
Partial fads2 Gene Knockout Diverts LC-PUFA Biosynthesis via an Alternative Δ8 Pathway with an Impact on the Reproduction of Female Zebrafish (Danio rerio)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. sgRNA Design and sgRNA/CRISPR/Cas9 Complex Preparation
2.3. Fish Source and In Vitro Fertilization
2.4. Microinjection of the sgRNA/CRISPR/Cas9 Complex into the In Vitro Fertilized Eggs, and Fish Maintenance
2.5. Genomic DNA Isolation, the Detection and Quantification of the CRISPR/Cas9 System-Mediated Gene Editing
2.6. The Eggs’ LC-PUFA FAMEs Profiles and Statistical Analyses
2.7. Phenotype of the Fertilized Eggs Produced by Crispant Females
3. Results
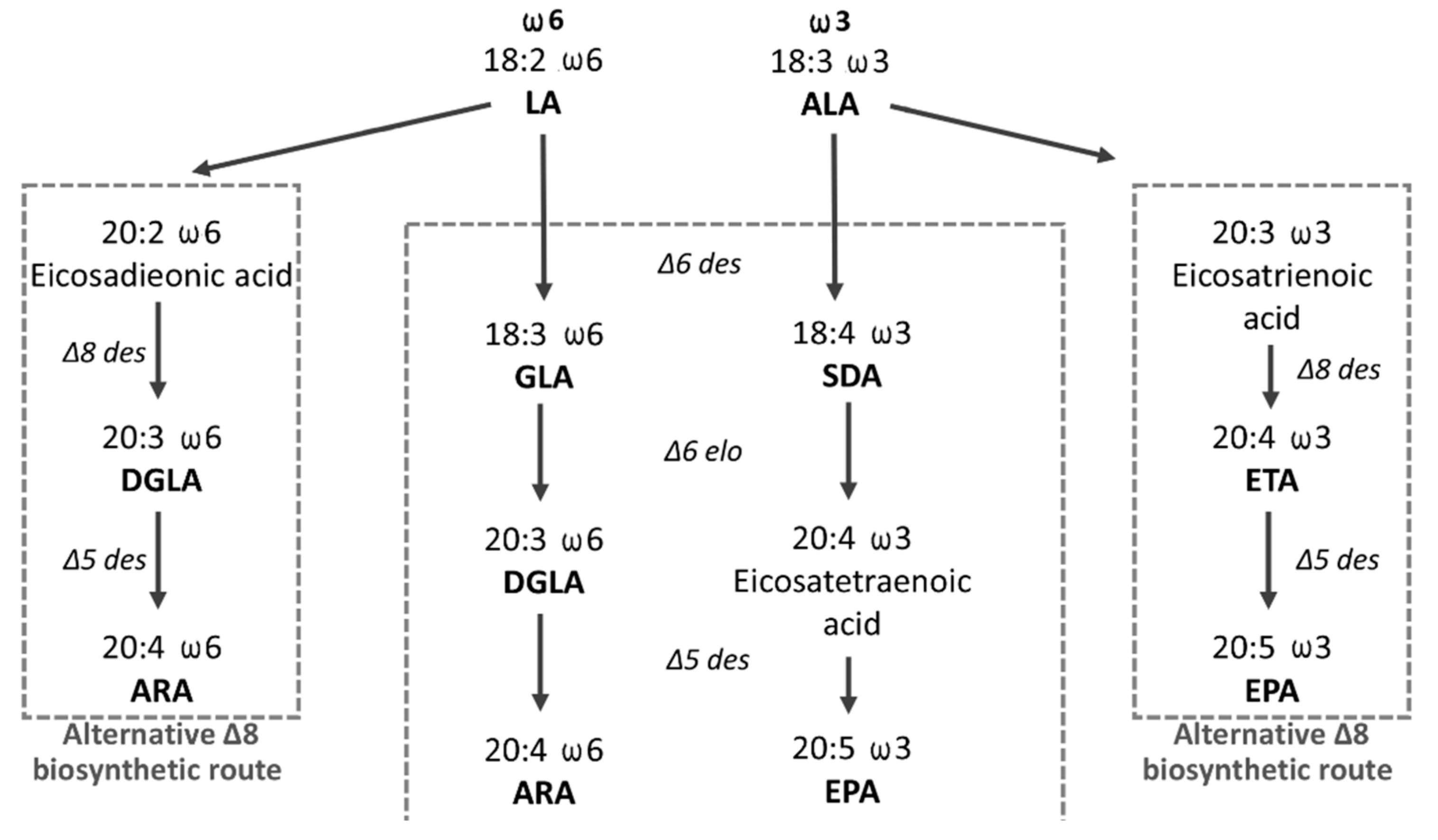
3.1. sgRNA Design
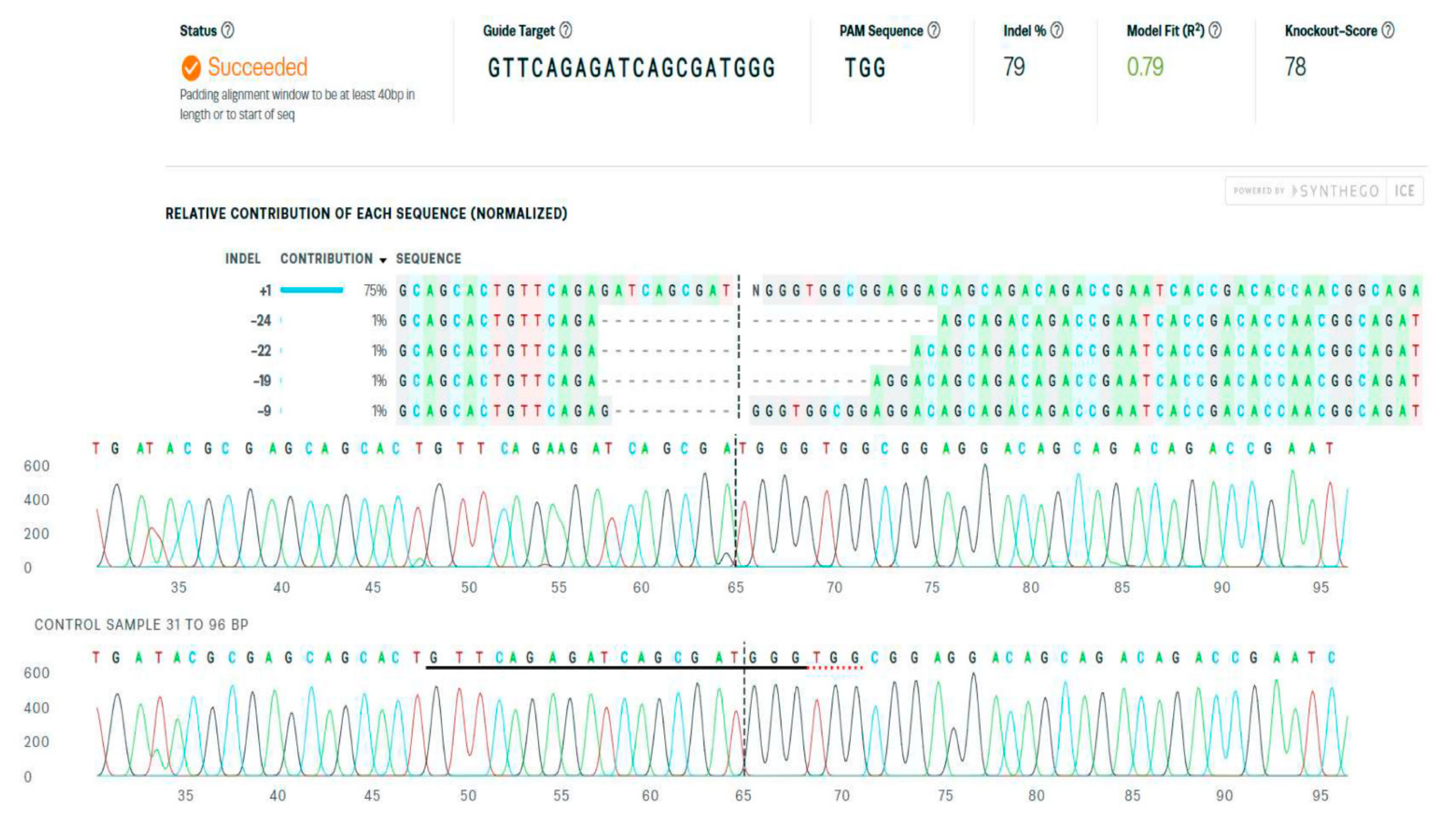
3.2. The Detection and Quantification of the CRISPR/Cas9 System-Mediated Gene Editing
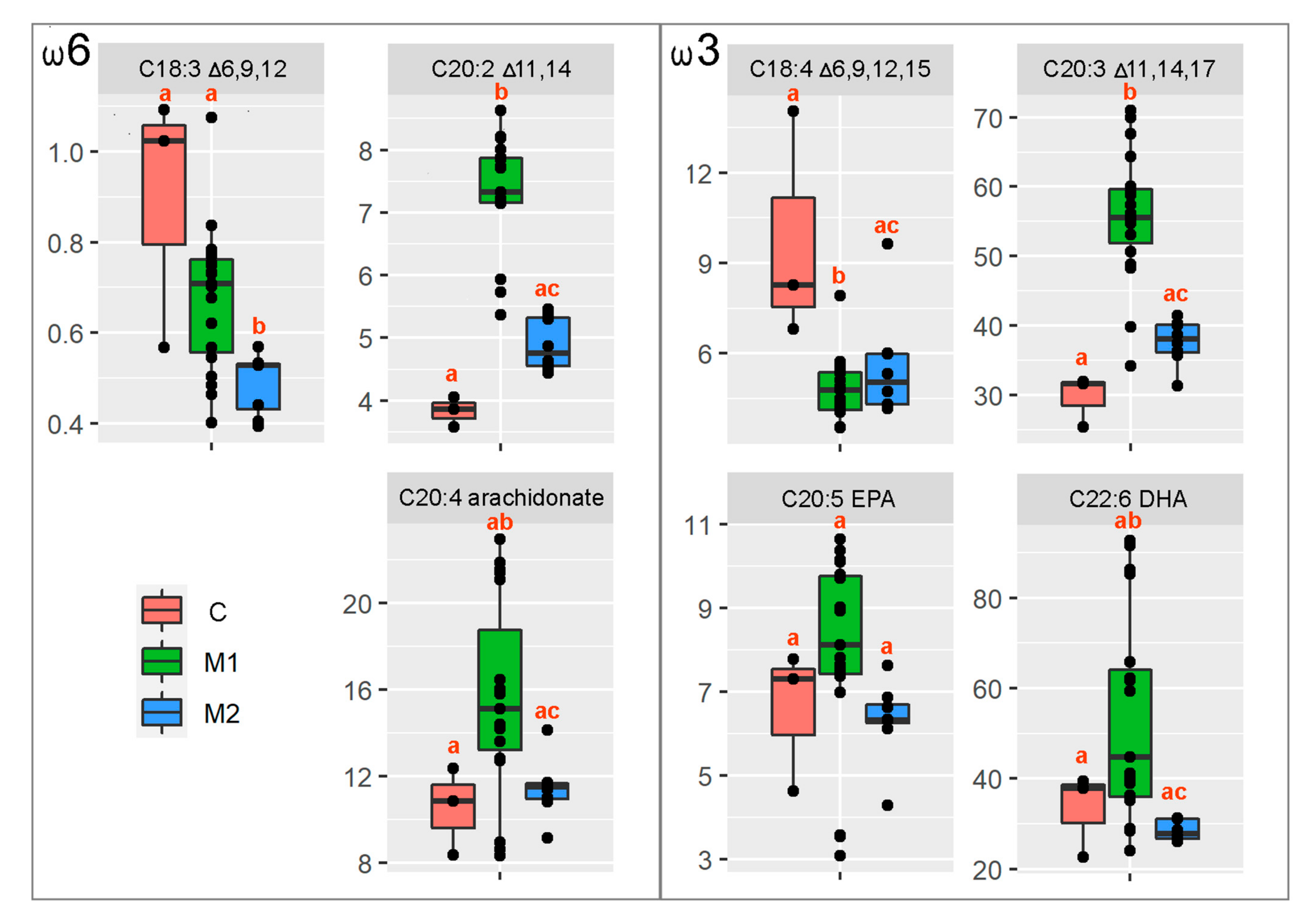
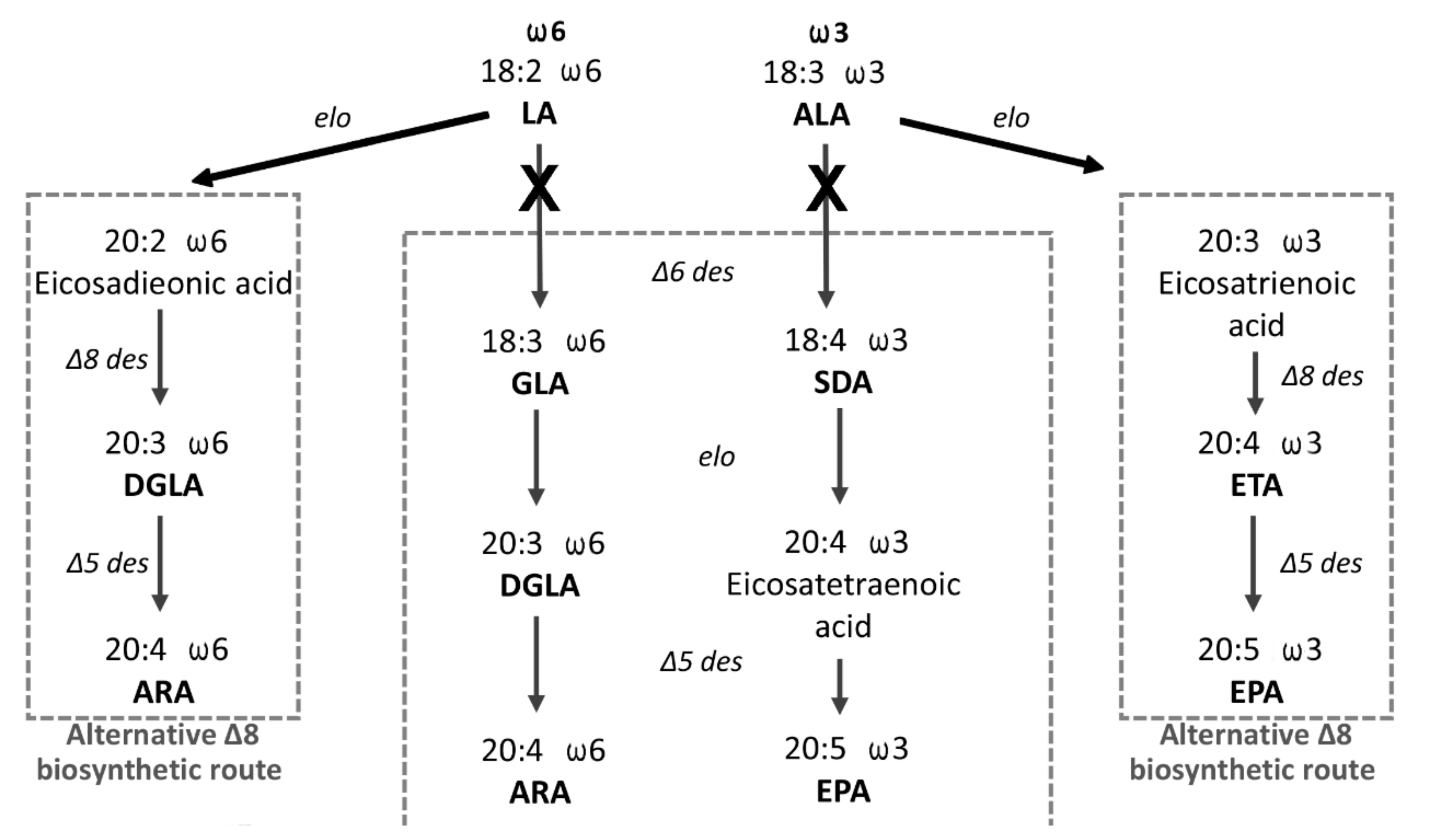
3.3. Egg Quality, LC-PUFA FAME Profile Analysis and Statistical Analysis
3.4. Altered Phenotype of the Embryos from a Fads2 Mutant Female x WT Male Inter-Crosses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schuchardt, J.P.; Huss, M.; Stauss-Grabo, M.; Hahn, A. Significance of long-chain polyunsaturated fatty acids (PUFAs) for the development and behaviour of children. Eur. J. Pediatr. 