
Cdh1 Deficiency Sensitizes TNBC Cells to PARP Inhibitors

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Western Blot Analysis
2.3. Assays for Cell Proliferation
2.4. Plasmids and Lentiviruses
2.5. Antibodies
2.6. Immunostaining
2.7. Ultrafine Anaphase Bridges Detection
2.8. Apoptosis Assay
2.9. Cell Cycle Analysis
2.10. Clinical Samples and Data Acquisition
2.11. Analysis of Differentially Expressed Genes between TNBC and Normal Tissue Samples
2.12. Statistical Analysis
3. Results
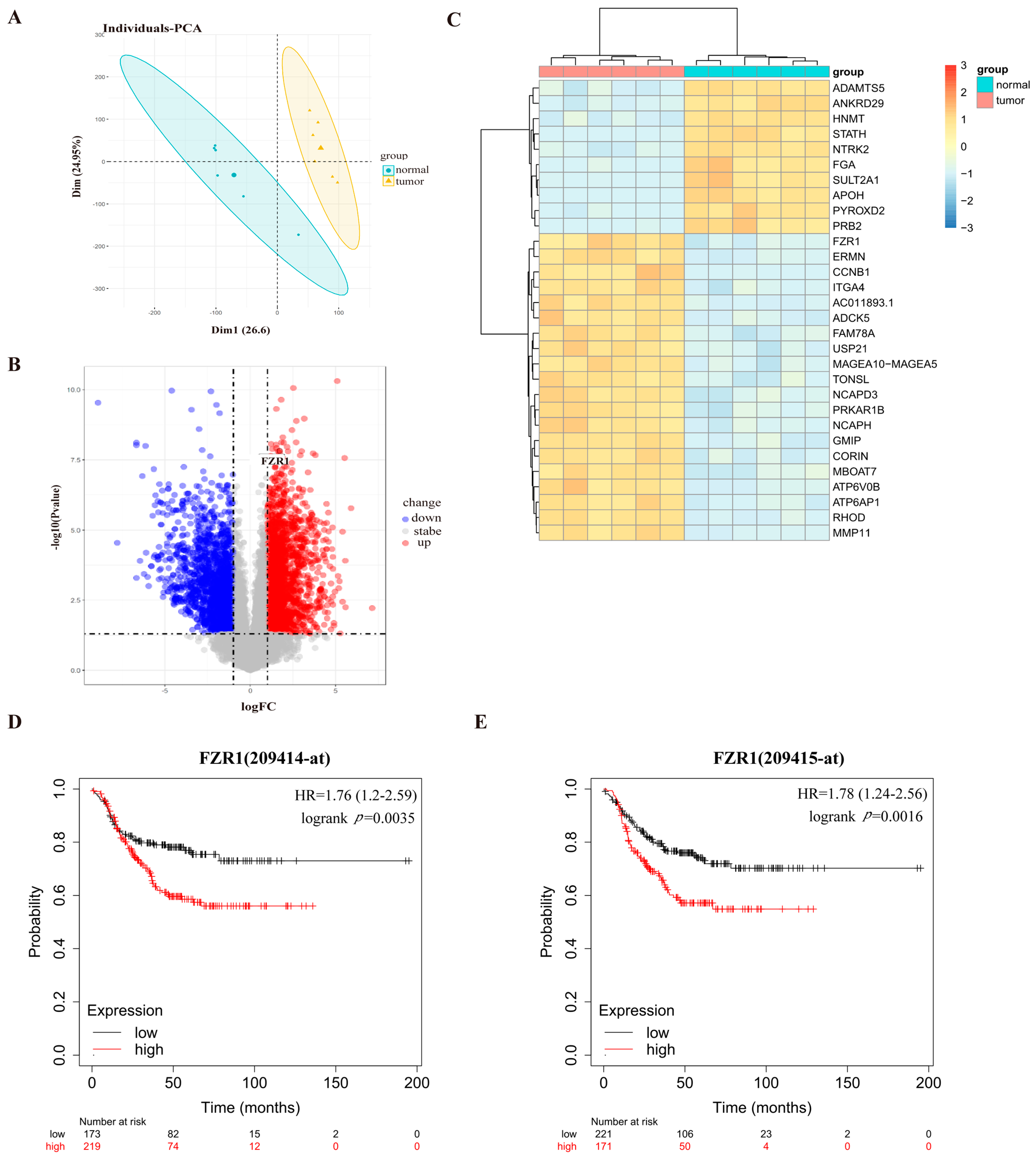
3.1. Cdh1 Expression Correlates with Poor Prognosis in TNBC
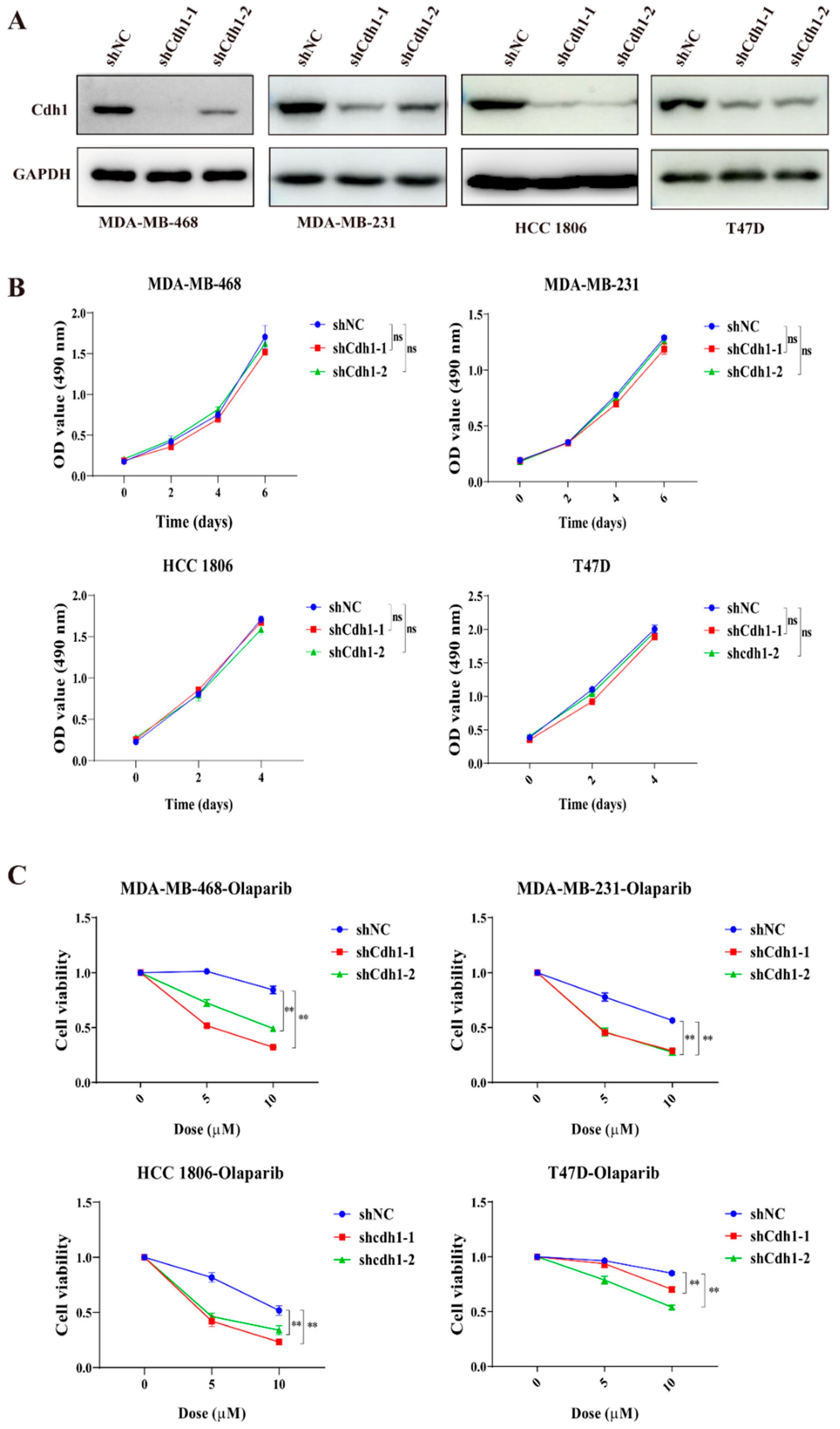
3.2. Cdh1-Deficient TNBC Cells Are Sensitive to the PARP Inhibitor Olaparib
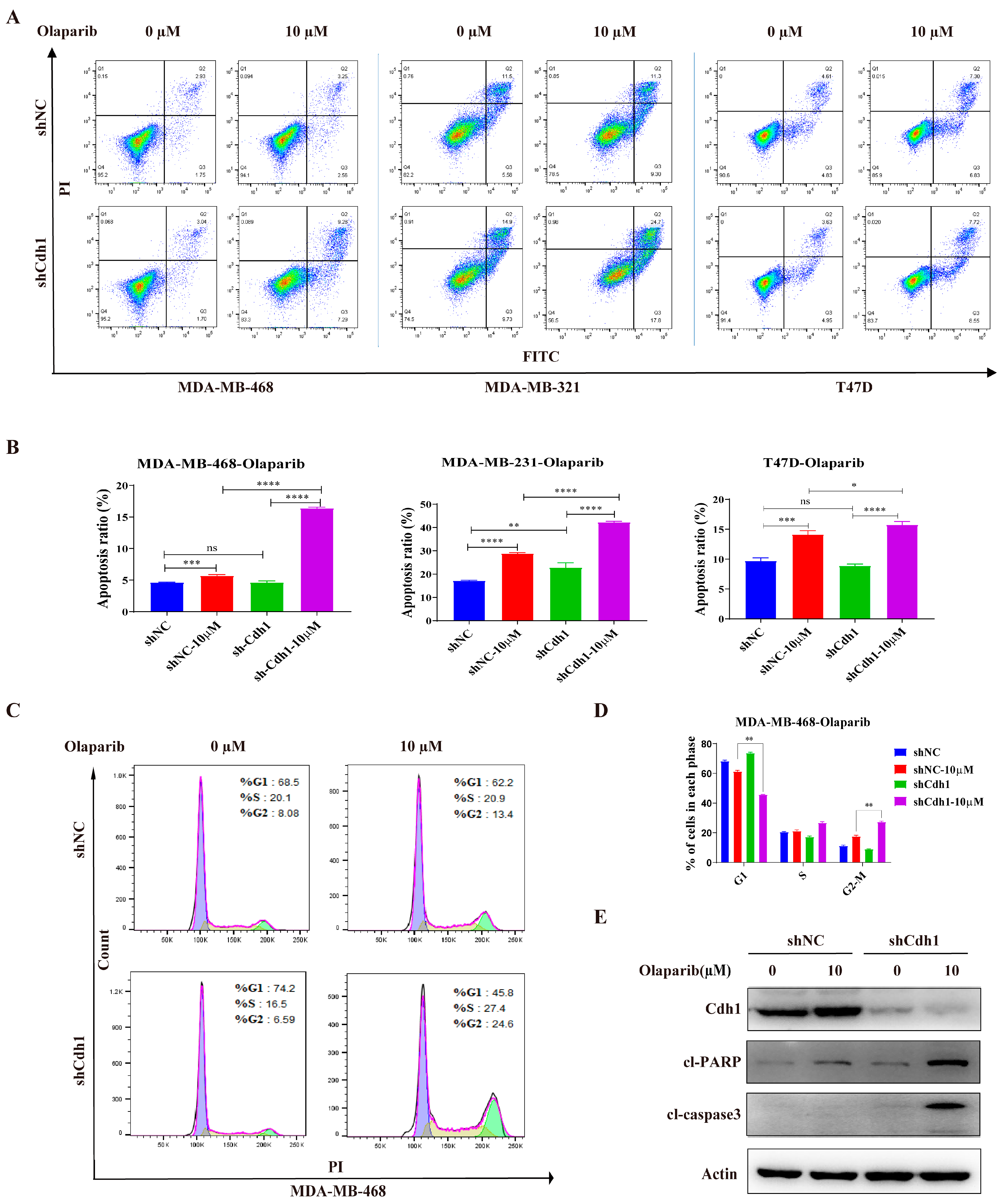
3.3. PARP Inhibitor Induces Elevated Levels of Apoptosis and G2-M Cell Cycle Arrest in Cdh1 Depleted TNBC Cells
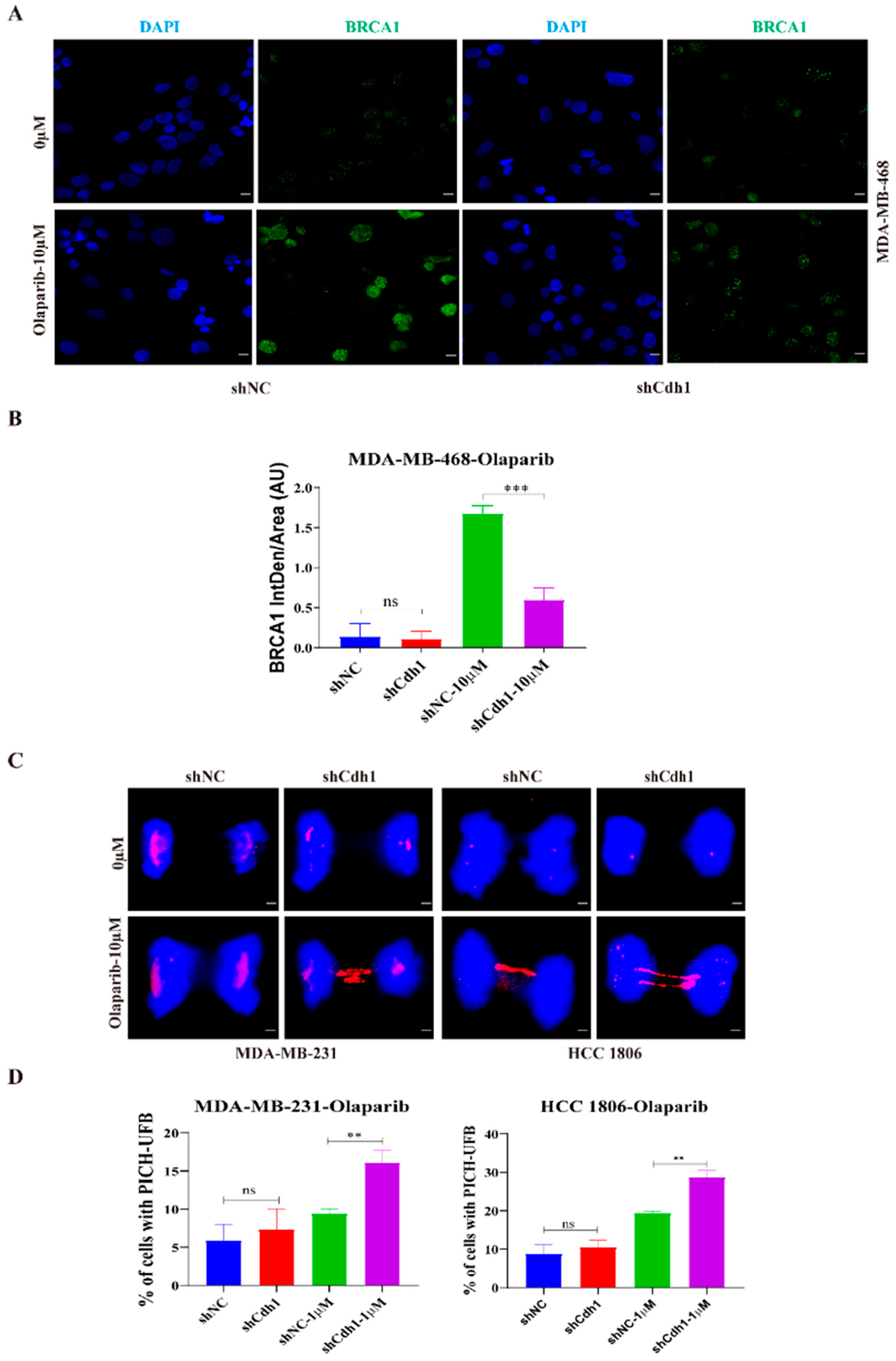
3.4. PARPi Causes DNA Damage in Cdh1 Depleted TNBC Cells
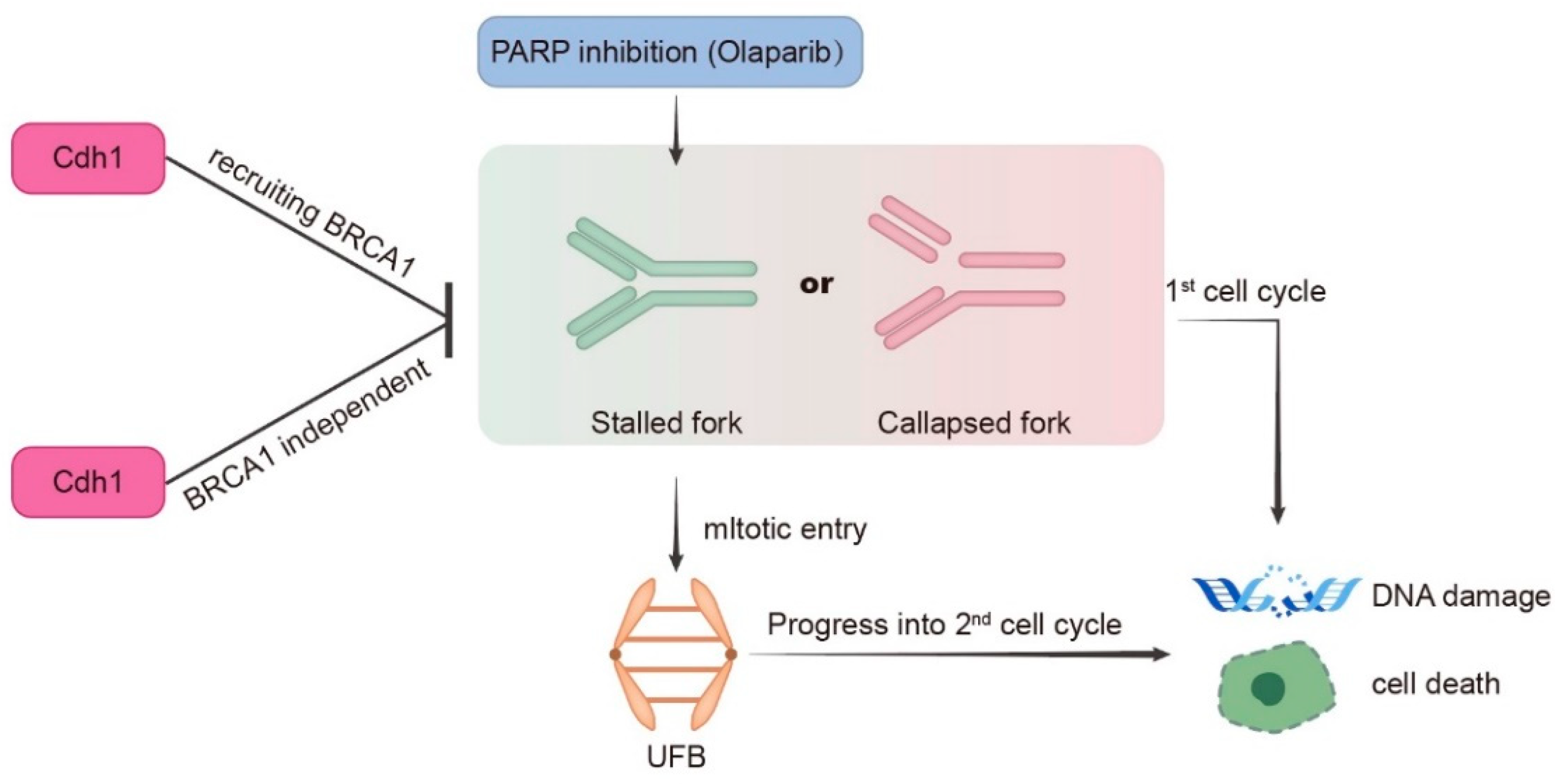
