
Complete Mitochondrial Genome of Malenka flexura (Plecoptera: Nemouridae) and Phylogenetic Analysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
2.2. Sequencing and Bioinformatics Analyses
2.3. Phylogenetic Analyses
3. Results and Discussion
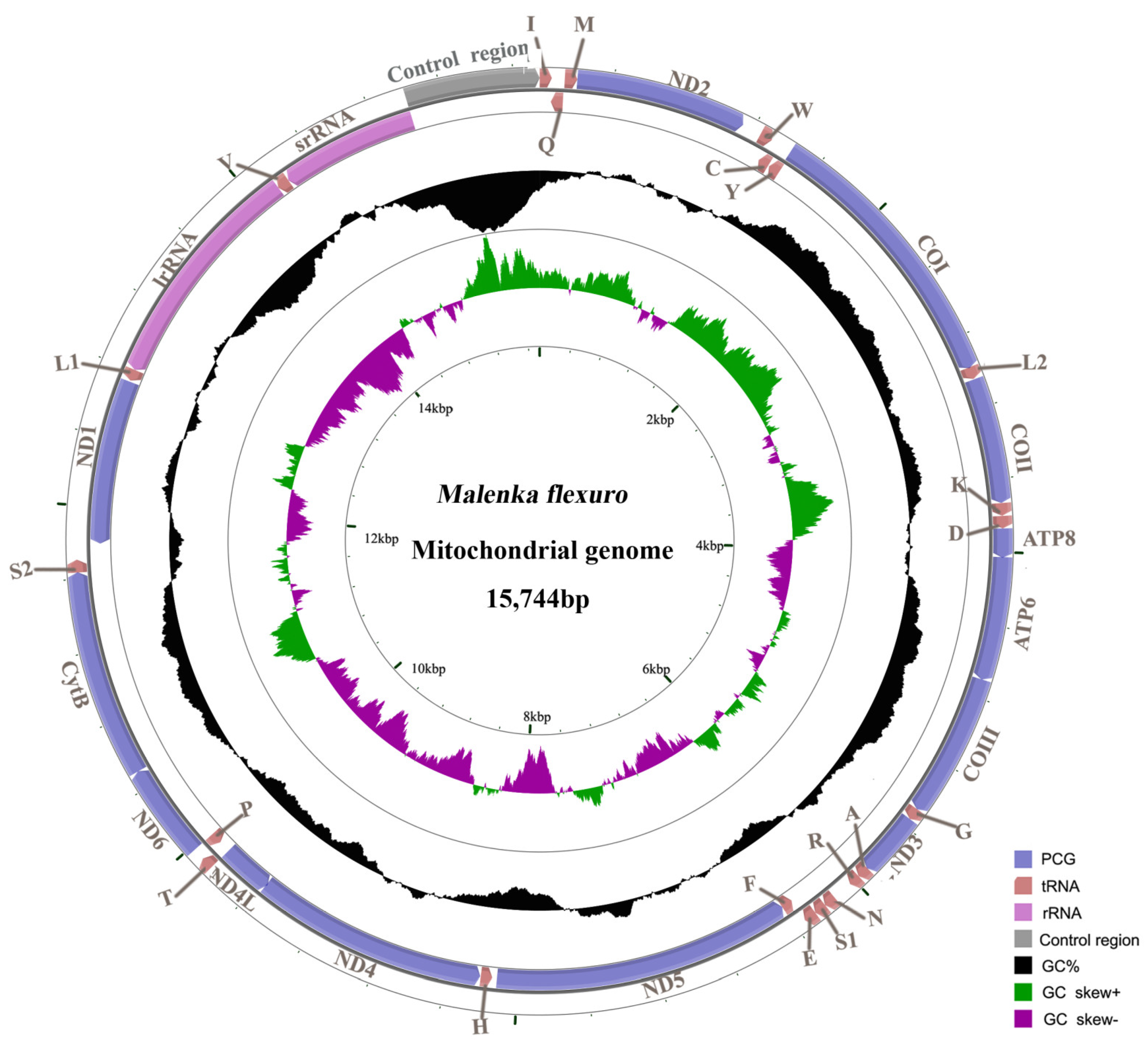
3.1. Mitogenome Organization and Base Composition
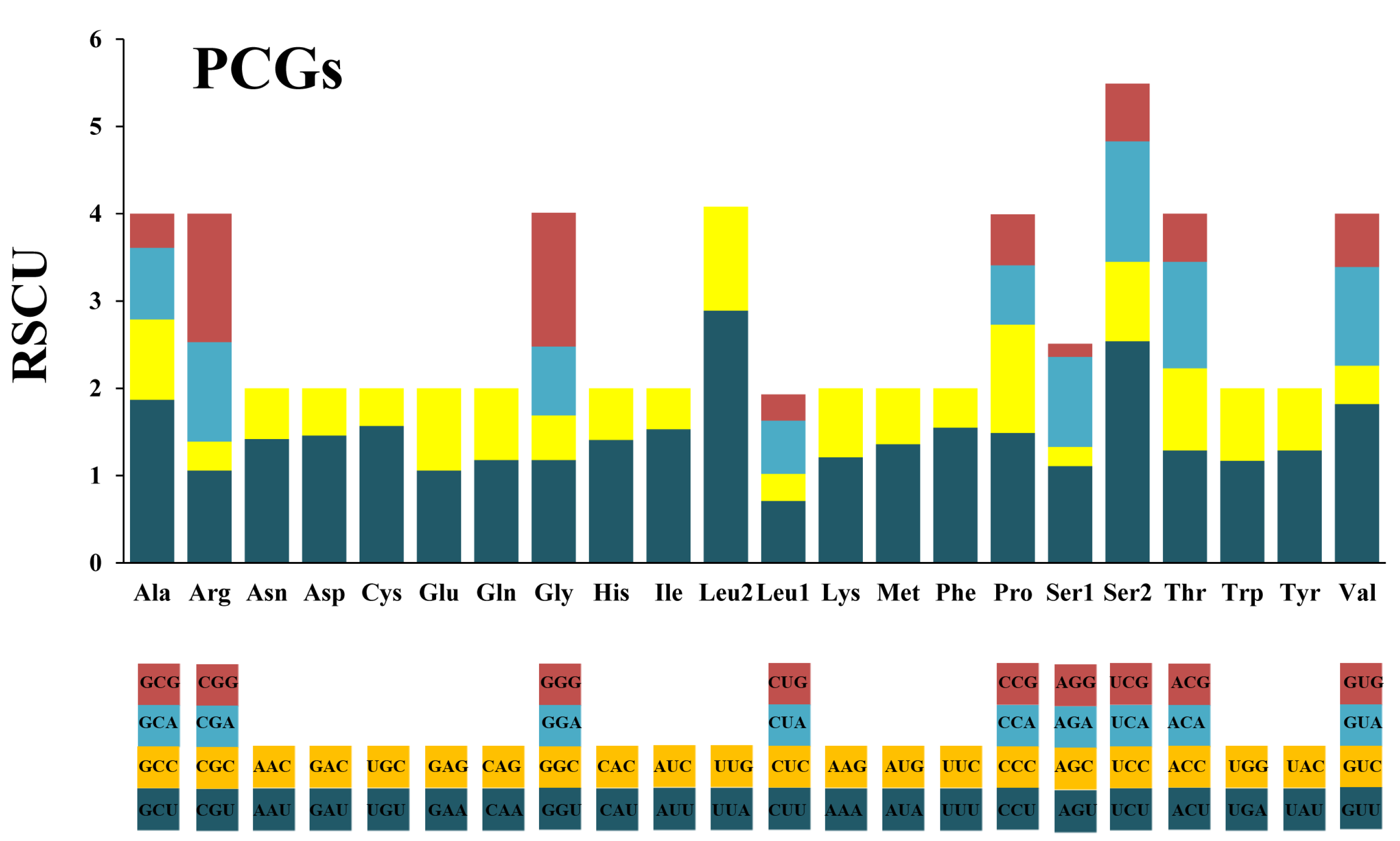
3.2. Protein-Coding Genes and Codon Usage
3.3. Transfer and Ribosomal RNA Genes
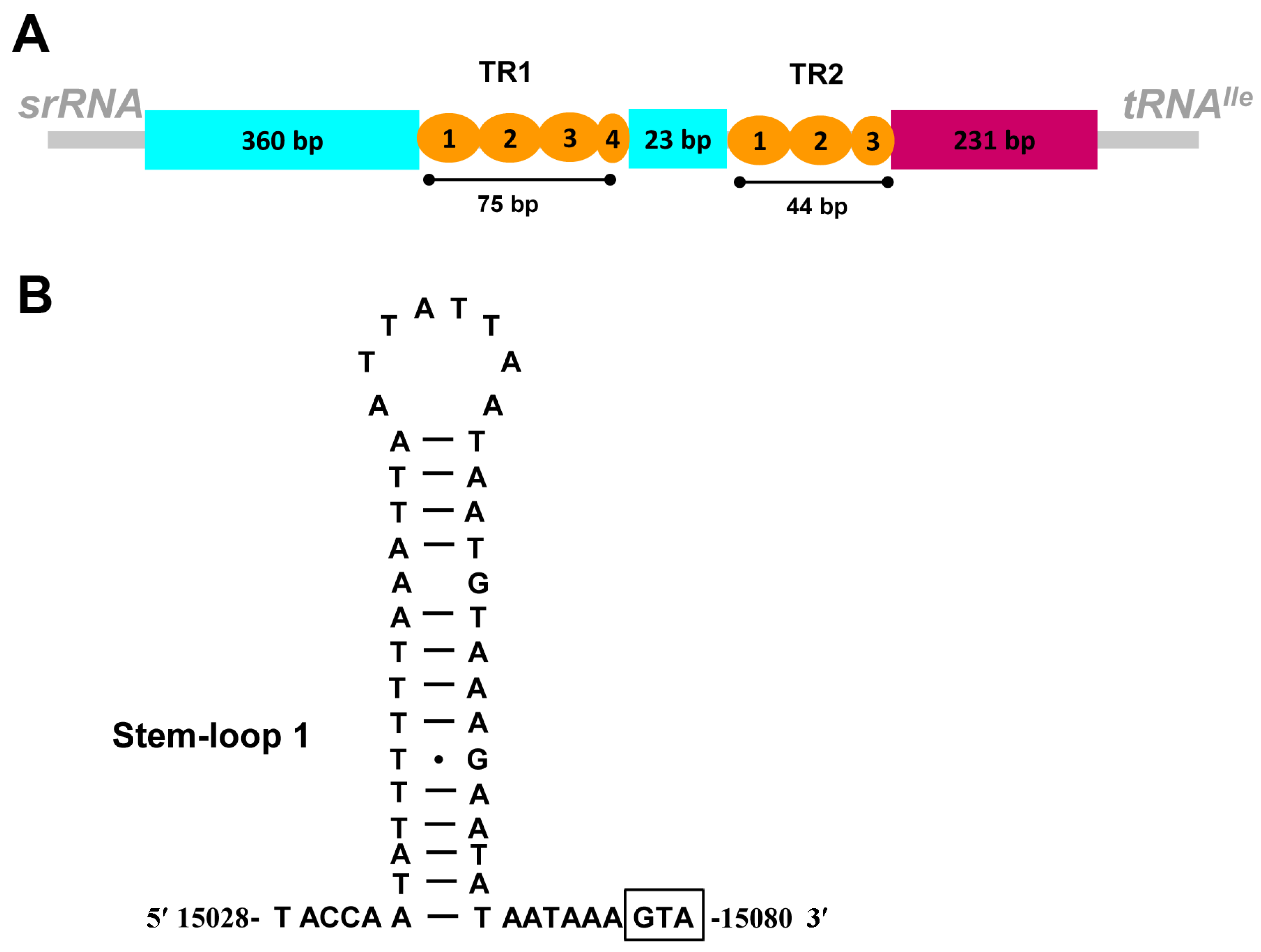
3.4. The Control Region
3.5. Phylogenetic Relationships
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, C.; Buckley, T.R.; Frati, F.; Stewart, J.B.; Beckenbach, A.T. Incorporating molecular evolution into phylogenetic analysis, and a new compilation of conserved polymerase chain reaction primers for animal mitochondrial DNA. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 545–579. [Google Scholar] [CrossRef] [Green Version]
- Wolstenholme, D. Genetic novelties in mitochondrial genomes of multicellular animals. Curr. Opin. Genet. Dev. 1992, 2, 918–925. [Google Scholar] [CrossRef]
- Cameron, S.L. Insect mitochondrial genomics: Implications for evolution and phylogeny. Annu. Rev. Èntomol. 2014, 59, 95–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Leavengood, J.M., Jr.; Chapman, E.G.; Burkhardt, D.; Song, F.; Jiang, P.; Liu, J.; Zhou, X.G.; Cai, W.Z. Mitochondrial phylogenomics of Hemiptera reveals adaptive innovations driving the diversification of true bugs. Proc. Roy. Soc. B–Biol. Sci. 2017, 284, 1862. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Li, H.; Jiang, P.; Zhou, X.; Liu, J.; Sun, C.; Vogler, A.P.; Cai, W. Capturing the phylogeny of holometabola with mitochondrial genome data and bayesian site-Heterogeneous mixture models. Genome Biol. Evol. 2016, 8, 1411–1426. [Google Scholar] [CrossRef]
