, K.; et al. Genomic prediction of genetic values for resistance to wheat rusts. Plant Genome 2012, 5, 136–148. [Google Scholar] [CrossRef] [Green Version]
- Juliana, P.; Singh, R.P.; Singh, P.K.; Crossa, J.; Rutkoski, J.E.; Poland, J.A.; Bergstrom, G.C.; Sorrells, M.E. Comparison of models and whole-genome profiling approaches for genomic-enabled prediction of Septoria tritici blotch, Stagonospora nodorum blotch, and tan spot resistance in wheat. Plant Genome 2017, 10, plantgenome2016.2008.0082. [Google Scholar] [CrossRef] [Green Version]
- Juliana, P.; Singh, R.P.; Singh, P.K.; Crossa, J.; Huerta-Espino, J.; Lan, C.; Bhavani, S.; Rutkoski, J.E.; Poland, J.A.; Bergstrom, G.C.; et al. Genomic and pedigree-based prediction for leaf, stem, and stripe rust resistance in wheat. Theor. Appl. Genet. 2017, 130, 1415–1430. [Google Scholar] [CrossRef] [Green Version]
- Arruda, M.P.; Brown, P.J.; Lipka, A.E.; Krill, A.M.; Thurber, C.; Kolb, F.L. Genomic selection for predicting Fusarium head blight resistance in a wheat breeding program. Plant Genome 2015, 8, plantgenome2015.2001.0003. [Google Scholar] [CrossRef] [Green Version]
- Arruda, M.P.; Brown, P.; Brown-Guedira, G.; Krill, A.M.; Thurber, C.; Merrill, K.R.; Foresman, B.J.; Kolb, F.L. Genome-wide association mapping of Fusarium head blight resistance in wheat using genotyping-by-sequencing. Plant Genome 2016, 9, plantgenome2015.2004.0028. [Google Scholar] [CrossRef]
- Azizinia, S.; Bariana, H.; Kolmer, J.; Pasam, R.; Bhavani, S.; Chhetri, M.; Toor, A.; Miah, H.; Hayden, M.J.; Del Carpio, D.P.; et al. Genomic prediction of rust resistance in tetraploid wheat under field and controlled environment conditions. Agronomy 2020, 10, 1843. [Google Scholar] [CrossRef]
- Beukert, U.; Liu, G.; Thorwarth, P.; Boeven, P.H.G.; Longin, C.F.H.; Zhao, Y.; Ganal, M.; Serfling, A.; Ordon, F.; Reif, J.C. The potential of hybrid breeding to enhance leaf rust and stripe rust resistance in wheat. Theor. Appl. Genet. 2020, 133, 2171–2181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beukert, U.; Thorwarth, P.; Zhao, Y.; Longin, C.F.H.; Serfling, A.; Ordon, F.; Reif, J.C. Comparing the potential of marker-assisted selection and genomic prediction for improving rust resistance in hybrid wheat. Front. Plant Sci. 2020, 11, 594113. [Google Scholar] [CrossRef] [PubMed]
- Daetwyler, H.D.; Bansal, U.K.; Bariana, H.S.; Hayden, M.J.; Hayes, B.J. Genomic prediction for rust resistance in diverse wheat landraces. Theor. Appl. Genet. 2014, 127, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Merrick, L.F.; Burke, A.B.; Chen, X.; Carter, A.H. Breeding with major and minor genes: Genomic selection for quantitative disease resistance. Front. Plant Sci. 2021, 12, 1599. [Google Scholar] [CrossRef] [PubMed]
- Alemu, A.; Brazauskas, G.; Gaikpa, D.S.; Henriksson, T.; Islamov, B.; Jørgensen, L.N.; Koppel, M.; Koppel, R.; Liatukas, Ž.; Svensson, J.T.; et al. Genome-wide association analysis and genomic prediction for adult-plant resistance to Septoria tritici blotch and powdery mildew in winter wheat. Front. Genet. 2021, 12, 661742. [Google Scholar] [CrossRef]
- Odilbekov, F.; Armoniené, R.; Koc, A.; Svensson, J.; Chawade, A. GWAS-assisted genomic prediction to predict resistance to Septoria tritici blotch in Nordic winter wheat at seedling stage. Front. Genet. 2019, 10, 1224. [Google Scholar] [CrossRef]
- Muqaddasi, Q.H.; Kamal, R.; Mirdita, V.; Rodemann, B.; Ganal, M.W.; Reif, J.C.; Röder, M.S. Genome-wide association studies and prediction of tan spot (Pyrenophora tritici-repentis) infection in european winter wheat via different marker platforms. Genes 2021, 12, 490. [Google Scholar] [CrossRef]
