
Independent COL5A1 Variants in Cats with Ehlers-Danlos Syndrome

 ,
,  , , , , ,
, , , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Selection
2.2. Clinical and Histopathological Examinations
2.3. Transmission Electron Microscopy (TEM)
2.4. DNA Extraction
2.5. Whole-Genome Sequencing and Variant Calling
2.6. Gene Analysis
2.7. PCR and Sanger Sequencing
3. Results
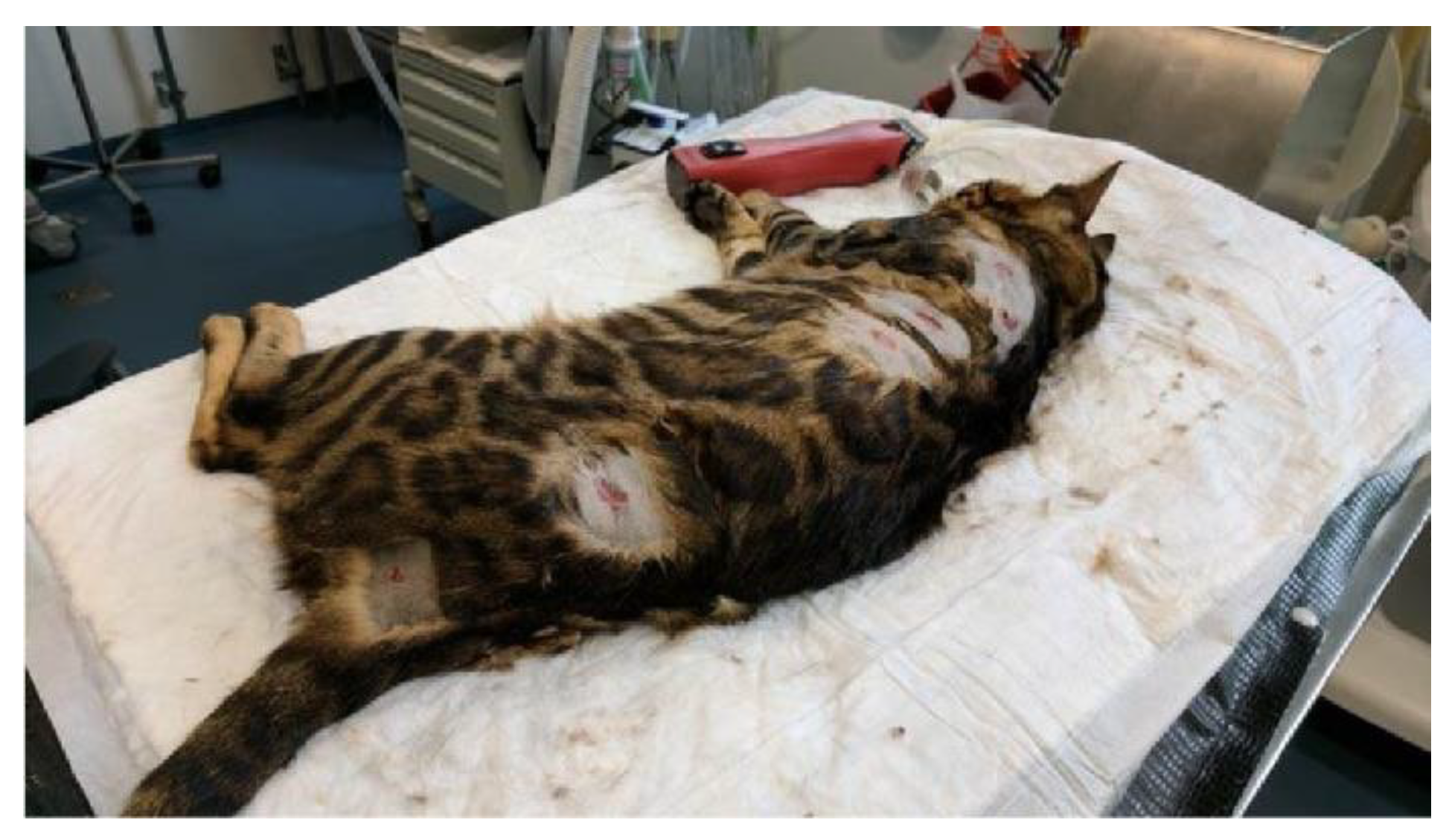
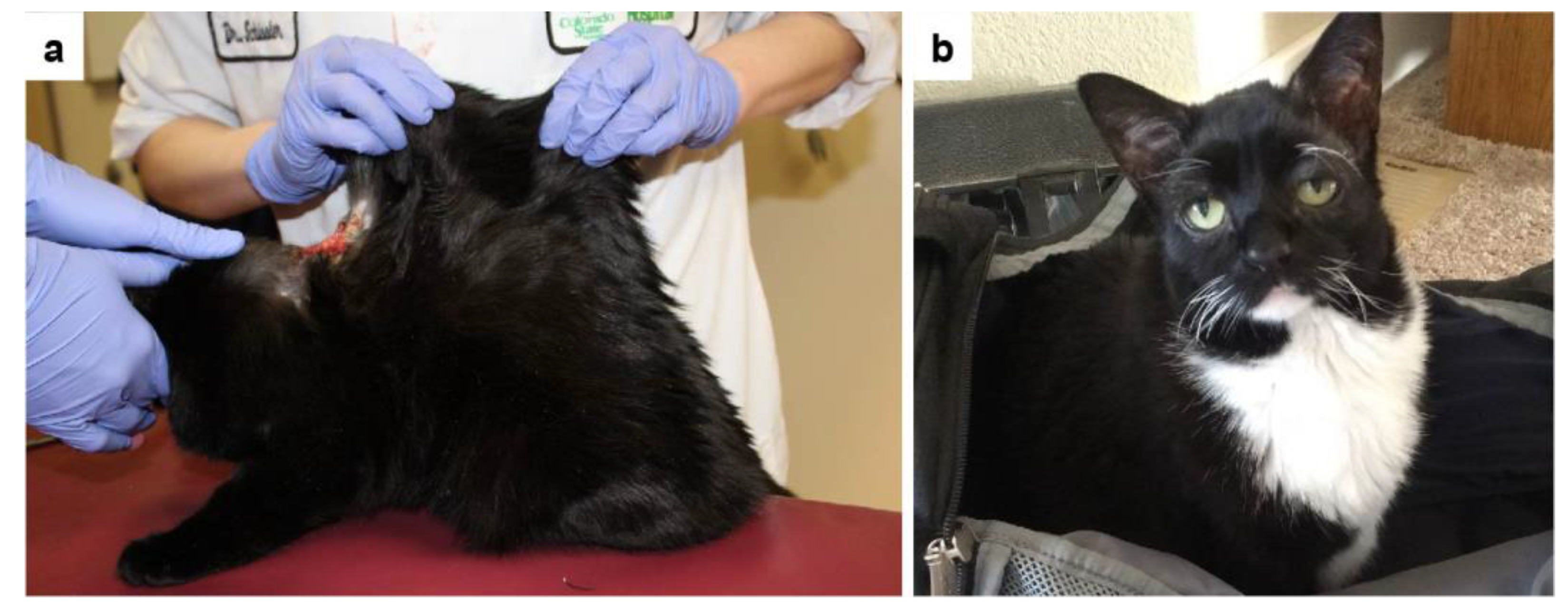
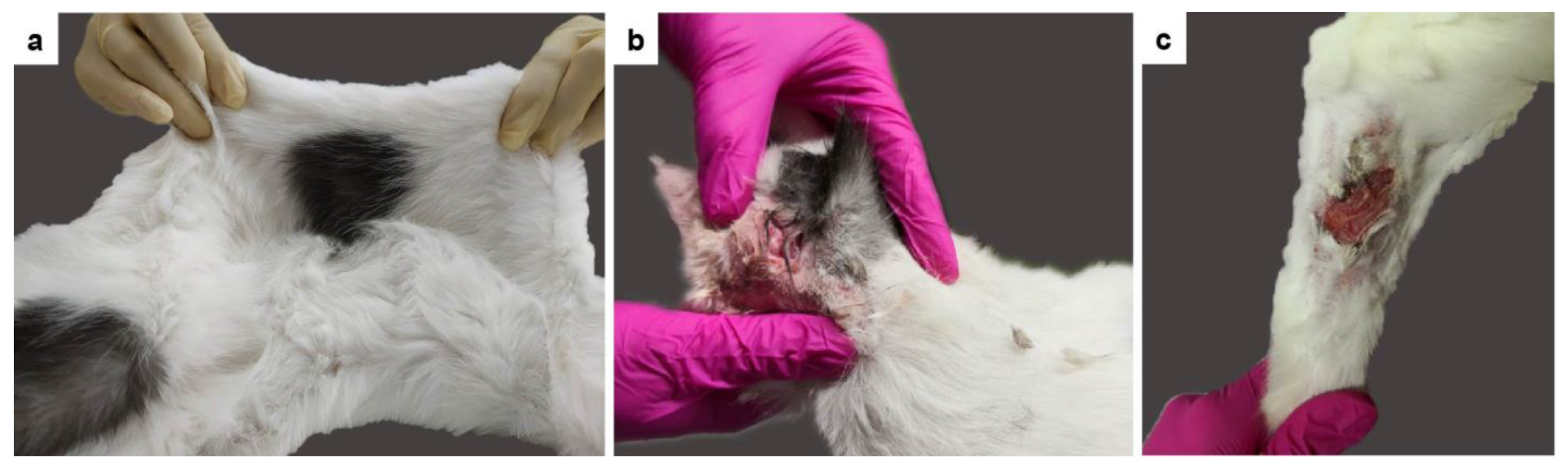
3.1. Clinical History and Examination
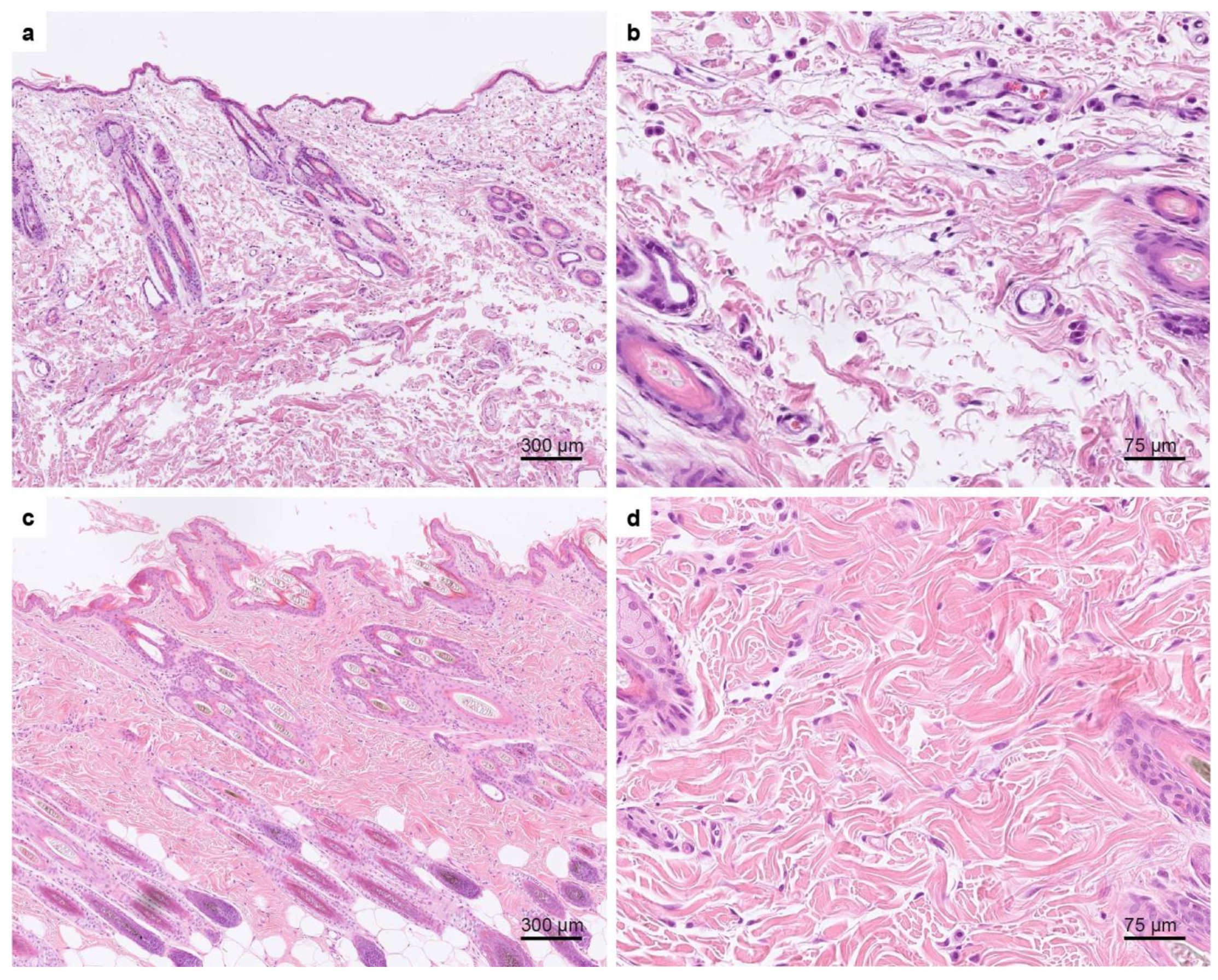
3.2. Histopathological Examination
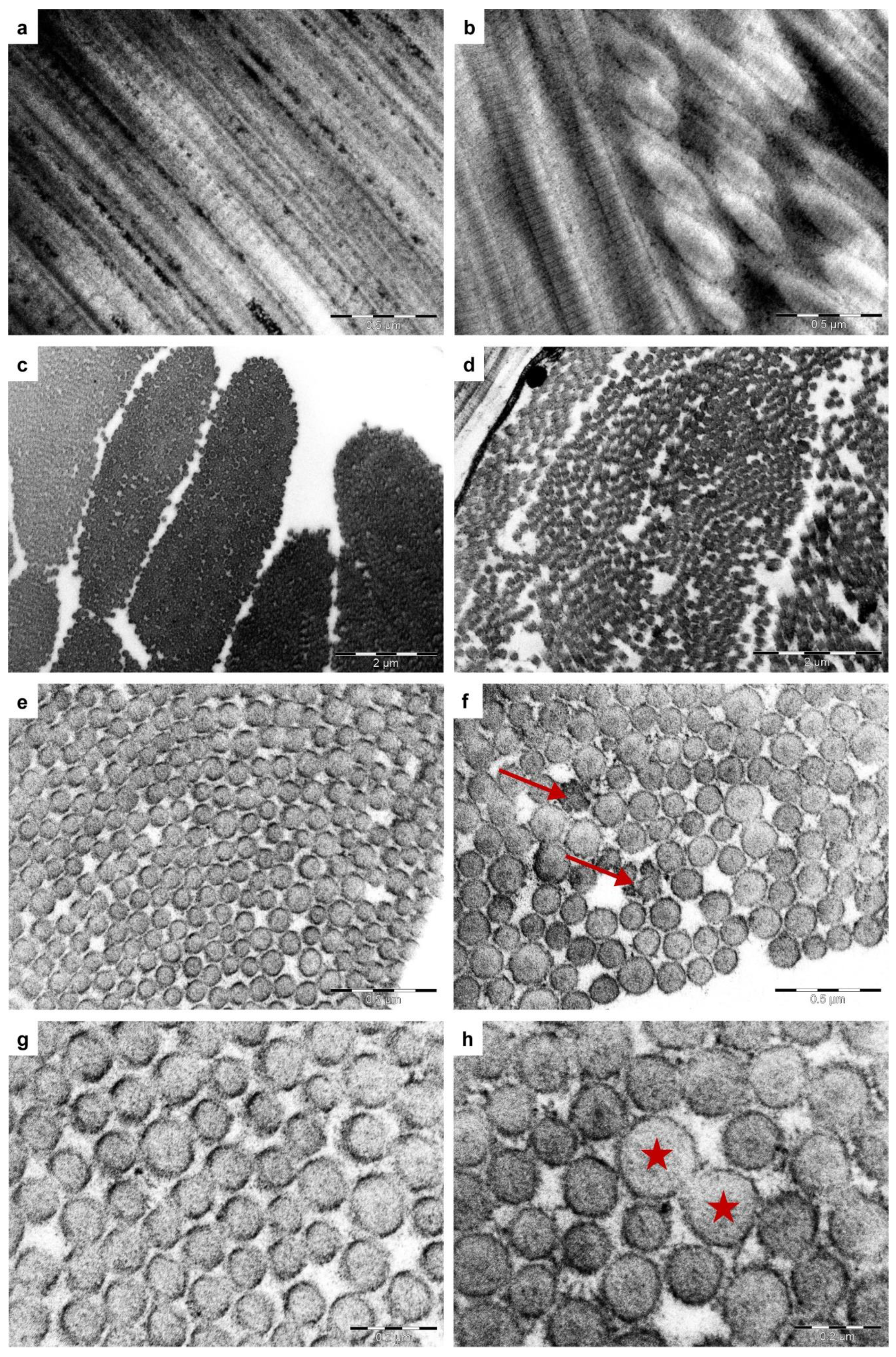
3.3. Ultrastructural Examination
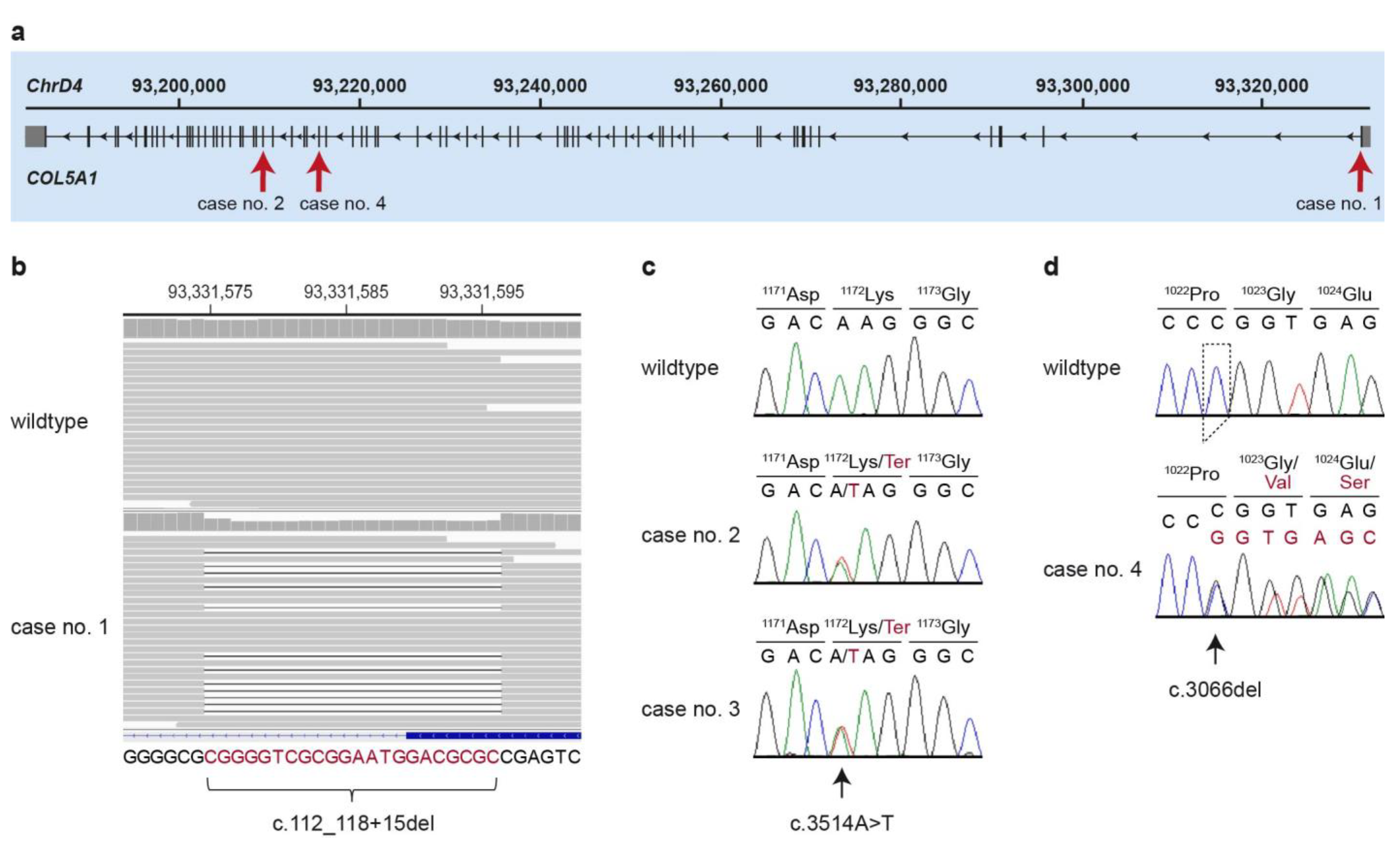
3.4. Genetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malfait, F.; Castori, M.; Francomano, C.A.; Giunta, C.; Kosho, T.; Byers, P.H. The Ehlers–Danlos syndromes. Nat. Rev. Dis. Prim. 2020, 6, 64. [Google Scholar] [CrossRef] [PubMed]
- Malfait, F.; Francomano, C.; Byers, P.; Belmont, J.; Berglund, B.; Black, J.; Bloom, L.; Bowen, J.M.; Brady, A.F.; Burrows, N.P.; et al. The 2017 international classification of the Ehlers-Danlos syndromes. Am. J. Med. Genet. Part C Semin. Med. Genet. 2017, 175, 8–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackburn, P.R.; Xu, Z.; Tumelty, K.E.; Zhao, R.W.; Monis, W.J.; Harris, K.G.; Gass, J.M.; Cousin, M.A.; Boczek, N.J.; Mitkov, M.V.; et al. Bi-allelic Alterations in AEBP1 Lead to Defective Collagen Assembly and Connective Tissue Structure Resulting in a Variant of Ehlers-Danlos Syndrome. Am. J. Hum. Genet. 