3.5. Cdh1 Promotes BRCA1 Foci Formation and Prevents Untangled DNA Entering Mitosis in Response to PARPi in TNBC Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Liu, C.; Yao, J.; Wan, H.; Wan, G.; Li, Y.; Chen, N. Breast cancer stem cells, heterogeneity, targeting therapies and therapeutic implications. Pharmacol. Res. 2021, 163, 105320. [Google Scholar] [CrossRef] [PubMed]
- Mehraj, U.; Dar, A.H.; Wani, N.A.; Mir, M.A. Tumor microenvironment promotes breast cancer chemoresistance. Cancer Chemother. Pharmacol. 2021, 87, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Onitilo, A.A.; Engel, J.M.; Greenlee, R.T.; Mukesh, B.N. Breast cancer subtypes based on ER/PR and Her2 expression: Comparison of clinicopathologic features and survival. Clin. Med. Res. 2009, 7, 4–13. [Google Scholar] [CrossRef]
- Wysocki, P.J.; Korski, K.; Lamperska, K.; Zaluski, J.; Mackiewicz, A. Primary resistance to docetaxel-based chemotherapy in metastatic breast cancer patients correlates with a high frequency of BRCA1 mutations. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2008, 14, Sc7–Sc10. [Google Scholar] [CrossRef]
- Ray Chaudhuri, A.; Nussenzweig, A. The multifaceted roles of PARP1 in DNA repair and chromatin remodelling. Nat. Rev. Mol. Cell Biol. 2017, 18, 610–621. [Google Scholar] [CrossRef]
