
Deregulated Clusterin as a Marker of Bone Fragility: New Insights into the Pathophysiology of Osteoporosis

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Clinical and Biochemical Parameters
2.3. Specimen Collection
2.4. Human Osteoblast Primary Cell Culture
2.5. RNA Extraction and qRT-PCR Analysis of Secreted Clusterin Expression
2.6. Immunohistochemical Analysis
2.7. Quantitative Measurement of CLU in Human Plasma
2.8. Genotyping Analysis of rs11136000
2.9. Statistical Analyses
3. Results
3.1. Clinical Characteristics of Individuals Included in the Study
3.2. Secreted Clusterin Expression Level Is Significantly Upregulated in Osteoblasts and PBMCs from OP Patients
3.3. Immunohistochemical Analysis
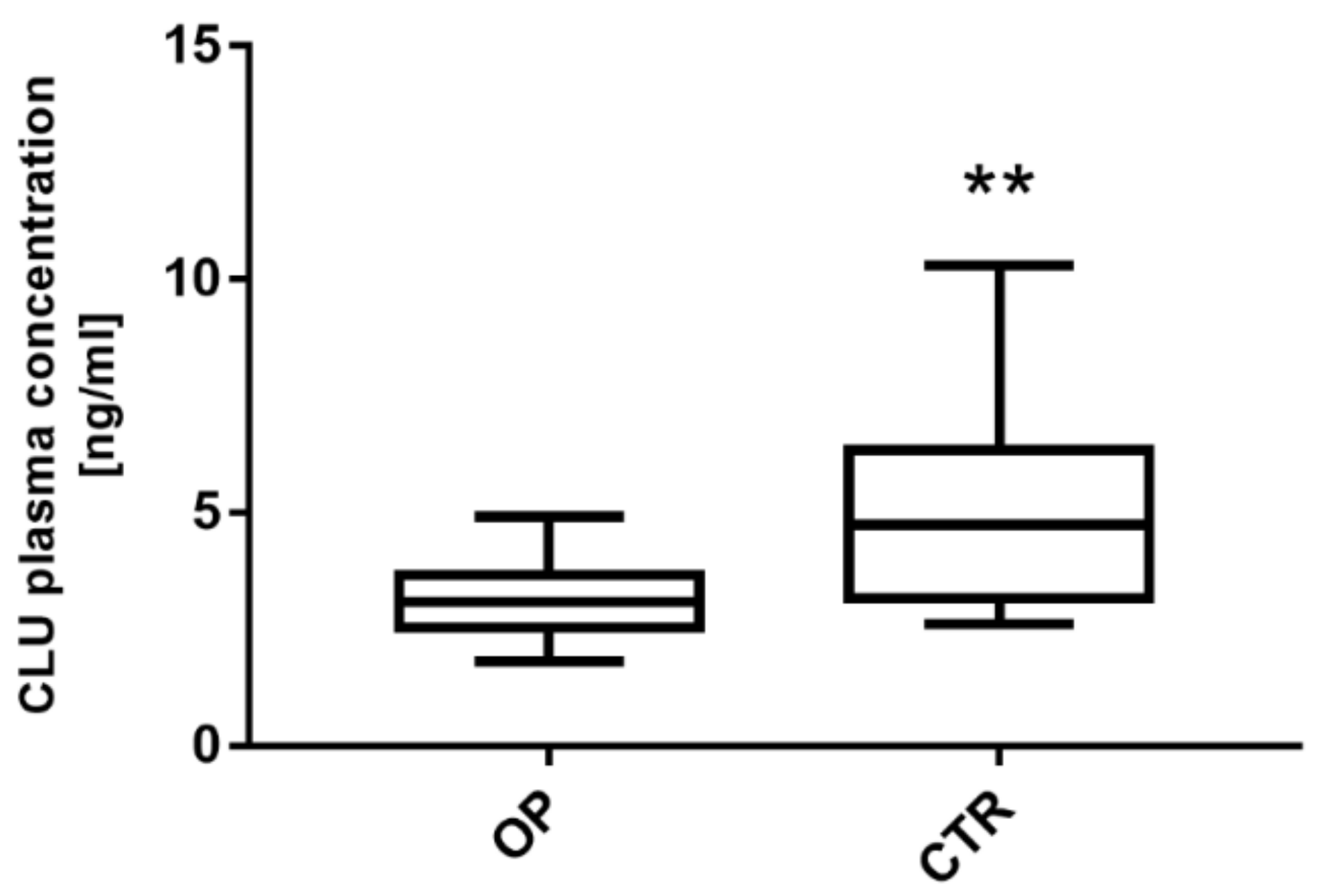
3.4. Clusterin Expression Level Is Significantly Downregulated in Plasma from OP Patients
3.5. The rs1136000 Polymorphism Is Not Associated with the Expression Levels of CLU
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pucci, S.; Bonanno, E.; Pichiorri, F.; Angeloni, C.; Spagnoli, L.G. Modulation of different clusterin isoforms in human colon tumorigenesis. Oncogene 2004, 23, 2298–2304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pucci, S.; Bonanno, E.; Sesti, F.; Mazzarelli, P.; Mauriello, A.; Ricci, F.; Zoccai, G.B.; Rulli, F.; Galatà, G.; Spagnoli, L.G. Clusterin in stool: A new biomarker for colon cancer screening? Am. J. Gastroenterol. 2009, 104, 2807–2815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.-T.; Tan, L. The role of clusterin in Alzheimer’s disease: Pathways, pathogenesis, and therapy. Mol. Neurobiol. 2012, 45, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rivera, C.; Garcia, M.M.; Molina-Álvarez, M.; González-Martín, C.; Goicoechea, C. Clusterin: Always protecting. Synthesis, function and potential issues. Biomed. Pharmacother. 2021, 134, 111174. [Google Scholar] [CrossRef]
- Xiu, P.; Dong, X.-F.; Li, X.-P.; Li, J. Clusterin: Review of research progress and looking ahead to direction in hepatocellular carcinoma. World J. Gastroenterol. 2015, 21, 8262. [Google Scholar] [CrossRef]
- Garcia-Aranda, M.; Serrano, A.; Redondo, M. Regulation of Clusterin Gene Expression. Curr. Protein Pept. Sci. 2018, 19, 612–622. [Google Scholar] [CrossRef]
- Pucci, S.; Greggi, C.; Polidoro, C.; Piro, M.C.; Celi, M.; Feola, M.; Gasbarra, E.; Iundusi, R.; Mastrangeli, F.; Novelli, G.; et al. Clusterin silencing restores myoblasts viability and down modulates the inflammatory process in osteoporotic disease. J. Transl. Med. 2019, 17, 118. [Google Scholar] [CrossRef]
- Abdallah, B.M.; Alzahrani, A.M.; Kassem, M. Secreted Clusterin protein inhibits osteoblast differentiation of bone marrow mesenchymal stem cells by suppressing ERK1/2 signaling pathway. Bone 2018, 110, 221–229. [Google Scholar] [CrossRef]