- Herter, C.P.; Ebmeyer, E.; Kollers, S.; Korzun, V.; Miedaner, T. An experimental approach for estimating the genomic selection advantage for Fusarium head blight and Septoria tritici blotch in winter wheat. Theor. Appl. Genet. 2019, 132, 2425–2437. [Google Scholar] [CrossRef]
- Herter, C.P.; Ebmeyer, E.; Kollers, S.; Korzun, V.; Würschum, T.; Miedaner, T. Accuracy of within- and among-family genomic prediction for Fusarium head blight and Septoria tritici blotch in winter wheat. Theor. Appl. Genet. 2019, 132, 1121–1135. [Google Scholar] [CrossRef]
- Mirdita, V.; He, S.; Zhao, Y.; Korzun, V.; Bothe, R.; Ebmeyer, E.; Reif, J.C.; Jiang, Y. Potential and limits of whole genome prediction of resistance to Fusarium head blight and Septoria tritici blotch in a vast Central European elite winter wheat population. Theor. Appl. Genet. 2015, 128, 2471–2481. [Google Scholar] [CrossRef] [PubMed]
- Jarquin, D.; Crossa, J.; Lacaze, X.; Cheyron, P.; Daucourt, J.l.; Lorgeou, J.; Piraux, F.C.; Guerreiro, L.; Perez, P.; Calus, M.; et al. A reaction norm model for genomic selection using high-dimensional genomic and environmental data. Theor. Appl. Genet. 2014, 127, 595–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgueno, J.; de los Campos, G.; Weigel, K.; Crossa, J. Genomic prediction of breeding values when modeling genotype × environment interaction using pedigree and dense molecular markers. Crop Sci. 2012, 52, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Crossa, J.; de los Campos, G.; Maccaferri, M.; Tuberosa, R.; Burgueño, J.; Pérez-Rodríguez, P. Extending the marker × environment interaction model for genomic-enabled prediction and genome-wide association analysis in durum wheat. Crop Sci. 2016, 56, 2193–2209. [Google Scholar] [CrossRef] [Green Version]
- Cuevas, J.; Crossa, J.; Montesinos-López, O.A.; Burgueño, J.; Pérez-Rodríguez, P.; de los Campos, G. Bayesian genomic prediction with genotype × environment interaction kernel models. Genes Genomes Genetics 2017, 7, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Cuevas, J.; Crossa, J.; Soberanis, V.; Pérez-Elizalde, S.; Pérez-Rodríguez, P.; Campos, G.d.l.; Montesinos-López, O.A.; Burgueño, J. Genomic prediction of genotype × environment interaction kernel regression models. Plant Genome 2016, 9, plantgenome2016.2003.0024. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Cruz, M.; Crossa, J.; Bonnett, D.; Dreisigacker, S.; Poland, J.; Jannink, J.L.; Singh, R.P.; Autrique, E.; de los Campos, G. Increased prediction accuracy in wheat breeding trials using a marker × environment interaction genomic selection model. Genes Genomes Genetics 2015, 5, 569–582. [Google Scholar] [CrossRef] [Green Version]
- Semagn, K.; Iqbal, M.; Crossa, J.; Jarquin, D.; Howard, R.; Chen, H.; Bemister, D.H.; Beres, B.L.; Randhawa, H.; N’Diaye, A.; et al. Genome-based prediction of agronomic traits in spring wheat under conventional and organic management systems. Theor. Appl. Genet. 2022, 135, 537–552. [Google Scholar] [CrossRef]
- Semagn, K.; Iqbal, M.; Alachiotis, N.; N’Diaye, A.; Pozniak, C.; Spaner, D. Genetic diversity and selective sweeps in historical and modern Canadian spring wheat cultivars using the 90K SNP array. Sci. Rep. 2021, 11, 23773. [Google Scholar] [CrossRef]
- Bokore, F.E.; Knox, R.E.; Cuthbert, R.D.; Pozniak, C.J.; McCallum, B.D.; N’Diaye, A.; DePauw, R.M.; Campbell, H.L.; Munro, C.; Singh, A.; et al. Mapping quantitative trait loci associated with leaf rust resistance in five spring wheat populations using single nucleotide polymorphism markers. PLoS ONE 2020, 15, e0230855. [Google Scholar] [CrossRef] [Green Version]
- Friesen, T.L.; Rasmussen, J.B.; Kwon, C.Y.; Ali, S.; Francl, L.J.; Meinhardt, S.W. Reaction of Ptr ToxA-insensitive wheat mutants to Pyrenophora tritici-repentis race 1. Phytopathology 2002, 92, 38–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aboukhaddour, R.; Turkington, T.K.; Strelkov, S.E. Race structure of Pyrenophora triciti-repentis (tan spot of wheat) in Alberta, Canada. Can. J. Plant Pathol. 2013, 35, 256–268. [Google Scholar] [CrossRef]