- Song, F.; Li, H.; Liu, G.-H.; Wang, W.; James, P.; Colwell, D.D.; Tran, A.; Gong, S.; Cai, W.; Shao, R. Mitochondrial genome fragmentation unites the parasitic lice of Eutherian Mammals. Syst. Biol. 2019, 68, 430–440. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene-sequences and a compilation of conserved polymerase chain–reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Plecoptera Species File Online. Available online: http://Plecoptera.SpeciesFile.org (accessed on 10 April 2022).
- Zwick, P. Phylogenetic system and zoogeography of the Plecoptera. Annu. Rev. Èntomol. 2000, 45, 709–746. [Google Scholar] [CrossRef]
- South, E.J.; Skinner, R.K.; DeWalt, R.E.; A Davis, M.; Johnson, K.P.; A Teslenko, V.; Lee, J.J.; Malison, R.L.; Hwang, J.M.; Bae, Y.J.; et al. A new family of stoneflies (Insecta: Plecoptera), Kathroperlidae, fam. n., with a phylogenomic analysis of the Paraperlinae (Plecoptera: Chloroperlidae). Insect Syst. Divers. 2021, 5, 1. [Google Scholar] [CrossRef]
- Karr, J.R. Defining and measuring river health. Freshw. Biol. 1999, 41, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Baumann, R.W. Revision of the stonefly family Nemouridae (Plecoptera): A study of the world fauna at the generic level. Smithson. Contrib. Zoöl. 1975, 211, 1–74. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Sivec, I. Sphaeronemoura, a new genus of the Amphinemurinae (Nemouridae, Plecoptera) from asia. In Trends in Research in Ephemeroptera and Plecoptera; Dominguez, E., Ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2001; pp. 393–399. [Google Scholar]
- Sivec, I.; Stark, B.P. Tominemoura, a new stonefly genus from Sabah, East Malaysia (Plecoptera: Nemouridae). Illiesia 2009, 5, 199–203. [Google Scholar]
- Thomas, M.A.; Walsh, K.A.; Wolf, M.R.; McPheron, B.A.; Marden, J.H. Molecular phylogenetic analysis of evolutionary trends in stonefly wing structure and locomotor behavior. Proc. Natl. Acad. Sci. USA 2000, 97, 13178–13183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terry, M.D. Phylogeny of the polyneopterous insects with emphasis on Plecoptera: Molecular and morpological evidence. Ph.D. Thesis, Brigham Young University, Provo, UT, USA, 2003; pp. 1–118. [Google Scholar]