2010, 169, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.V.; Tocher, D. Biosynthesis of polyunsaturated fatty acids in aquatic ecosystems: General pathways and new directions. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M.T., Kainz, M., Eds.; Springer: New York, NY, USA, 2009; pp. 211–236. [Google Scholar]
- Monroig, Ó.; Rotllant, J.; Sánchez, E.; Cerdá-Reverter, J.M.; Tocher, D.R. Expression of long-chain polyunsaturated fatty acid (LC-PUFA) biosynthesis genes during zebrafish Danio rerio early embryogenesis. Biochim. Biophys. Acta 2009, 1791, 1093–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoffel, W.; Holz, B.; Jenke, B.; Binczek, E.; Günter, R.H.; Kiss, C.; Karakesisoglou, I.; Thevis, M.; Weber, A.-A.; Arnhold, S.; et al. Δ6-Desaturase (FADS2) deficiency unveils the role of ω3- and ω6-polyunsaturated fatty acids. EMBO J. 2008, 27, 2281–2292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, B.; Hirsh, D. Receptor-mediated Endocytosis in the Caenorhabditis elegans Oocyte. Mol. Biol. Cell 1999, 10, 4311–4326. [Google Scholar] [CrossRef] [Green Version]
- Brock, T.J.; Browse, J.; Watts, J.L. Fatty acid desaturation and the regulation of adiposity in Caenorhabditis elegans. Genetics 2007, 176, 865–875. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.-W.; Yi, Y.-H.; Chien, C.-H.; Hsiung, K.-C.; Ma, T.-H.; Lin, Y.-C.; Lo, S.J.; Chang, T.-C. Specific polyunsaturated fatty acids modulate lipid delivery and oocyte development in C. elegans revealed by molecular-selective label-free imaging. Sci. Rep. 2016, 6, 32021. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.V.; Tocher, D. Molecular species composition of the major phospholipids in brain and retina from rainbow trout (Salmo gairdneri). Occurrence of high levels of di-(n-3)polyunsaturated fatty acid species. Biochem. J. 1989, 264, 909–915. [Google Scholar] [CrossRef] [Green Version]
- Tocher, D.R.; Betancor, M.B.; Sprague, M.; Olsen, R.E.; Napier, J.A. Omega-3 Long-Chain Polyunsaturated Fatty Acids, EPA and DHA: Bridging the Gap between Supply and Demand. Nutrients 2019, 11, 89. [Google Scholar] [CrossRef] [Green Version]
- Gil, A.; Ramirez, M.; Gil, M. Role of long-chain polyunsaturated fatty acids in infant nutrition. Eur. J. Clin. Nutr. 2003, 57, S31–S34. [Google Scholar] [CrossRef]
- Zárate, R.; El Jaber-Vazdekis, N.; Tejera, N.; Pérez, J.A.P.; Rodríguez, C. Significance of long chain polyunsaturated fatty acids in human health. Clin. Transl. Med. 2017, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Kabeya, N.; Yevzelman, S.; Oboh, A.; Tocher, D.R.; Monroig, O. Essential fatty acid metabolism and requirements of the cleaner fish, ballan wrasse Labrus bergylta: Defining pathways of long-chain polyunsaturated fatty acid biosynthesis. Aquaculture 2018, 488, 199–206. [Google Scholar] [CrossRef]