2018, 102, 696–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholls, A.C.; Oliver, J.E.; McCarron, S.; Harrison, J.B.; Greenspan, D.S.; Pope, F.M. An exon skipping mutation of a type V collagen gene (COL5A1) in Ehlers-Danlos syndrome. J. Med. Genet. 1996, 33, 940–946. [Google Scholar] [CrossRef] [Green Version]
- Wenstrup, R. A splice-junction mutation in the region of COL5A1 that codes for the carboxyl propeptide of pro alpha 1(V) chains results in the gravis form of the Ehlers-Danlos syndrome (type I). Hum. Mol. Genet. 1996, 5, 1733–1736. [Google Scholar] [CrossRef] [Green Version]
- De Paepe’, A.; Nuytinck, L.; Hausser, I.; Anton-Lamprecht, I.; Naeyaert2, J.-M. Mutations in the COL5A1 Gene Are Causal in the Ehlers-Danlos Syndromes I and II. Am. J. Hum. Genet 1997, 60, 547–554. [Google Scholar]
- Richards, A.J.; Martin, S.; Nicholls, A.C.; Harrison, J.B.; Pope, F.M.; Burrows, N.P. A single base mutation in COL5A2 causes Ehlers-Danlos syndrome type II. J. Med. Genet. 1998, 35, 846–848. [Google Scholar] [CrossRef] [Green Version]
- Bouma, P.; Cabral, W.A.; Cole, W.G.; Marini, J.C. COL5A1 Exon 14 Splice Acceptor Mutation Causes a Functional Null Allele, Haploinsufficiency of α1(V) and Abnormal Heterotypic Interstitial Fibrils in Ehlers-Danlos Syndrome II. J. Biol. Chem. 2001, 276, 13356–13364. [Google Scholar] [CrossRef] [Green Version]