- Cao, J.-J.; Wang, Y.; Li, W.-H. Comparative mitogenomic analysis of species in the subfamily Amphinemurinae (Plecoptera: Nemouridae) reveal conserved mitochondrial genome organization. Int. J. Biol. Macromol. 2019, 138, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Wang, Y.; Guo, X.; Wang, G.; Li, W.; Murányi, D. Two complete mitochondrial genomes from Leuctridae (Plecoptera: Nemouroidea): Implications for the phylogenetic relationships among stoneflies. J. Insect Sci. 2021, 21, 16. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, J.; Murányi, D.; Chen, X.; Yan, F. The complete mitochondrial genome of Amphinemura bulla Shimizu, 1997 (Plecoptera: Nemouridae) from Japan. Mitochondrial DNA Part B 2021, 6, 846–847. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, J.; Li, W. The complete mitochondrial genome of the styloperlid stonefly species Styloperla spinicercia Wu (Insecta: Plecoptera) with family-level phylogenetic analyses of the Pteronarcyoidea. Zootaxa 2017, 4243, 125–138. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, J.; Murányi, D.; Li, W. Comparison of two complete mitochondrial genomes from Perlodidae (Plecoptera: Perloidea) and the family-level phylogenetic implications of Perloidea. Gene 2018, 675, 254–264. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, J.J.; Li, N.; Ma, G.-Y.; Li, W.-H. The first mitochondrial genome from Scopuridae (Insecta: Plecoptera) reveals structural features and phylogenetic implications. Int. J. Biol. Macromol. 2019, 122, 893–902. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenetics Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Abascal, F.; Zardoya, R.; Telford, M.J. TranslatorX: Multiple alignment of nucleotide sequences guided by amino acid translations. Nucleic Acids Res. 2010, 38, W7–W13. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQTREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Clary, D.O.; Wolstenholme, D.R. The ribosomal RNA genes of Drosophila mitochondrial DNA. Nucleic Acids Res. 1985, 13, 4029–4045. [Google Scholar] [CrossRef] [Green Version]