- Banerjee, S.; Kaye, S. PARP inhibitors in BRCA gene-mutated ovarian cancer and beyond. Curr. Oncol. Rep. 2011, 13, 442–449. [Google Scholar] [CrossRef]
- Turner, N.; Tutt, A.; Ashworth, A. Hallmarks of ‘BRCAness’ in sporadic cancers. Nat. Rev. Cancer 2004, 4, 814–819. [Google Scholar] [CrossRef]
- Yamano, H. APC/C: Current understanding and future perspectives. F1000Research 2019, 8, 725. [Google Scholar] [CrossRef]
- Benanti, J.A. Coordination of cell growth and division by the ubiquitin-proteasome system. Semin. Cell Dev. Biol. 2012, 23, 492–498. [Google Scholar] [CrossRef] [Green Version]
- Rape, M.; Kirschner, M.W. Autonomous regulation of the anaphase-promoting complex couples mitosis to S-phase entry. Nature 2004, 432, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Ha, K.; Ma, C.; Lin, H.; Tang, L.; Lian, Z.; Zhao, F.; Li, J.M.; Zhen, B.; Pei, H.; Han, S.; et al. The anaphase promoting complex impacts repair choice by protecting ubiquitin signalling at DNA damage sites. Nat. Commun. 2017, 8, 15751. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Watanabe, K.; Mistrik, M.; Vesela, E.; Protivankova, I.; Mailand, N.; Lee, M.; Masai, H.; Lukas, J.; Bartek, J. ATR-Chk1-APC/CCdh1-dependent