- Symoens, S.; Syx, D.; Malfait, F.; Callewaert, B.; De Backer, J.; Vanakker, O.; Coucke, P.; De Paepe, A. Comprehensive molecular analysis demonstrates type V collagen mutations in over 90% of patients with classic EDS and allows to refine diagnostic criteria. Hum. Mutat. 2012, 33, 1485–1493. [Google Scholar] [CrossRef]
- Vroman, R.; Malfait, A.-M.; Miller, R.E.; Malfait, F.; Syx, D. Animal Models of Ehlers–Danlos Syndromes: Phenotype, Pathogenesis, and Translational Potential. Front. Genet. 2021, 12, 1610. [Google Scholar] [CrossRef]
- Scott, D.V. Cutaneous asthenia in a cat, resembling Ehlers-Danlos syndrome in man. Vet. Med. Small Anim. Clin. 1974, 69, 1256–1258. [Google Scholar] [PubMed]
- Smids, M.Y.L. Experience with a cat with cutis asthenia (Ehlers-Danlos syndrome). Tijdschr. Diergeneeskd. 2008, 133, 612–614. [Google Scholar] [PubMed]
- Halper, J. Connective Tissue Disorders in Domestic Animals. In Progress in Heritable Soft Connective Tissue Diseases; Springer: Dordrecht, The Netherlands, 2014; pp. 231–240. [Google Scholar]
- Hansen, N.; Foster, S.F.; Burrows, A.K.; Mackie, J.; Malik, R. Cutaneous asthenia (Ehlers–Danlos-like syndrome) of Burmese cats. J. Feline Med. Surg. 2015, 17, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.H.; Halper, J. Connective Tissue Disorders in Domestic Animals. In Advances in Experimental Medicine and Biology; Springer: Dordrecht, The Netherlands, 2021; Volume 1348, pp. 325–335. [Google Scholar]
- Patterson, D.F.; Minor, R.R. Hereditary fragility and hyperextensibility of the skin of cats. A defect in collagen fibrillogenesis. Lab. Investig. 1977, 37, 170–179. [Google Scholar]