- Singh, P.K.; Mergoum, M.; Ali, S.; Adhikari, T.B.; Elias, E.M.; Anderson, J.A.; Glover, K.D.; Berzonsky, W.A. Evaluation of elite wheat germ plasm for resistance to tan spot. Plant Dis. 2006, 90, 1320–1325. [Google Scholar] [CrossRef] [Green Version]
- Peterson, R.F.; Campbell, A.B.; Hannah, A.E. A diagrammatic scale for estimating rust intensity on leaves and stems of cereals. Can. J. Res. 1948, 26, 496–500. [Google Scholar] [CrossRef]
- Gaudet, D.A.; Puchalski, B.J.; Kozub, G.C.; Schaalje, G.B. Susceptibility and resistance in Canadian spring wheat cultivars to common bunt (Tilletia tritici and T. laevis). Can. J. Plant Sci. 1993, 73, 1217–1224. [Google Scholar] [CrossRef]
- Zou, J.; Semagn, K.; Chen, H.; Iqbal, M.; Asif, M.; N’Diaye, A.; Navabi, A.; Perez-Lara, E.; Pozniak, C.; Yang, R.C.; et al. Mapping of QTLs associated with resistance to common bunt, tan spot, leaf rust, and stripe rust in a spring wheat population. Mol. Breed. 2017, 37, 144. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90,000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Beales, J.; Turner, A.; Griffiths, S.; Snape, J.W.; Laurie, D.A. A pseudo-response regulator is misexpressed in the photoperiod insensitive Ppd-D1a mutant of wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 115, 721–733. [Google Scholar] [CrossRef]
- Chen, F.; Gao, M.; Zhang, J.; Zuo, A.; Shang, X.; Cui, D. Molecular characterization of vernalization and response genes in bread wheat from the Yellow and Huai Valley of China. BMC Plant Biol. 2013, 13, 199. [Google Scholar] [CrossRef] [Green Version]
- Ellis, M.H.; Spielmeyer, W.; Gale, K.R.; Rebetzke, G.J.; Richards, R.A. Perfect markers for the Rht-B1b and Rht-D1b dwarfing genes in wheat. Theor. Appl. Genet. 2002, 105, 1038–1042. [Google Scholar] [CrossRef]
- Perez-Lara, E.; Semagn, K.; Chen, H.; Iqbal, M.; N’Diaye, A.; Kamran, A.; Navabi, A.; Pozniak, C.; Spaner, D. QTLs Associated with agronomic traits in the Cutler × AC Barrie spring wheat mapping population using single nucleotide polymorphic markers. PLoS ONE 2016, 11, e0160623. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Bemister, D.H.; Iqbal, M.; Strelkov, S.E.; Spaner, D.M. Mapping genomic regions controlling agronomic traits in spring wheat under conventional and organic managements. Crop Sci. 2020, 60, 2038–2052. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Crossa, J.; Jarquín, D.; Franco, J.; Pérez-Rodríguez, P.; Burgueño, J.; Saint-Pierre, C.; Vikram, P.; Sansaloni, C.; Petroli, C.; Akdemir, D.; et al. Genomic prediction of gene bank wheat landraces. Genes Genomes Genetics 2016, 6, 1819–1834. [Google Scholar] [CrossRef] [Green Version]
- Pérez, P.; De Los Campos, G. Genome-wide regression and prediction with the BGLR statistical package. Genetics 2014, 198, 483–495. [Google Scholar] [CrossRef]
- Goates, B.J. Identification of new pathogenic races of common bunt and dwarf bunt fungi, and evaluation of known races using an expanded set of differential wheat lines. Plant Dis. 2012, 96, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Goates, B.J.; Bockelman, H.E. Identification of new sources of high levels of resistance to dwarf bunt and common bunt among winter wheat landraces in the USDA-ARS national small grains collection. Crop Sci. 2012, 52, 2595–2605. [Google Scholar] [CrossRef]
- McIntosh, R.A.; Dubcovsky, W.J.; Rogers, W.J.; Raupp, W.J. Catalogue of gene symbols for wheat: 2020 supplement. Annu. Wheat Newsl. 2020, 66. Available online: https://wheat.pw.usda.gov/GG3/wgc (accessed on 20 March 2022).