stabilization of Cdc7-ASK (Dbf4) kinase is required for DNA lesion bypass under replication stress. Genes Dev. 2013, 27, 2459–2472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ercilla, A.; Llopis, A.; Feu, S.; Aranda, S.; Ernfors, P.; Freire, R.; Agell, N. New origin firing is inhibited by APC/CCdh1 activation in S-phase after severe replication stress. Nucleic Acids Res. 2016, 44, 4745–4762. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Smith, L.E.H. Notice of Withdrawal: Retinal Vasculature in Development and Diseases. Annu. Rev. Vis. Sci. 2020, 73–96. [Google Scholar] [CrossRef]
- Toledo, L.I.; Altmeyer, M.; Rask, M.B.; Lukas, C.; Larsen, D.H.; Povlsen, L.K.; Bekker-Jensen, S.; Mailand, N.; Bartek, J.; Lukas, J. ATR prohibits replication catastrophe by preventing global exhaustion of RPA. Cell 2013, 155, 1088–1103. [Google Scholar] [CrossRef] [Green Version]
- Bianco, J.N.; Bergoglio, V.; Lin, Y.L.; Pillaire, M.J.; Schmitz, A.L.; Gilhodes, J.; Lusque, A.; Mazières, J.; Lacroix-Triki, M.; Roumeliotis, T.I.; et al. Overexpression of Claspin and Timeless protects cancer cells from replication stress in a checkpoint-independent manner. Nat. Commun. 