- Stewart, J.B.; Beckenbach, A.T. Insect mitochondrial genomics: The complete mitogenome sequence of the meadow spittlebug Philaenus spumarius (Hemiptera: Auchenorrhyncha: Cercopoidae). Genome 2005, 48, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Carapelli, A.; Vannini, L.; Nardi, F.; Boore, J.L.; Beani, L.; Dallai, R.; Frati, F. The mitochondrial genome of the entomophagous endoparasite Xenos vesparum (Insecta: Strepsiptera). Gene 2006, 376, 248–259. [Google Scholar] [CrossRef] [Green Version]
- Hassanin, A.; Léger, N.; Deutsch, J. Evidence for multiple reversals of asymmetric mutational constraints during the evolution of the mitochondrial genome of Metazoa, and consequences for phylogenetic inferences. Syst. Biol. 2005, 54, 277–298. [Google Scholar] [CrossRef] [PubMed]
- Hassanin, A. Phylogeny of Arthropoda inferred from mitochondrial sequences: Strategies for limiting the misleading effects of multiple changes in pattern and rates of substitution. Mol. Phylogenetics Evol. 2006, 38, 100–116. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.J.; Shi, M.; Chen, X.X.; Sharkey, M.J.; van Achterberg, C.; Ye, G.Y.; He, J.H. New views on strand asymmetry in insect mitochondrial genomes. PLoS ONE 2010, 5, e12708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.T.; Du, Y.Z. First Mitochondrial Genome from Nemouridae (Plecoptera) Reveals novel features of the elongated control region and phylogenetic implications. Int. J. Mol. Sci. 2017, 18, 996. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.Y.; Ji, X.Y.; Yu, W.W.; Du, Y.Z. Complete mitochondrial genome of the stonefly Cryptoperla stilifera Sivec (Plecop-tera: Peltoperlidae) and the phylogeny of Polyneopteran insects. Gene 2014, 537, 177–183. [Google Scholar] [CrossRef]