- Ferraz, R.; Kabeya, N.; Lopes-Marques, M.; Machado, A.M.; Ribeiro, R.A.; Salaro, A.L.; Ozorio, R.; Castro, L.F.C.; Monroig, Ó. A complete enzymatic capacity for long-chain polyunsaturated fatty acid biosynthesis is present in the Amazonian teleost tambaqui, Colossoma macropomum. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2019, 227, 90–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, D.; Monroig, Ó.; Galindo, A.; Betancor, M.; Pérez, J.A.P.; Kabeya, N.; Marrero, M.; Rodríguez, C. Lipid metabolism in Tinca tinca and its n-3 LC-PUFA biosynthesis capacity. Aquaculture 2020, 523, 735147. [Google Scholar] [CrossRef]
- Galindo, A.; Garrido, D.; Monroig, Ó.; Pérez, J.; Betancor, M.; Acosta, N.; Kabeya, N.; Marrero, M.; Bolaños, A.; Rodríguez, C. Polyunsaturated fatty acid metabolism in three fish species with different trophic level. Aquaculture 2021, 530, 735761. [Google Scholar] [CrossRef]
- Park, W.J.; Kothapalli, K.S.; Lawrence, P.; Tyburczy, C.; Brenna, J.T. An alternate pathway to long-chain polyunsaturates: The FADS2 gene product Δ8-desaturates 20:2n-6 and 20:3n-3. J. Lipid Res. 2009, 50, 1195–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heindel, J.J.; Vandenberg, L.N. Developmental origins of health and disease: A paradigm for understanding disease etiology and prevention. Curr. Opin. Pediatr. 2015, 27, 248–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.-Y.; Ramos, K.S.; Chapkin, R.S. Dietary γ-Linolenic Acid Enhances Mouse Macrophage-Derived Prostaglandin E1 Which Inhibits Vascular Smooth Muscle Cell Proliferation. J. Nutr. 1997, 127, 1765–1771. [Google Scholar] [CrossRef] [Green Version]
