

Circulating microRNAs in Hidradenitis Suppurativa

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Recruitment of Patients
2.2. RNA Extraction
2.3. Small RNA Library Preparation and Sequencing
2.4. Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
2.5. MiRNA Target Prediction
3. Results
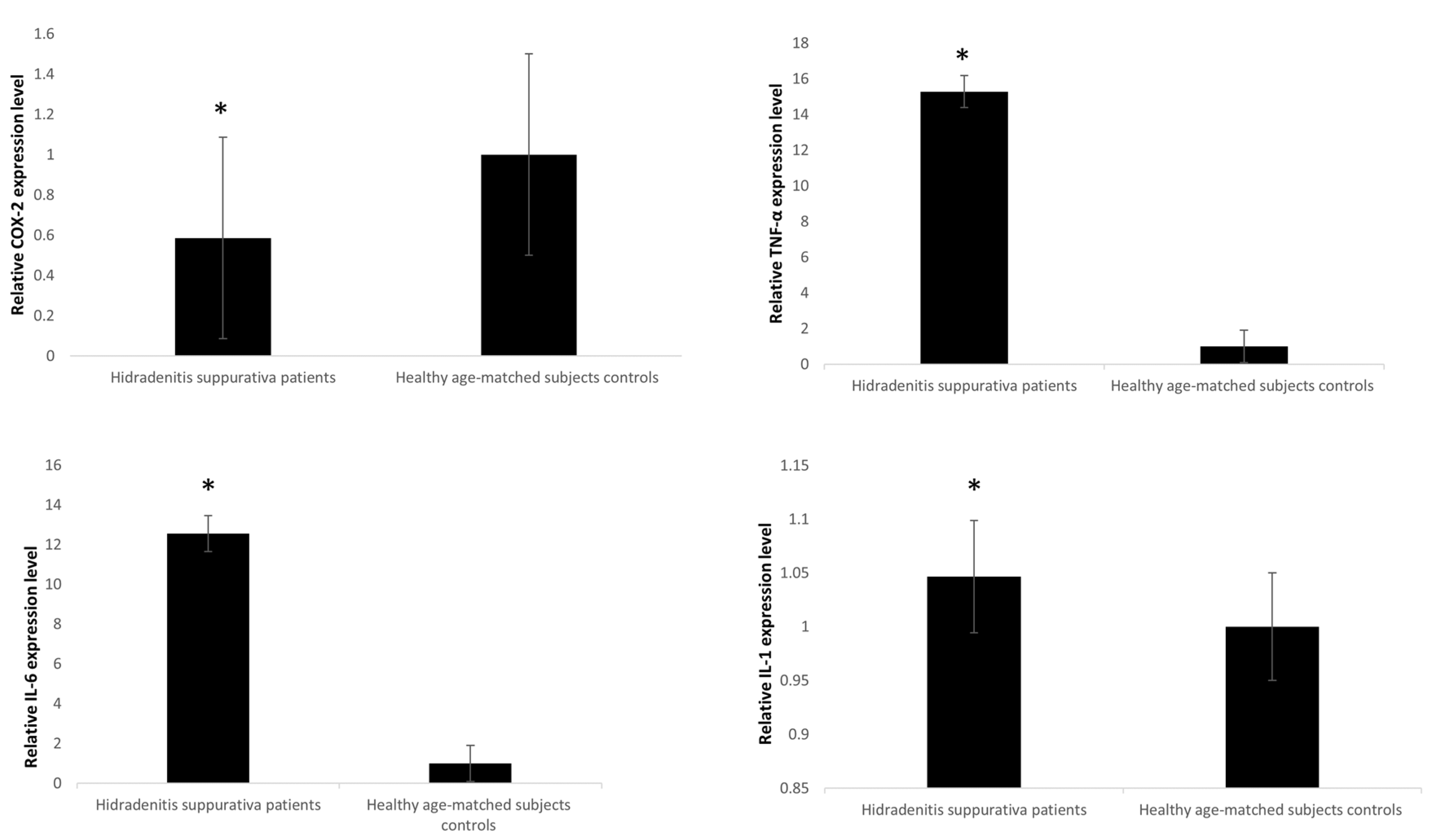
3.1. Anthropometric and Laboratory Investigations
3.2. qRTPCR microRNA Expression
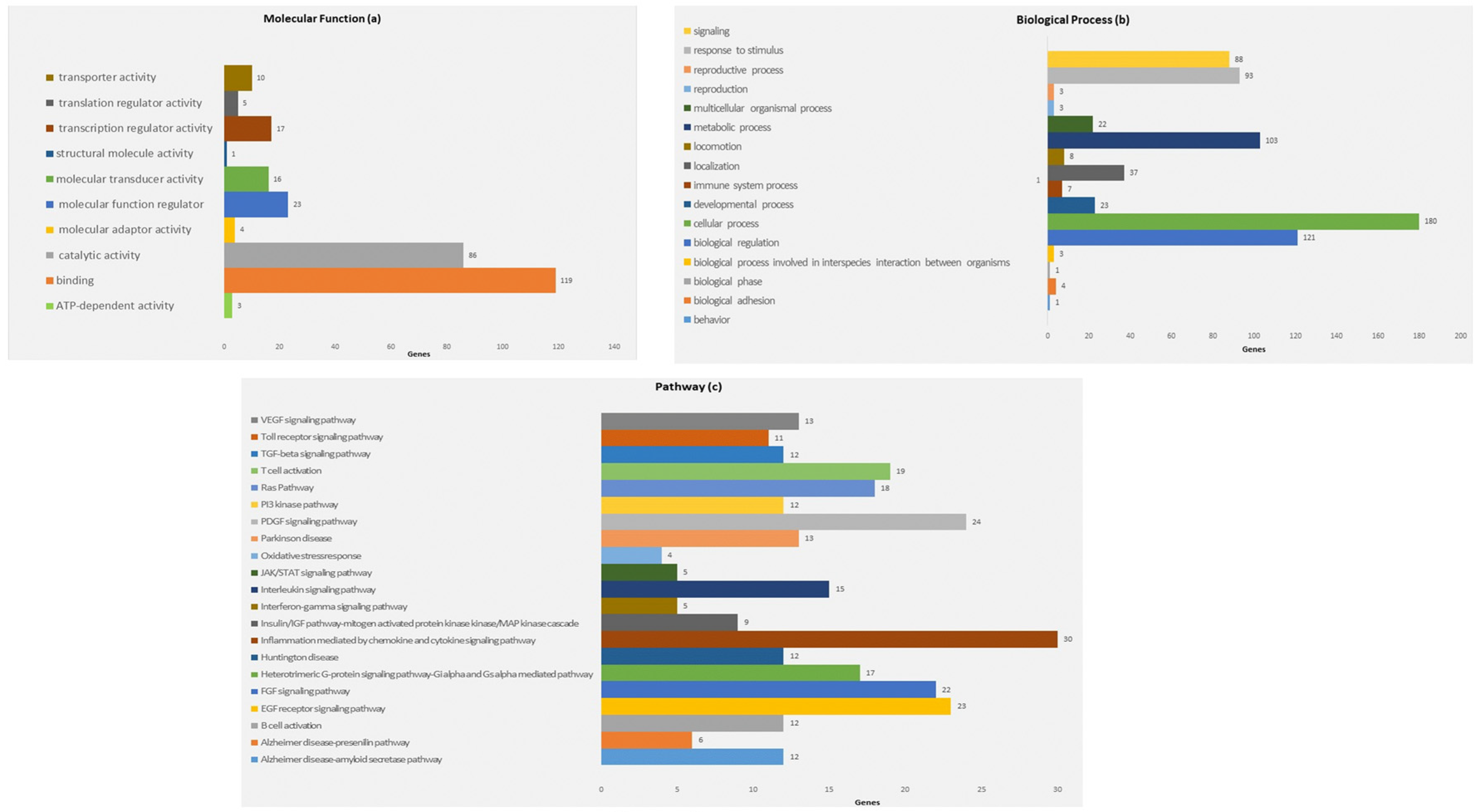
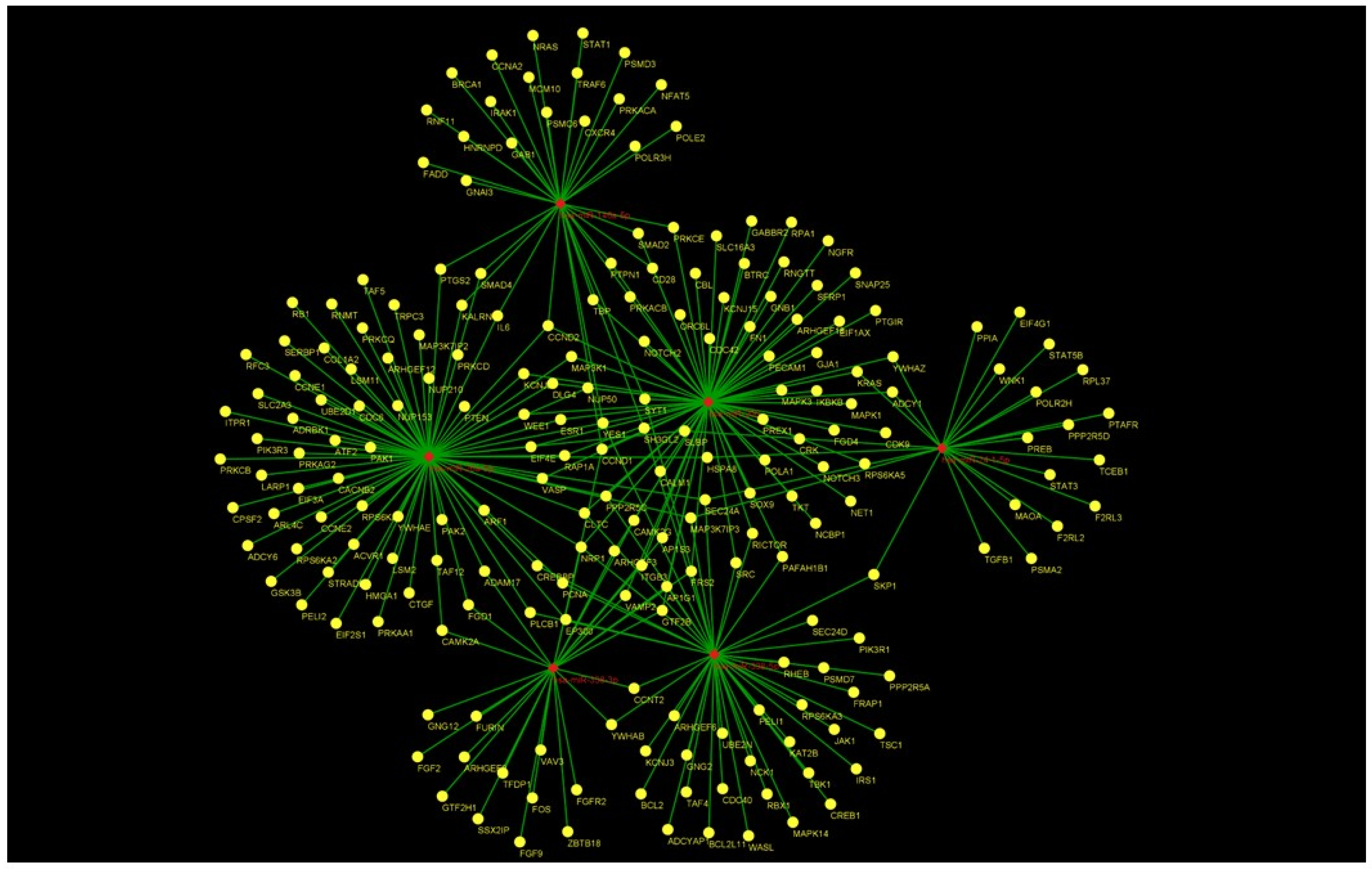
3.3. Computational Predictions of the Putative miRNAs Target
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coates, M.; Mariottoni, P.; Corcoran, D.L.; Kirshner, H.F.; Jaleel, T.; Brown, D.A.; Brooks, S.R.; Murray, J.; Morasso, M.I.; MacLeod, A.S. The skin transcriptome in hidradenitis suppurativa uncovers an antimicrobial and sweat gland gene signature which has distinct overlap with wounded skin. PLoS ONE 2019, 14, e0216249. [Google Scholar] [CrossRef] [PubMed]