- Bae, J.S.; Kim, I.; Sohn, H.D.; Jin, B.R. The mitochondrial genome of the firefly, Pyrocoelia rufa: Complete DNA sequence, genome organization, and phylogenetic analysis with other insects. Mol. Phylogenetics Evol. 2004, 32, 978–985. [Google Scholar] [CrossRef]
- Sheffield, N.C.; Song, H.; Cameron, S.L.; Whiting, M.F. A Comparative analysis of mitochondrial genomes in Coleoptera (Arthropoda: Insecta) and genome descriptions of six new beetles. Mol. Biol. Evol. 2008, 25, 2499–2509. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Ding, S.; Yang, D. The complete mitochondrial genome of a stonefly species, Kamimuria chungnanshana Wu, 1948 (Plecoptera: Perlidae). Mitochondrial DNA Part A 2016, 27, 3810–3811. [Google Scholar] [CrossRef] [PubMed]
- Ojala, D.; Montoya, J.; Attardi, G. tRNA punctuation model of RNA processing in human mitochondria. Nature 1981, 290, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Boore, J.L. Complete mitochondrial genome sequence of Urechis caupo, a representative of the phylum Echiura. BMC Genom. 2004, 5, e67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, P.M.; Li, W.H. An evolutionary perspective on synonymous codon usage in unicellular organisms. J. Mol. Evol. 1986, 24, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Taanman, J.W. The mitochondrial genome: Structure, transcription, translation and replication. Biochim. Biophys. Acta BBA-Bioenergies 1999, 1410, 103–123. [Google Scholar] [CrossRef] [Green Version]
- Clary, D.O.; Wolstenholme, D.R. Drosophila mitochondrial DNA: Conserved sequences in the A+T–rich region and sup-porting evidence for a secondary structure model of the small ribosomal RNA. J. Mol. Evol. 1987, 25, 116–125. [Google Scholar] [CrossRef]
- Chen, Z.T.; Du, Y.Z. Comparison of the complete mitochondrial genome of the stonefly Sweltsa longistyla (Plecoptera: Chloroperlidae) with mitogenomes of three other stoneflies. Gene 2015, 558, 82–87. [Google Scholar] [CrossRef]