- Balcar, V.J.; Zeman, T.; Janout, V.; Janoutová, J.; Lochman, J.; Šerý, O. Single Nucleotide Polymorphism rs11136000 of CLU Gene (Clusterin, ApoJ) and the Risk of Late-Onset Alzheimer’s Disease in a Central European Population. Neurochem. Res. 2020, 46, 411–422. [Google Scholar] [CrossRef]
- Chen, L.H.; Heng Mak, T.S.; Fan, Y.; Yin Ho, D.T.; Sham, P.C.; Chu, L.W.; Song, Y.Q. Associations between CLU polymorphisms and memory performance: The role of serum lipids in Alzheimer’s disease. J. Psychiatr. Res. 2020, 129, 281–288. [Google Scholar] [CrossRef]
- Zhu, R.; Liu, X.; He, Z. Association between CLU gene rs11136000 polymorphism and Alzheimer’s disease: An updated meta-analysis. Neurol. Sci. 2018, 39, 679–689. [Google Scholar] [CrossRef]
- Harold, D.; Abraham, R.; Hollingworth, P.; Sims, R.; Gerrish, A.; Hamshere, M.L.; Pahwa, J.S.; Moskvina, V.; Dowzell, K.; Williams, A.; et al. Genome-wide association study identifies variants at CLU and PICALM associated with Alzheimer’s disease. Nat. Genet. 2009, 41, 1088–1093. [Google Scholar] [CrossRef] [Green Version]
- Cai, R.; Han, J.; Sun, J.; Huang, R.; Tian, S.; Shen, Y.; Dong, X.; Xia, W.; Wang, S. Plasma Clusterin and the CLU Gene rs11136000 Variant Are Associated with Mild Cognitive Impairment in Type 2 Diabetic Patients. Front. Aging Neurosci. 2016, 8, 179. [Google Scholar] [CrossRef] [Green Version]
- Tarquini, C.; Pucci, S.; Scioli, M.G.; Doldo, E.; Agostinelli, S.; D’Amico, F.; Bielli, A.; Ferlosio, A.; Caredda, E.; Tarantino, U.; et al. Clusterin exerts a cytoprotective and antioxidant effect in human osteoarthritic cartilage. Aging 2020, 12, 10129–10146. [Google Scholar] [CrossRef]
- Connor, J.R.; Kumar, S.; Sathe, G.; Mooney, J.; O’Brien, S.P.; Mui, P.; Murdock, P.R.; Gowen, M.; Lark, M.W. Clusterin expression in adult human normal and osteoarthritic articular cartilage. Osteoarthr. Cartil. 2001, 9, 727–737. [Google Scholar] [CrossRef] [Green Version]
- Fandridis, E.; Apergis, G.; Korres, D.S.; Nikolopoulos, K.; Zoubos, A.B.; Papassideri, I.; Trougakos, I.P. Increased expression levels of apolipoprotein J/clusterin during primary osteoarthritis. In Vivo 2011, 25, 745–749. [Google Scholar]
- Ma, J.; Gao, W.; Gao, J. sCLU as prognostic biomarker and therapeutic target in osteosarcoma. Bioengineered 2019, 10, 229. [Google Scholar] [CrossRef] [Green Version]
- Scimeca, C.; Centofanti, F.; Celi, M.; Gasbarra, E.; Novelli, G.; Botta, A.; Tarantino, U. Vitamin D Receptor in Muscle Atrophy of Elderly Patients: A Key Element of Osteoporosis-Sarcopenia Connection. Aging Dis. 2018, 9, 952–964. [Google Scholar] [CrossRef] [Green Version]