- Counts, D.F.; Byers, P.H.; Holbrook, K.A.; Hegreberg, G.A. Dermatosparaxis in a Himalayan cat: I. Biochemical studies of dermal collagen. J. Investig. Dermatol. 1980, 74, 96–99. [Google Scholar] [CrossRef] [Green Version]
- Holbrook, K.A.; Byers, P.H.; Counts, D.F.; Hegreberg, G.A. Dermatosparaxis in a Himalayan cat: II. Ultrastructural studies of dermal collagen. J. Investig. Dermatol. 1980, 74, 100–104. [Google Scholar] [CrossRef] [Green Version]
- Verweij, C.G.; van Zuylen, A.L. Cutaneous asthenia, a congenital skin disease in a Burmese cat. Tijdschr. Diergeneeskd. 1986, 111, 244–246. [Google Scholar]
- Freeman, L.J.; Hegreberg, G.A.; Robinette, J.D. Ehlers-Danlos syndrome in dogs and cats. Semin. Vet. Med. Surg. (Small Anim.) 1987, 2, 221–227. [Google Scholar]
- Plotnick, A.; Brunt, J.E.; Reitz, B. Cutaneous Asthenia in a Cat. Feline Pract. 1992, 20, 9–12. [Google Scholar]
- Sequeira, J.L.; Rocha, N.S.; Bandarra, E.P.; Figueiredo, L.M.; Eugenio, F.R. Collagen dysplasia (cutaneous asthenia) in a cat. Vet. Pathol. 1999, 36, 603–606. [Google Scholar] [CrossRef] [Green Version]
- Benitah, N.; Matousek, J.L.; Barnes, R.F.; Lichtensteiger, C.A.; Campbell, K.L. Diaphragmatic and perineal hernias associated with cutaneous asthenia in a cat. J. Am. Vet. Med. Assoc. 2004, 224, 706–709, 698. [Google Scholar] [CrossRef] [PubMed]
- Spycher, M.; Bauer, A.; Jagannathan, V.; Frizzi, M.; De Lucia, M.; Leeb, T. A frameshift variant in the COL5A1 gene in a cat with Ehlers-Danlos syndrome. Anim. Genet. 2018, 49, 641–644. [Google Scholar] [CrossRef] [PubMed]