- Fraher, D.; Sanigorski, A.; Mellett, N.A.; Meikle, P.; Sinclair, A.J.; Gibert, Y. Zebrafish Embryonic Lipidomic Analysis Reveals that the Yolk Cell Is Metabolically Active in Processing Lipid. Cell Rep. 2016, 14, 1317–1329. [Google Scholar] [CrossRef] [Green Version]
- Miyares, R.L.; de Rezende, V.B.; Farber, S.A. Zebrafish yolk lipid processing: A tractable tool for the study of vertebrate lipid transport and metabolism. Dis. Model. Mech. 2014, 7, 915–927. [Google Scholar] [CrossRef] [Green Version]
- Sant, K.; Timme-Laragy, A.R. Zebrafish as a Model for Toxicological Perturbation of Yolk and Nutrition in the Early Embryo. Curr. Environ. Health Rep. 2018, 5, 125–133. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio rerio); University of Oregon Press: Eugene, OR, USA, 2007. [Google Scholar]
- Jao, L.-E.; Wente, S.R.; Chen, W. Efficient multiplex biallelic zebrafish genome editing using a CRISPR nuclease system. Proc. Natl. Acad. Sci. USA 2013, 110, 13904–13909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varshney, G.K.; Pei, W.; LaFave, M.C.; Idol, J.; Xu, L.; Gallardo, V.; Carrington, B.; Bishop, K.; Jones, M.; Li, M.; et al. High-throughput gene targeting and phenotyping in zebrafish using CRISPR/Cas9. Genome Res. 2015, 