- Celi, M.; Rao, C.; Scialdoni, A.; Tempesta, V.; Gasbarra, E.; Pistillo, P.; Tarantino, U. Bone mineral density evaluation in osteoporosis: Why yes and why not? Aging Clin. Exp. Res. 2013, 25, 47–49. [Google Scholar] [CrossRef]
- Schürmann, B.; Wiese, B.; Bickel, H.; Weyerer, S.; Riedel-Heller, S.G.; Pentzek, M.; Bachmann, C.; Williams, J.; Bussche, H.; Maier, W.; et al. Association of the Alzheimer’s disease clusterin risk allele with plasma clusterin concentration. J. Alzheimer’s Dis. 2011, 25, 421–424. [Google Scholar] [CrossRef]
- Uda, Y.; Azab, E.; Sun, N.; Shi, C.; Pajevic, P.D. Osteocyte Mechanobiology. Curr. Osteoporos. Rep. 2017, 15, 318–325. [Google Scholar] [CrossRef]
- Cariati, I.; Bonanni, R.; Onorato, F.; Mastrogregori, A.; Rossi, D.; Iundusi, R.; Gasbarra, E.; Tancredi, V.; Tarantino, U. Role of Physical Activity in Bone-Muscle Crosstalk: Biological Aspects and Clinical Implications. J. Funct. Morphol. Kinesiol. 2021, 6, 55. [Google Scholar] [CrossRef]
- Kropáčková, T.; Šléglová, O.; Růžičková, O.; Vencovský, J.; Pavelka, K.; Šenolt, L. Lower serum clusterin levels in patients with erosive hand osteoarthritis are associated with more pain. BMC Musculoskelet. Disord. 2018, 19, 264. [Google Scholar] [CrossRef] [Green Version]
- Trougakos, I.P.; Petropoulou, C.; Franceschi, C.; Gonos, E.S. Reduced expression levels of the senescence biomarker clusterin/apolipoprotein j in lymphocytes from healthy centenarians. Ann. N. Y. Acad. Sci. 2006, 1067, 294–300. [Google Scholar] [CrossRef]
- Kimball, J.S.; Johnson, J.P.; Carlson, D.W.A. Oxidative Stress and Osteoporosis. J. Bone Jt. Surg. Am. 2021, 103, 1451–1461. [Google Scholar] [CrossRef]
- Kang, S.W.; Yoon, S.Y.; Park, J.Y.; Kim, D.H. Unglycosylated clusterin variant accumulates in the endoplasmic reticulum and induces cytotoxicity. Int. J. Biochem. Cell Biol. 2013, 45, 221–231. [Google Scholar] [CrossRef]
- Briggs, A.M.; Woolf, A.D.; Dreinhöfer, K.; Homb, N.; Hoy, D.G.; Kopansky-Giles, D.; Åkesson, K.; March, L. Reducing the global burden of musculoskeletal conditions. Bull. World Health Organ. 2018, 96, 366. [Google Scholar] [CrossRef]
- Gheno, R.; Cepparo, J.M.; Rosca, C.E.; Cotten, A. Musculoskeletal disorders in the elderly. J. Clin. Imaging Sci. 2012, 2, 39. [Google Scholar] [CrossRef]
- Rashki Kemmak, A.; Rezapour, A.; Jahangiri, R.; Nikjoo, S.; Farabi, H.; Soleimanpour, S. Economic burden of osteoporosis in the world: A systematic review. Med. J. Islam. Repub. Iran 2020, 34, 154. [Google Scholar] [CrossRef]
- Visconti, V.V.; Fittipaldi, S.; Ciuffi, S.; Marini, F.; Isaia, G.; D’Amelio, P.; Migliaccio, S.; Marcocci, C.; Minisola, S.; Nuti, R.; et al. Circulating Long Non-Coding RNA GAS5 Is Overexpressed in Serum from Osteoporotic Patients and Is Associated with Increased Risk of Bone Fragility. Int. J. Mol. Sci. 2020, 21, 6930. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | OP (n = 30) | CTRs (n = 30) | p Value |