- Zhang, D.X.; Hewitt, G.M. Insect mitochondrial control region: A review of its structure, evolution and usefulness in evolutionary studies. Biochem. Syst. Ecol. 1997, 25, 99–120. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily | Species | Number (bp) | Accession Number |
|---|---|---|---|
| Amphinemurinae | Amphinemura longispina | 15,709 | MH085446 |
| Amphinemura yao | 15,876 | MH085447 | |
| Indonemoura auriformis | 15,718 | MN419915 | |
| Indonemoura jacobsoni | 15,642 | MH085448 | |
| Indonemoura nohirae | 15,738 | MH085449 | |
| Malenka flexura | 15,744 | ON411527 | |
| Mesonemoura metafiligera | 15,739 | MH085450 | |
| Mesonemoura tritaenia | 15,778 | MH085451 | |
| Protonemura datongensis | 15,756 | MT276842 | |
| Protonemura kohnoae | 15,707 | MH085452 | |
| Protonemura orbiculata | 15,758 | MH085453 | |
| Sphaeronemoura acutispina | 15,016 | MH085455 * | |
| Sphaeronemoura elephas | 15,846 | MN944385 | |
| Sphaeronemoura grandicauda | 15,661 | MH085454 * | |
| Nemourinae (Outgroup) | Nemoura meniscata | 15,895 | MN944386 |
| Nemoura nankinensis | 16,602 | KY940360 | |
| Nemoura papilla | 15,774 | MK290826 |
| Gene | Direction | Coordinates (bp) | Size (bp) | Anticodon or Start/Stop Codons | IGN (bp) |
|---|---|---|---|---|---|
| tRNAIle | J | 1–66 | 66 | 30–32 GAT | 0 |
| tRNAGln | N | 64–132 | 69 | 100–102 TTG | −3 |
| tRNAMet | J | 137–204 | 68 | 167–169 CAT | 4 |
| ND2 | J | 205–1239 | 1035 | ATG/TAA | 0 |
| tRNATrp | J | 1247–1315 | 69 | 1277–1279 TCA | 7 |
| tRNACys | N | 1308–1370 | 63 | 1339–1341 GCA | −8 |
| tRNATyr | N | 1377–1442 | 66 | 1409–1411 GTA | 6 |
| COI | J | 1435–2979 | 1545 | ATT/TAA | −8 |
| tRNALeu(UUR) | J | 2975–3041 | 67 | 3017–3019 TAA | −5 |
| COII | J | 3045–3732 | 688 | ATG/T- | 3 |
| tRNALys | J | 3733–3803 | 71 | 3763–3765 CTT | 0 |
| tRNAAsp | J | 3803–3870 | 68 | 3832–3834 GTC | −1 |
| ATP8 | J | 3871–4029 | 159 | ATT/TAA | 0 |
| ATP6 | J | 4023–4700 | 678 | ATG/TAA | −7 |
| COIII | J | 4700–5488 | 789 | ATG/TAA | −1 |
| tRNAGly | J | 5488–5553 | 66 | 5517–5519 TCC | −1 |
| ND3 | J | 5554–5907 | 354 | ATT/TAG | 3 |