2019, 10, 910. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Swami, U.; Preet, R.; Zhang, J. Harnessing DNA Replication Stress for Novel Cancer Therapy. Genes 2020, 11, 990. [Google Scholar] [CrossRef]
- Guerrero Llobet, S.; van der Vegt, B.; Jongeneel, E.; Bense, R.D.; Zwager, M.C.; Schröder, C.P.; Everts, M.; Fehrmann, R.S.N.; de Bock, G.H.; van Vugt, M. Cyclin E expression is associated with high levels of replication stress in triple-negative breast cancer. NPJ Breast Cancer 2020, 6, 40. [Google Scholar] [CrossRef]
- Wang, J.; Dong, Y.; Ma, H.; Wu, L.; Zhen, X.; Tang, L.; Jin, J.; Han, S.; Zhang, P.; Peng, J. The deubiquitinase USP28 stabilizes the expression of RecQ family helicases and maintains the viability of triple negative breast cancer cells. J. Biol. Chem. 2022, 298, 101443. [Google Scholar] [CrossRef]
- Bizard, A.H.; Nielsen, C.F.; Hickson, I.D. Detection of Ultrafine Anaphase Bridges. Methods Mol. Biol. (Clifton N.J.) 2018, 1672, 495–508. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Chen, Y.; Tang, X.; Wei, D.; Xu, X.; Yan, F. Long Noncoding RNA DCST1-AS1 Promotes Cell Proliferation and Metastasis in Triple-negative