|---|---|---|---|
| Age (years) | 63 ± 15.3 | 45.3 ± 17.8 | ** (p < 0.01) |
| BMI (Kg/cm2) | 24.6 ± 4.5 | 25.8 ± 2.3 | NS (p = 0.494) |
| t-score L1–L4 | −1.0 ± 1.2 | 0.3 ± 1.0 | ** (p < 0.01) |
| t-score total femur | −1.4 ± 1.1 | 1.0 ± 0.8 | **** (p < 0.0001) |
| t-score femoral neck | −1.9 ± 0.6 | 0.2 ± 0.7 | **** (p < 0.0001) |
| BMD L1-L4 (g/cm2) | 1.0 ± 0.1 | 1.2 ± 0.1 | ** (p < 0.01) |
| BMD total femur (g/cm2) | 0.8 ± 0.1 | 1.2 ± 0.1 | **** (p < 0.0001) |
| BMD femoral neck (g/cm2) | 0.7 ± 0.1 | 1.1 ± 0.1 | *** (p < 0.001) |
| Calcium (mg/dL) | 8.3 ± 0.4 | 8.4 ± 0.3 | NS (p = 0.614) |
| PTH (pg/mL) | 98.9 ± 70 | 73.3 ± 18.5 | NS (p = 0.410) |
| 25-(OH)-Vit D (ng/mL) | 16.2 ± 11 | 23.5 ± 4.5 | * (p < 0.05) |
| Gene | Sequence (5′–3′) | |
|---|---|---|
| CLU | Forward | ACAGGGTGCCGCTGACC |
| Reverse | CAGCAGAGTCTTCATCATGC | |
| OCN | Forward | ACACTCCTCGCCCTATTG |
| Reverse | GATGTGGTCAGCCAACTC | |
| GAPDH | Forward | GATCATCAGCAATGCCTCCTG |
| Reverse | GTCTTCTGGGTGGCAGTGAT | |
| B2M | Forward | CTGGAACGGTGAAGGTGACA |
| Reverse | AAGGGACTTCCTGTAACAATGCA | |
| β-actin | Forward | ACTCCATGCCCAGGAAGGAA |
| Reverse | GAGATGGCCACGGCTGCTT |
| Genotype | p | Allele | p | Odds Ratio (95% CI) | ||||
|---|---|---|---|---|---|---|---|---|
| C/C | C/T | T/T | C | T | ||||
| OP (n = 30) | 10 (33.3%) | 18 (60%) | 2 (6.7%) | 0.162 | 38 (63.3%) | 22 (36.7%) | 1 | 1 (0.563–1.777) |
| CTRs (n = 30) | 11(37%) | 15 (50%) | 4 (13%) | 38 (63.3%) | 22 (36.7%) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visconti, V.V.; Greggi, C.; Cariati, I.; Gasperini, B.; Mastrogregori, A.; Botta, A.; Tarantino, U. Deregulated Clusterin as a Marker of Bone Fragility: New Insights into the Pathophysiology of Osteoporosis. Genes 2022, 13, 652. https://doi.org/10.3390/genes13040652
Visconti VV, Greggi C, Cariati I, Gasperini B, Mastrogregori A, Botta A, Tarantino U. Deregulated Clusterin as a Marker of Bone Fragility: New Insights into the Pathophysiology of Osteoporosis. Genes. 2022; 13(4):652. https://doi.org/10.3390/genes13040652
Chicago/Turabian StyleVisconti, Virginia Veronica, Chiara Greggi, Ida Cariati, Beatrice Gasperini, Ambra Mastrogregori, Annalisa Botta, and Umberto Tarantino. 2022. "Deregulated Clusterin as a Marker of Bone Fragility: New Insights into the Pathophysiology of Osteoporosis" Genes 13, no. 4: 652. https://doi.org/10.3390/genes13040652
APA StyleVisconti, V. V., Greggi, C., Cariati, I., Gasperini, B., Mastrogregori, A., Botta, A., & Tarantino, U. (2022). Deregulated Clusterin as a Marker of Bone Fragility: New Insights into the Pathophysiology of Osteoporosis. Genes, 13(4), 652. https://doi.org/10.3390/genes13040652