- Delplace, F.; Huard-Chauveau, C.; Dubiella, U.; Khafif, M.; Alvarez, E.; Langin, G.; Roux, F.; Peyraud, R.; Roby, D. Robustness of plant quantitative disease resistance is provided by a decentralized immune network. Proc. Natl. Acad. Sci. USA 2020, 117, 18099. [Google Scholar] [CrossRef]
- Veneault-Fourrey, C.; Rep, M. Quantitative resistance linked to late effectors. New Phytol. 2021, 231, 1301–1303. [Google Scholar] [CrossRef] [PubMed]
- Bemister, D.H.; Semagn, K.; Iqbal, M.; Randhawa, H.; Strelkov, S.E.; Spaner, D.M. Mapping QTL associated with stripe rust, leaf rust, and leaf spotting in a Canadian spring wheat population. Crop Sci. 2019, 59, 650–658. [Google Scholar] [CrossRef]
- Perez-Lara, E.; Semagn, K.; Tran, A.N.; Ciechanowska, I.; Chen, H.; Iqbal, M.; N’Diaye, A.; Pozniak, C.; Strelkov, S.E.; Hucl, P.J.; et al. Population structure and genomewide association analysis of resistance to disease and insensitivity to Ptr toxins in Canadian spring wheat using 90K SNP array. Crop Sci. 2017, 57, 1522–1539. [Google Scholar] [CrossRef]
- Arruda, M.P.; Lipka, A.E.; Brown, P.J.; Krill, A.M.; Thurber, C.; Brown-Guedira, G.; Dong, Y.; Foresman, B.J.; Kolb, F.L. Comparing genomic selection and marker-assisted selection for Fusarium head blight resistance in wheat (Triticum aestivum L.). Mol. Breed. 2016, 36, 84. [Google Scholar] [CrossRef]
- Jarquin, D.; Howard, R.; Crossa, J.; Beyene, Y.; Gowda, M.; Martini, J.W.R.; Covarrubias Pazaran, G.; Burgueño, J.; Pacheco, A.; Grondona, M.; et al. Genomic prediction enhanced sparse testing for multi-environment trials. G3 Genes Genomes Genet. 2020, 10, 2725. [Google Scholar] [CrossRef]
- Heslot, N.; Akdemir, D.; Sorrells, M.E.; Jannink, J.L. Integrating environmental covariates and crop modeling into the genomic selection framework to predict genotype by environment interactions. Theor. Appl. Genet. 2013, 127, 463–480. [Google Scholar] [CrossRef]
- Sukumaran, S.; Crossa, J.; Jarquin, D.; Lopes, M.; Reynolds, M.P. Genomic prediction with pedigree and genotype × environment interaction in spring wheat grown in South and West Asia, North Africa, and Mexico. G3 Genes Genomes Genet. 2017, 7, 481–495. [Google Scholar] [CrossRef] [Green Version]
- Juliana, P.; Singh, R.P.; Braun, H.-J.; Huerta-Espino, J.; Crespo-Herrera, L.; Govindan, V.; Mondal, S.; Poland, J.; Shrestha, S. Genomic Selection for Grain Yield in the CIMMYT Wheat Breeding Program—Status and Perspectives. Front. Plant Sci. 2020, 11, 1418. [Google Scholar] [CrossRef]