- Jagannathan, V.; Drögemüller, C.; Leeb, T.; Aguirre, G.; André, C.; Bannasch, D.; Becker, D.; Davis, B.; Ekenstedt, K.; Faller, K.; et al. A comprehensive biomedical variant catalogue based on whole genome sequences of 582 dogs and eight wolves. Anim. Genet. 2019, 50, 695–704. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Titeux, E.; Gilbert, C.; Briand, A.; Cochet-Faivre, N. From Feline Idiopathic Ulcerative Dermatitis to Feline Behavioral Ulcerative Dermatitis: Grooming Repetitive Behaviors Indicators of Poor Welfare in Cats. Front. Vet. Sci. 2018, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favrot, C.; Steffan, J.; Seewald, W.; Hobi, S.; Linek, M.; Marignac, G.; Olivry, T.; Beco, L.; Nett, C.; Fontaine, J.; et al. Establishment of diagnostic criteria for feline nonflea-induced hypersensitivity dermatitis. Vet. Dermatol. 2012, 23, 45-e11. [Google Scholar] [CrossRef]
- Santoro, D.; Pucheu-Haston, C.M.; Prost, C.; Mueller, R.S.; Jackson, H. Clinical signs and diagnosis of feline atopic syndrome: Detailed guidelines for a correct diagnosis. Vet. Dermatol. 2021, 32, 26-e6. [Google Scholar] [CrossRef]
- Angwin, C.; Ghali, N.; Baker, D.; Brady, A.F.; Pope, F.M.; Vandersteen, A.; Wagner, B.; Ferguson, D.J.P.; Dijk, F. Electron microscopy in the diagnosis of Ehlers–Danlos syndromes: Correlation with clinical and genetic investigations. Br. J. Dermatol. 2020, 182, 698–707. [Google Scholar] [CrossRef]