25, 1030–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirro, V.; Guffey, S.C.; Sepúlveda, M.S.; Mahapatra, C.T.; Ferreira, C.R.; Jarmusch, A.K.; Cooks, R.G. Lipid dynamics in zebrafish embryonic development observed by DESI-MS imaging and nanoelectrospray-MS. Mol. BioSyst. 2016, 12, 2069–2079. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.F.; Tocher, D.R.; Monroig, O. Long-chain polyunsaturated fatty acid biosynthesis in chordates: Insights into the evolution of Fads and Elovl gene repertoire. Prog. Lipid Res. 2016, 62, 25–40. [Google Scholar] [CrossRef]
- Monroig, Ó.; Li, Y.; Tocher, D.R. Delta-8 desaturation activity varies among fatty acyl desaturases of teleost fish: High activity in delta-6 desaturases of marine species. Comp. Biochem. Physiol. 2011, 159, 206–213. [Google Scholar] [CrossRef]
- Hastings, N.; Agaba, M.; Tocher, D.R.; Leaver, M.J.; Dick, J.R.; Sargent, J.R.; Teale, A.J. A vertebrate fatty acid desaturase with Δ5 and Δ6 activities. Proc. Natl. Acad. Sci. USA 2001, 98, 14304–14309. [Google Scholar] [CrossRef] [Green Version]
- Kabir, M.A.; Munir, M.B.; Alias, S.L.F.W.; Leja, A.; Lee, S.W.; Hashim, R. Effect of Different Dietary Lipid Levels on Spawning Performance and Egg Quality of Pangasianodon hypophthalmus (Sauvage, 1878). Asian Fish. Sci. 2019, 32, 138–146. [Google Scholar] [CrossRef]