- Roorkiwal, M.; Jarquin, D.; Singh, M.K.; Gaur, P.M.; Bharadwaj, C.; Rathore, A.; Howard, R.; Srinivasan, S.; Jain, A.; Garg, V.; et al. Genomic-enabled prediction models using multi-environment trials to estimate the effect of genotype × environment interaction on prediction accuracy in chickpea. Sci. Rep. 2018, 8, 11701. [Google Scholar] [CrossRef] [Green Version]
- Jarquín, D.; Lemes da Silva, C.; Gaynor, R.C.; Poland, J.; Fritz, A.; Howard, R.; Battenfield, S.; Crossa, J. Increasing genomic-enabled prediction accuracy by modeling genotype × environment interactions in Kansas wheat. Plant Genome 2017, 10, plantgenome2016.2012.0130. [Google Scholar] [CrossRef] [Green Version]
- Emebiri, L.; Singh, S.; Tan, M.-K. DArTseq physical mapping of QTLs linked to Karnal bunt (Tilletia indica) resistance in two historical wheat populations. Plant Pathol. 2021, 70, 1779–1789. [Google Scholar] [CrossRef]
- Zhang, A.; Wang, H.; Beyene, Y.; Semagn, K.; Liu, Y.; Cao, S.; Cui, Z.; Ruan, Y.; Burgueño, J.; San Vicente, F.; et al. Effect of trait heritability, training population size and marker density on genomic prediction accuracy estimation in 22 bi-parental tropical maize populations. Front. Plant Sci. 2017, 8, 1916. [Google Scholar] [CrossRef] [PubMed]
- Norman, A.; Taylor, J.; Edwards, J.; Kuchel, H. Optimising genomic selection in wheat: Effect of marker density, population size and population structure on prediction accuracy. G3 Genes Genomes Genet. 2018, 8, 2889–2899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutkoski, J.; Benson, J.; Jia, Y.; Brown-Guedira, G.; Jannink, J.-L.; Sorrells, M. Evaluation of genomic prediction methods for Fusarium head blight resistance in wheat. Plant Genome 2012, 5, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Mirdita, V.; Liu, G.; Zhao, Y.; Miedaner, T.; Longin, C.F.H.; Gowda, M.; Mette, M.F.; Reif, J.C. Genetic architecture is more complex for resistance to Septoria tritici blotch than to Fusarium head blight in Central European winter wheat. BMC Genom. 2015, 16, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population 1 | Trait | No. of Environments | Correlation 2 | Individual Environments 3 | All Environments 3 | Broad-Sense Heritability | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Range | Mean | Range | Mean | Std | Range | Mean | Std | ||||
| BVC | Stripe rust | 8 | 0.10–0.81 | 0.52 | 1.0–9.0 | 3.7 | 2.5 | 1.2–8.1 | 3.76 | 1.80 | 0.89 |
| Leaf rust | 4 | 0.43–0.75 | 0.55 | 1.0–9.0 | 3.6 | 2.3 | 1.0–8.1 | 3.52 | 1.78 | 0.78 | |