- Uri, M.; Verin, R.; Ressel, L.; Buckley, L.; McEwan, N. Ehlers–Danlos Syndrome Associated with Fatal Spontaneous Vascular Rupture in a Dog. J. Comp. Pathol. 2015, 152, 211–216. [Google Scholar] [CrossRef]
- Weingart, C.; Haußer, I.; Kershaw, O.; Kohn, B. Ehlers-Danlos-like-Syndrom bei einer Katze. Schweiz. Arch. Tierheilkd. 2014, 156, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.; Bateman, J.F.; Lamandé, S.R.; Hanssen, E.; Kirejczyk, S.G.M.; Yee, M.; Ramiche, A.; Jagannathan, V.; Welle, M.; Leeb, T.; et al. Identification of Two Independent COL5A1 Variants in Dogs with Ehlers–Danlos Syndrome. Genes 2019, 10, 731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birk, D.E. Type V collagen: Heterotypic type I/V collagen interactions in the regulation of fibril assembly. Micron 2001, 32, 223–237. [Google Scholar] [CrossRef]
- Schwarze, U.; Atkinson, M.; Hoffman, G.G.; Greenspan, D.S.; Byers, P.H. Null Alleles of the COL5A1 Gene of Type V Collagen Are a Cause of the Classical Forms of Ehlers-Danlos Syndrome (Types I and II). Am. J. Hum. Genet. 2000, 66, 1757–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenstrup, R.J.; Florer, J.B.; Willing, M.C.; Giunta, C.; Steinmann, B.; Young, F.; Susic, M.; Cole, W.G. COL5A1 Haploinsufficiency Is a Common Molecular Mechanism Underlying the Classical Form of EDS. Am. J. Hum. Genet. 2000, 66, 1766–1776. [Google Scholar] [CrossRef] [Green Version]