| tRNAAla | J | 5906–5969 | 64 | 5935–5937 TGC | −2 |
| tRNAArg | J | 5970–6032 | 63 | 5999–6001 TCG | 0 |
| tRNAAsn | J | 6144–6209 | 66 | 6174–6172 GTT | 111 |
| tRNASer(AGN) | J | 6209–6277 | 69 | 6235–6237GCT | −1 |
| tRNAGlu | J | 6277–6345 | 69 | 6307–6309 TTC | −1 |
| tRNAPhe | N | 6344–6408 | 65 | 6376–6378 GAA | −2 |
| ND5 | N | 6409–8143 | 1735 | GTG/T- | 0 |
| tRNAHis | N | 8144–8209 | 66 | 8177–8179 GTG | 0 |
| ND4 | N | 8213–9553 | 1341 | ATG/TAA | 3 |
| ND4L | N | 9547–9843 | 297 | ATG/TAA | −7 |
| tRNAThr | J | 9846–9911 | 66 | 9877–9879 TGT | 2 |
| tRNAPro | N | 9911–9975 | 65 | 9943–9945 TGG | −1 |
| ND6 | J | 9977–10,501 | 525 | ATT/TAA | 0 |
| CytB | J | 10,501–11,637 | 1137 | ATG/TAG | −1 |
| tRNASer(UCN) | J | 11,636–11,705 | 70 | 11,667–11,669 TGA | −2 |
| ND1 | N | 11,792–12,742 | 951 | TTG/TAG | 86 |
| tRNALeu(CUN) | N | 12,744–12,809 | 66 | 12,778–12,780TAG | 1 |
| lrRNA | N | 12,810–14,148 | 1339 | 0 | |
| tRNAVal | N | 14,149–14,219 | 71 | 14,184–14,1186 TAC | 0 |
| srRNA | N | 14,220–15,009 | 790 | 0 | |
| CR | 15,010–15,744 | 735 | 0 |
| Feature | Proportion of Nucleotides (%) | AT Skew | GC Skew | ||||
|---|---|---|---|---|---|---|---|
| T | C | A | G | A + T | |||
| Whole mitogenome | 36.3 | 12.5 | 32.3 | 18.8 | 68.6 | −0.059 | 0.200 |
| Protein-coding genes | 39.3 | 17.1 | 27.3 | 16.3 | 66.6 | −0.180 | −0.022 |
| Protein-coding genes J-strand | 35.6 | 20.7 | 29.1 | 14.7 | 64.7 | −0.101 | −0.171 |
| Protein-coding genes N-strand | 45.1 | 11.4 | 24.4 | 19.1 | 69.6 | −0.297 | 0.2552 |
| tRNA genes | 35.5 | 12.2 | 35.4 | 16.9 | 70.9 | −0.001 | 0.160 |
| tRNA genes J-strand | 34.9 | 13.9 | 36.0 | 15.2 | 70.9 | 0.015 | 0.044 |
| tRNA genes N-strand | 36.5 | 9.2 | 34.5 | 19.9 | 71.0 | −0.029 | 0.368 |
| rRNA genes | 39.0 | 10.1 | 32.9 | 18.1 | 71.9 | −0.084 | 0.285 |
| lrRNA | 40.4 | 9.0 | 33.2 | 17.4 | 73.6 | −0.097 | 0.320 |
| srRNA | 36.5 | 12.0 | 32.3 | 19.2 | 68.9 | −0.061 | 0.231 |
| Control region | 43.6 | 8.1 | 41.6 | 6.7 | 85.2 | −0.023 | −0.100 |