| Leaf spot | 3 | 0.07–0.28 | 0.09 | 1.0–9.0 | 4.1 | 2.0 | 2.2–6.8 | 4.12 | 0.92 | 0.22 | |
| Common bunt | 2 | 0.50–0.64 | 0.57 | 1.0–9.0 | 2.5 | 1.8 | 1.0–7.9 | 2.48 | 1.61 | 0.69 | |
| FHB incidence | 6 | 0.12–0.56 | 0.28 | 1.0–9.0 | 5.9 | 2.8 | 2.4–7.5 | 5.81 | 0.74 | 0.29 | |
| FHB severity | 6 | 0.11–0.67 | 0.41 | 1.0–9.0 | 3.8 | 2.1 | 1.8–6.8 | 4.15 | 0.94 | 0.58 | |
| FHB index | 6 | 0.10–0.68 | 0.42 | 0.5–82.1 | 26.3 | 20.4 | 3.7–49.6 | 25.97 | 9.11 | 0.50 | |
| PAC | Stripe rust | 8 | 0.14–0.66 | 0.42 | 1.0–9.0 | 2.1 | 1.4 | 1.1–5.7 | 2.07 | 0.79 | 0.72 |
| Leaf rust | 3 | 0.45–0.99 | 0.64 | 1.0–6.9 | 1.7 | 1.1 | 1.0–5.8 | 1.74 | 0.94 | 0.82 | |
| Leaf spot | 4 | 0.52–1.00 | 0.66 | 1.4–8.3 | 4.4 | 1.3 | 2.3–7.1 | 4.45 | 1.10 | 0.85 | |
| Common bunt | 3 | 0.76–1.00 | 0.84 | 1.0–7.5 | 1.6 | 1.1 | 1.0–6.0 | 1.59 | 1.06 | 0.90 | |
| ACG | Stripe rust | 3 | 0.38–0.50 | 0.46 | 1.0–9.0 | 3.9 | 2.0 | 1.5–7.9 | 3.85 | 1.33 | 0.62 |
| Leaf rust | 3 | 0.18–0.29 | 0.24 | 1.0–8.5 | 4.2 | 1.6 | 1.9–6.1 | 3.44 | 0.82 | 0.44 | |
| Leaf spot | 3 | 0.10–0.44 | 0.24 | 1.0–9.0 | 6.2 | 1.7 | 3.0–8.7 | 6.00 | 1.30 | 0.57 | |
| Common bunt | 3 | 0.22–0.37 | 0.32 | 1.0–7.0 | 2.3 | 1.4 | 1.0–4.9 | 1.79 | 0.82 | 0.52 | |
| CV1 | CV2 | CV0 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M1 | M2 | M3 | M1 | M2 | M3 | M1 | M2 | M3 | |||||||||||
| Population 1 | Trait 2 | Mean | Std 3 | Mean | Std | Mean | Std | Mean | Std | Mean | Std | Mean | Std | Mean | Std | Mean | Std | Mean | Std |
| BVC | Bunt | −0.11 | 0.07 | 0.43 | 0.02 | 0.44 | 0.03 | 0.56 | 0.03 | 0.62 | 0.02 | 0.62 | 0.03 | 0.62 | 0.02 | 0.63 | 0.00 | 0.62 | 0.04 |
| FHB inc. | −0.13 | 0.06 | 0.22 | 0.09 | 0.27 | 0.13 | 0.41 | 0.12 | 0.43 | 0.13 | 0.45 | 0.13 | 0.44 | 0.14 | 0.45 | 0.14 | 0.45 | 0.14 | |
| FHB index | −0.10 | 0.07 | 0.39 | 0.16 | 0.40 | 0.18 | 0.56 | 0.13 | 0.58 | 0.15 | 0.63 | 0.16 | 0.59 | 0.16 | 0.59 | 0.17 | 0.60 | 0.16 | |
| FHB sev. | −0.11 | 0.07 | 0.42 | 0.14 | 0.41 | 0.17 | 0.58 | 0.15 | 0.59 | 0.16 | 0.62 | 0.18 | 0.60 | 0.17 | 0.61 | 0.18 | 0.61 | 0.18 | |
| Lr | −0.09 | 0.08 | 0.49 | 0.11 | 0.48 | 0.12 | 0.58 | 0.09 | 0.64 | 0.10 | 0.64 | 0.09 | 0.63 | 0.13 | 0.65 | 0.12 | 0.59 | 0.05 | |
| Ls | −0.13 | 0.05 | 0.18 | 0.20 | 0.38 | 0.16 | 0.07 | 0.10 | 0.21 | 0.18 | 0.41 | 0.15 | 0.12 | 0.10 | 0.11 | 0.14 | 0.14 | 0.11 | |
| Yr | −0.10 | 0.07 | 0.48 | 0.08 | 0.49 | 0.09 | 0.75 | 0.07 | 0.76 | 0.07 | 0.76 | 0.06 | 0.77 | 0.08 | 0.76 | 0.07 | 0.77 | 0.08 | |