- Wenstrup, R.J.; Florer, J.B.; Davidson, J.M.; Phillips, C.L.; Pfeiffer, B.J.; Menezes, D.W.; Chervoneva, I.; Birk, D.E. Murine model of the Ehlers-Danlos syndrome. col5a1 haploinsufficiency disrupts collagen fibril assembly at multiple stages. J. Biol. Chem. 2006, 281, 12888–12895. [Google Scholar] [CrossRef] [Green Version]
- Savasta, S.; Merli, P.; Ruggieri, M.; Bianchi, L.; Spartà, M.V. Ehlers–Danlos syndrome and neurological features: A review. Child’s Nerv. Syst. 2011, 27, 365–371. [Google Scholar] [CrossRef]
- Cortini, F.; Villa, C. Ehlers-Danlos syndromes and epilepsy: An updated review. Seizure 2018, 57, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Cazzato, D.; Castori, M.; Lombardi, R.; Caravello, F.; Bella, E.D.; Petrucci, A.; Grammatico, P.; Dordoni, C.; Colombi, M.; Lauria, G. Small fiber neuropathy is a common feature of Ehlers-Danlos syndromes. Neurology 2016, 87, 155–159. [Google Scholar] [CrossRef] [Green Version]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–423. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Filtering Step | Variants Case No. 1 | Variants Case No. 2 | Variants Case No. 4 | |||
|---|---|---|---|---|---|---|
| hom | het | hom | het | hom | het | |
| all variants | 5,758,034 | 9,091,644 | 4,612,879 | 8,593,879 | 4,949,250 | 8,310,846 |
| private variants | 44,007 | 938,018 | 6242 | 194,315 | 11,606 | 168,540 |
| protein-changing private variants | 91 | 2901 | 15 | 880 | 51 | 775 |
| in 20 known EDS candidate genes | 0 | 1 | 0 | 1 | 0 | 3 |
| Cats | HGVS Variant Designations | ||
|---|---|---|---|