Breast Cancer by Forming a Positive Regulatory Loop with miR-873-5p and MYC. J. Cancer 2020, 11, 311–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Shin, Y.H.; Hou, L.; Huang, X.; Wei, Z.; Klann, E.; Zhang, P. The adaptor protein of the anaphase promoting complex Cdh1 is essential in maintaining replicative lifespan and in learning and memory. Nat. Cell Biol. 2008, 10, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Ha, K.; Kim, M.S.; Noh, Y.W.; Lin, H.; Tang, L.; Zhu, Q.; Zhang, D.; Chen, H.; Han, S.; et al. The anaphase promoting complex promotes NHEJ repair through stabilizing Ku80 at DNA damage sites. Cell Cycle (Georget. Tex.) 2018, 17, 1138–1145. [Google Scholar] [CrossRef] [Green Version]
- Schoonen, P.M.; Talens, F.; Stok, C.; Gogola, E.; Heijink, A.M.; Bouwman, P.; Foijer, F.; Tarsounas, M.; Blatter, S.; Jonkers, J.; et al. Progression through mitosis promotes PARP inhibitor-induced cytotoxicity in homologous recombination-deficient cancer cells. Nat. Commun. 2017, 8, 15981. [Google Scholar] [CrossRef]
- Liu, Y.; Nielsen, C.F.; Yao, Q.; Hickson, I.D. The origins and processing of ultra fine anaphase DNA bridges. Curr. Opin. Genet. Dev. 2014, 26, 1–5. [Google Scholar] [CrossRef]