| PAC | Bunt | −0.10 | 0.10 | 0.31 | 0.06 | 0.29 | 0.07 | 0.84 | 0.07 | 0.84 | 0.06 | 0.84 | 0.06 | 0.87 | 0.09 | 0.87 | 0.09 | 0.87 | 0.09 |
| Lr | −0.06 | 0.07 | 0.31 | 0.08 | 0.29 | 0.11 | 0.70 | 0.19 | 0.71 | 0.20 | 0.70 | 0.20 | 0.77 | 0.27 | 0.73 | 0.24 | 0.77 | 0.27 | |
| Ls | −0.03 | 0.08 | 0.33 | 0.04 | 0.32 | 0.07 | 0.73 | 0.12 | 0.73 | 0.11 | 0.74 | 0.10 | 0.75 | 0.14 | 0.74 | 0.13 | 0.75 | 0.13 | |
| Yr | −0.09 | 0.07 | 0.26 | 0.07 | 0.26 | 0.10 | 0.54 | 0.09 | 0.54 | 0.09 | 0.55 | 0.08 | 0.56 | 0.10 | 0.56 | 0.08 | 0.56 | 0.10 | |
| ACG | Bunt | −0.14 | 0.07 | 0.02 | 0.07 | 0.02 | 0.09 | 0.39 | 0.05 | 0.39 | 0.06 | 0.34 | 0.08 | 0.41 | 0.04 | 0.42 | 0.06 | 0.42 | 0.05 |
| Lr | −0.14 | 0.07 | 0.13 | 0.05 | 0.11 | 0.06 | 0.27 | 0.04 | 0.29 | 0.03 | 0.25 | 0.04 | 0.30 | 0.03 | 0.33 | 0.01 | 0.32 | 0.02 | |
| Ls | −0.16 | 0.06 | 0.04 | 0.10 | 0.08 | 0.07 | 0.28 | 0.12 | 0.28 | 0.13 | 0.31 | 0.08 | 0.32 | 0.14 | 0.30 | 0.16 | 0.32 | 0.15 | |
| Yr | −0.12 | 0.09 | 0.25 | 0.06 | 0.29 | 0.08 | 0.53 | 0.05 | 0.54 | 0.05 | 0.56 | 0.06 | 0.54 | 0.06 | 0.53 | 0.05 | 0.54 | 0.05 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semagn, K.; Iqbal, M.; Jarquin, D.; Crossa, J.; Howard, R.; Ciechanowska, I.; Henriquez, M.A.; Randhawa, H.; Aboukhaddour, R.; McCallum, B.D.; et al. Genomic Predictions for Common Bunt, FHB, Stripe Rust, Leaf Rust, and Leaf Spotting Resistance in Spring Wheat. Genes 2022, 13, 565. https://doi.org/10.3390/genes13040565
Semagn K, Iqbal M, Jarquin D, Crossa J, Howard R, Ciechanowska I, Henriquez MA, Randhawa H, Aboukhaddour R, McCallum BD, et al. Genomic Predictions for Common Bunt, FHB, Stripe Rust, Leaf Rust, and Leaf Spotting Resistance in Spring Wheat. Genes. 2022; 13(4):565. https://doi.org/10.3390/genes13040565
Chicago/Turabian StyleSemagn, Kassa, Muhammad Iqbal, Diego Jarquin, José Crossa, Reka Howard, Izabela Ciechanowska, Maria Antonia Henriquez, Harpinder Randhawa, Reem Aboukhaddour, Brent D. McCallum, and et al. 2022. "Genomic Predictions for Common Bunt, FHB, Stripe Rust, Leaf Rust, and Leaf Spotting Resistance in Spring Wheat" Genes 13, no. 4: 565. https://doi.org/10.3390/genes13040565
APA StyleSemagn, K., Iqbal, M., Jarquin, D., Crossa, J., Howard, R., Ciechanowska, I., Henriquez, M. A., Randhawa, H., Aboukhaddour, R., McCallum, B. D., Brûlé-Babel, A. L., Navabi, A., N’Diaye, A., Pozniak, C., & Spaner, D. (2022). Genomic Predictions for Common Bunt, FHB, Stripe Rust, Leaf Rust, and Leaf Spotting Resistance in Spring Wheat. Genes, 13(4), 565. https://doi.org/10.3390/genes13040565