| Codon | Count | Codon | Count | Codon | Count | Codon | Count |
|---|---|---|---|---|---|---|---|
| UUU(F) | 270 | UCU(S) | 95 | UAU(Y) | 109 | UGU(C) | 34 |
| UUC(F) | 51 | UCC(S) | 31 | UAC(Y) | 48 | UGC(C) | 8 |
| UUA(L) | 364 | UCA(S) | 74 | UAA(*) | 0 | UGA(W) | 87 |
| UUG(L) | 46 | UCG(S) | 12 | UAG(*) | 0 | UGG(W) | 19 |
| CUU(L) | 102 | CCU(P) | 63 | CAU(H) | 58 | CGU(R) | 14 |
| CUC(L) | 31 | CCC(P) | 38 | CAC(H) | 25 | CGC(R) | 7 |
| CUA(L) | 71 | CCA(P) | 42 | CAA(Q) | 64 | CGA(R) | 32 |
| CUG(L) | 12 | CCG(P) | 9 | CAG(Q) | 117 | CGG(R) | 7 |
| AUU(I) | 271 | ACU(T) | 87 | AAU(N) | 127 | AGU(S) | 45 |
| AUC(I) | 41 | ACC(T) | 27 | AAC(N) | 23 | AGC(S) | 17 |
| AUA(I) | 153 | ACA(T) | 75 | AAA(K) | 52 | AGA(S) | 63 |
| AUG(M) | 38 | ACG(T) | 12 | AAG(K) | 20 | AGG(S) | 3 |
| GUU(V) | 105 | GCU(A) | 92 | GAU(D) | 52 | GGU(G) | 53 |
| GUC(V) | 28 | GCC(A) | 38 | GAC(D) | 18 | GGC(G) | 34 |
| GUA(V) | 77 | GCA(A) | 54 | GAA(E) | 55 | GGA(G) | 83 |
| GUG(V) | 23 | GCG(A) | 24 | GAG(E) | 24 | GGG(G) | 79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, J.; Guo, X.; Guo, C.; Wang, X.; Wang, Y.; Yan, F. Complete Mitochondrial Genome of Malenka flexura (Plecoptera: Nemouridae) and Phylogenetic Analysis. Genes 2022, 13, 911. https://doi.org/10.3390/genes13050911
Cao J, Guo X, Guo C, Wang X, Wang Y, Yan F. Complete Mitochondrial Genome of Malenka flexura (Plecoptera: Nemouridae) and Phylogenetic Analysis. Genes. 2022; 13(5):911. https://doi.org/10.3390/genes13050911
Chicago/Turabian StyleCao, Jinjun, Xuan Guo, Caiyue Guo, Xuan Wang, Ying Wang, and Fengming Yan. 2022. "Complete Mitochondrial Genome of Malenka flexura (Plecoptera: Nemouridae) and Phylogenetic Analysis" Genes 13, no. 5: 911. https://doi.org/10.3390/genes13050911
APA StyleCao, J., Guo, X., Guo, C., Wang, X., Wang, Y., & Yan, F. (2022). Complete Mitochondrial Genome of Malenka flexura (Plecoptera: Nemouridae) and Phylogenetic Analysis. Genes, 13(5), 911. https://doi.org/10.3390/genes13050911