- Foulkes, W.D.; Smith, I.E.; Reis-Filho, J.S. Triple-negative breast cancer. N. Engl. J. Med. 2010, 363, 1938–1948. [Google Scholar] [CrossRef] [Green Version]
- Robson, M.; Im, S.A.; Senkus, E.; Xu, B.; Domchek, S.M.; Masuda, N.; Delaloge, S.; Li, W.; Tung, N.; Armstrong, A.; et al. Olaparib for Metastatic Breast Cancer in Patients with a Germline BRCA Mutation. N. Engl. J. Med. 2017, 377, 523–533. [Google Scholar] [CrossRef]
- Robson, M.E.; Tung, N.; Conte, P.; Im, S.A.; Senkus, E.; Xu, B.; Masuda, N.; Delaloge, S.; Li, W.; Armstrong, A.; et al. OlympiAD final overall survival and tolerability results: Olaparib versus chemotherapy treatment of physician’s choice in patients with a germline BRCA mutation and HER2-negative metastatic breast cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2019, 30, 558–566. [Google Scholar] [CrossRef]
- Medina, M.A.; Oza, G.; Sharma, A.; Arriaga, L.G.; Hernández Hernández, J.M.; Rotello, V.M.; Ramirez, J.T. Triple-Negative Breast Cancer: A Review of Conventional and Advanced Therapeutic Strategies. Int. J. Environ. Res. Public Health 2020, 17, 2078. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| shRNA | Sequence |