- Xu, H.; Ferosekhan, S.; Turkmen, S.; Afonso, J.M.; Zamorano, M.J.; Izquierdo, M. Influence of Parental fatty acid desaturase 2 (fads2) Expression and Diet on Gilthead Seabream (Sparus aurata) Offspring fads2 Expression during Ontogenesis. Animals 2020, 10, 2191. [Google Scholar] [CrossRef]
- Oseeva, M.; Paluchova, V.; Zacek, P.; Janovska, P.; Mracek, T.; Rossmeisl, M.; Hamplova, D.; Cadova, N.; Stohanzlova, I.; Flachs, P.; et al. Omega-3 index in the Czech Republic: No difference between urban and rural populations. Chem. Phys. Lipids 2019, 220, 23–27. [Google Scholar] [CrossRef]
- Stoffel, W.; Schmidt-Soltau, I.; Binczek, E.; Thomas, A.; Thevis, M.; Wegner, I. Dietary ω3-and ω6-Polyunsaturated fatty acids reconstitute fertility of Juvenile and adult Fads2-Deficient mice. Mol. Metab. 2020, 36, 100974. [Google Scholar] [CrossRef]
- Zhong, Z.; Niu, P.; Wang, M.; Huang, G.; Xu, S.; Sun, Y.; Xu, X.; Hou, Y.; Sun, X.; Yan, Y.; et al. Targeted disruption of sp7 and myostatin with CRISPR-Cas9 results in severe bone defects and more muscular cells in common carp. Sci. Rep. 2016, 6, 22953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, C.J.; Monstad-Rios, A.T.; Bhimani, R.M.; Gistelinck, C.; Willaert, A.; Coucke, P.; Hsu, Y.-H.; Kwon, R.Y. Phenomics-Based Quantification of CRISPR-Induced Mosaicism in Zebrafish. Cell Syst. 2020, 10, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.synthego.com/guide/how-to-use-crispr/ice-analysis-guide (accessed on 1 April 2022).