| Genomic (felCat9.0) | mRNA (XM_023242950.1) | Protein (XP_023098718.1) | |
| case no. 1 | ChrD4:93,331,577_93,331,598del | c.112_118+15del | r.spl? |
| cases no. 2 and 3 | ChrD4:93,209,345T>A | c.3514A>T | p.(Lys1172*) |
| case no. 4 | ChrD4:93,215,496del | c.3066del | p.(Gly1023Valfs*50) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiener, S.; Apostolopoulos, N.; Schissler, J.; Hass, P.-K.; Leuthard, F.; Jagannathan, V.; Schuppisser, C.; Soto, S.; Welle, M.; Mayer, U.; et al. Independent COL5A1 Variants in Cats with Ehlers-Danlos Syndrome. Genes 2022, 13, 797. https://doi.org/10.3390/genes13050797
Kiener S, Apostolopoulos N, Schissler J, Hass P-K, Leuthard F, Jagannathan V, Schuppisser C, Soto S, Welle M, Mayer U, et al. Independent COL5A1 Variants in Cats with Ehlers-Danlos Syndrome. Genes. 2022; 13(5):797. https://doi.org/10.3390/genes13050797
Chicago/Turabian StyleKiener, Sarah, Neoklis Apostolopoulos, Jennifer Schissler, Pascal-Kolja Hass, Fabienne Leuthard, Vidhya Jagannathan, Carole Schuppisser, Sara Soto, Monika Welle, Ursula Mayer, and et al. 2022. "Independent COL5A1 Variants in Cats with Ehlers-Danlos Syndrome" Genes 13, no. 5: 797. https://doi.org/10.3390/genes13050797
APA StyleKiener, S., Apostolopoulos, N., Schissler, J., Hass, P. -K., Leuthard, F., Jagannathan, V., Schuppisser, C., Soto, S., Welle, M., Mayer, U., Leeb, T., Fischer, N. M., & Kaessmeyer, S. (2022). Independent COL5A1 Variants in Cats with Ehlers-Danlos Syndrome. Genes, 13(5), 797. https://doi.org/10.3390/genes13050797