|---|---|
| shNC | 5′-TTCTCCGAACGTGTCACGT-3′ |
| shCdh1-1 | 5′-TGAGAAGTCTCCCAGTCAG-3′ |
| shCdh1-2 | 5′-GGATTAACGAGAATGAGAA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Lan, M.; Peng, J.; Xiong, Q.; Xu, Y.; Yang, Y.; Zhou, Y.; Liu, J.; Zeng, Z.; Yang, X.; et al. Cdh1 Deficiency Sensitizes TNBC Cells to PARP Inhibitors. Genes 2022, 13, 803. https://doi.org/10.3390/genes13050803
Li J, Lan M, Peng J, Xiong Q, Xu Y, Yang Y, Zhou Y, Liu J, Zeng Z, Yang X, et al. Cdh1 Deficiency Sensitizes TNBC Cells to PARP Inhibitors. Genes. 2022; 13(5):803. https://doi.org/10.3390/genes13050803
Chicago/Turabian StyleLi, Junjun, Mengjiao Lan, Jin Peng, Qunli Xiong, Yongfeng Xu, Yang Yang, Ying Zhou, Jinlu Liu, Zhu Zeng, Xiaojuan Yang, and et al. 2022. "Cdh1 Deficiency Sensitizes TNBC Cells to PARP Inhibitors" Genes 13, no. 5: 803. https://doi.org/10.3390/genes13050803
APA StyleLi, J., Lan, M., Peng, J., Xiong, Q., Xu, Y., Yang, Y., Zhou, Y., Liu, J., Zeng, Z., Yang, X., Zhang, Z., Zhang, P., Zhu, Q., & Wu, W. (2022). Cdh1 Deficiency Sensitizes TNBC Cells to PARP Inhibitors. Genes, 13(5), 803. https://doi.org/10.3390/genes13050803