- Hsiau, T.; Conant, D.; Rossi, N.; Maures, T.; Waite, K.; Yang, J.; Joshi, S.; Kelso, R.; Holden, K.; Enzmann, B.L.; et al. Inference of CRISPR Edits from Sanger trace data. bioRxiv 2018, 251082. [Google Scholar] [CrossRef]
- Castro, L.F.C.; Monroig, Ó.; Leaver, M.J.; Wilson, J.; Cunha, I.; Tocher, D.R. Functional Desaturase Fads1 (Δ5) and Fads2 (Δ6) Orthologues Evolved before the Origin of Jawed Vertebrates. PLoS ONE 2012, 7, e31950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stroud, C.K.; Nara, T.Y.; Roqueta-Rivera, M.; Radlowski, E.C.; Lawrence, P.; Zhang, Y.; Cho, B.H.; Segre, M.; Hess, R.; Brenna, J.T.; et al. Disruption of FADS2 gene in mice impairs male reproduction and causes dermal and intestinal ulceration. J. Lipid Res. 2009, 50, 1870–1880. [Google Scholar] [CrossRef] [Green Version]
- Sibbons, C.M.; Irvine, N.A.; Pérez-Mojica, J.E.; Calder, P.C.; Lillycrop, K.A.; Fielding, B.A.; Burdge, G.C. Polyunsaturated Fatty Acid Biosynthesis Involving Δ8 Desaturation and Differential DNA Methylation of FADS2 Regulates Proliferation of Human Peripheral Blood Mononuclear Cells. Front. Immunol. 2018, 9, 432. [Google Scholar] [CrossRef] [Green Version]
- Datsomor, A.K.; Olsen, R.E.; Zic, N.; Madaro, A.; Bones, A.M.; Edvardsen, R.B.; Wargelius, A.; Winge, P. CRISPR/Cas9-mediated editing of Δ5 and Δ6 desaturases impairs Δ8-desaturation and docosahexaenoic acid synthesis in Atlantic salmon (Salmo salar L.). Sci. Rep. 2019, 9, 16888. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H-Value (F-Value) | Number of d.f. | p-Value | |
|---|---|---|---|
| C18:3 Δ6,9,132 | 9.99 | 2 | 0.007 |
| C20:2 Δ11,14 | 20.39 | 2 | <0.001 |
| C20:4, arachnidonate | 9.37 | 2 | 0.009 |
| C18:4 Δ6,9,12,15 | 7.56 | 2 | 0.023 |
| C20:3 Δ11,14,17 | 17.50 | 2 | <0.001 |
| C20:5, EPA | 2.12 | 2.27 | 0.139 |
| C22:6, DHA | 11.36 | 2 | 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bláhová, Z.; Franěk, R.; Let, M.; Bláha, M.; Pšenička, M.; Mráz, J. Partial fads2 Gene Knockout Diverts LC-PUFA Biosynthesis via an Alternative Δ8 Pathway with an Impact on the Reproduction of Female Zebrafish (Danio rerio). Genes 2022, 13, 700. https://doi.org/10.3390/genes13040700
Bláhová Z, Franěk R, Let M, Bláha M, Pšenička M, Mráz J. Partial fads2 Gene Knockout Diverts LC-PUFA Biosynthesis via an Alternative Δ8 Pathway with an Impact on the Reproduction of Female Zebrafish (Danio rerio). Genes. 2022; 13(4):700. https://doi.org/10.3390/genes13040700
Chicago/Turabian StyleBláhová, Zuzana, Roman Franěk, Marek Let, Martin Bláha, Martin Pšenička, and Jan Mráz. 2022. "Partial fads2 Gene Knockout Diverts LC-PUFA Biosynthesis via an Alternative Δ8 Pathway with an Impact on the Reproduction of Female Zebrafish (Danio rerio)" Genes 13, no. 4: 700. https://doi.org/10.3390/genes13040700
APA StyleBláhová, Z., Franěk, R., Let, M., Bláha, M., Pšenička, M., & Mráz, J. (2022). Partial fads2 Gene Knockout Diverts LC-PUFA Biosynthesis via an Alternative Δ8 Pathway with an Impact on the Reproduction of Female Zebrafish (Danio rerio). Genes, 13(4), 700. https://doi.org/10.3390/genes13040700