- Miller, I.M.; McAndrew, R.J.; Hamzavi, I.H. Prevalence, risk factors, and comorbidities of hidradenitis suppurativa. Dermatol. Clin. 2016, 34, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Pink, A.E.; Simpson, M.A.; Desai, N.; Dafou, D.; Hills, A.; Mortimer, P.; Smith, C.H.; Trembath, R.C.; Barker, J.N. Mutations in the gamma-secretase genes NCSTN, PSENEN, and PSEN1 underlie rare forms of hidradenitis suppurativa (acne inversa). J. Investig. Dermatol. 2012, 132, 2459–2461. [Google Scholar] [CrossRef] [PubMed]
- Pink, A.E.; Simpson, M.A.; Desai, N.; Trembath, R.C.; Barker, J.N. Gamma-secretase mutations in hidradenitis suppurativa: New insights into disease pathogenesis. J. Investig. Dermatol. 2013, 133, 601–607. [Google Scholar] [CrossRef]
- Garg, A.; Lavian, J.; Lin, G.; Strunk, A.; Alloo, A. Incidence of hidradenitis suppurativa in the United States: A sex- and age-adjusted population analysis. J. Am. Acad. Dermatol. 2017, 77, 118–122. [Google Scholar] [CrossRef]
- van der Zee, H.H.; de Ruiter, L.; van den Broecke, D.G.; Dik, W.A.; Laman, J.D.; Prens, E.P. Elevated levels of tumour necrosis factor (TNF)-a, interleukin (IL)-1b and IL-10 in hidradenitis suppurativa skin: A rationale for targeting TNFa and IL-1b. Br. J. Dermatol. 2011, 164, 1292–1298. [Google Scholar] [CrossRef]
- Egeberg, A.; Gislason, G.H.; Hansen, P.R. Risk of major adverse cardiovascular events and all-cause mortality in patients with hidradenitis suppurativa. JAMA Dermatol. 2016, 152, 429–434. [Google Scholar] [CrossRef]
- Richette, P.; Molto, A.; Viguier, M.; Dawidowicz, K.; Hayem, G.; Nassif, A.; Wendling, D.; Aubin, F.; Lioté, F.; Bachelez, H. Hidradenitis suppurativa associated with spondyloarthritis—Results from a multicenter national prospective study. J. Rheumatol. 2014, 41, 490–494. [Google Scholar] [CrossRef]
- Van der Zee, H.H.; de Winter, K.; van der Woude, C.J.; Prens, E.P. The prevalence of hidradenitis suppurativa in 1093 patients with inflammatory bowel disease. Br. J. Dermatol. 2014, 171, 673–675. [Google Scholar]
- Schrader, A.M.; Deckers, I.E.; van der Zee, H.H.; Boer, J.; Prens, E.P. Hidradenitis suppurativa: A retrospective study of 846 Dutch patients to identify factors associated with disease severity. J. Am. Acad. Dermatol. 2014, 71, 460–467. [Google Scholar] [CrossRef]
- Gold, D.A.; Reeder, V.J.; Mahan, M.G.; Hamzavi, I.H. The prevalence of metabolic syndrome in patients with hidradenitis suppurativa. J. Am. Acad. Dermatol. 2014, 70, 699–703. [Google Scholar] [CrossRef] [PubMed]
- Sabat, R.; Chanwangpong, A.; Schneider-Burrus, S.; Metternich, D.; Kokolakis, G.; Kurek, A.; Philipp, S.; Uribe, D.; Wolk, K.; Sterry, W. Increased prevalence of metabolic syndrome in patients with acne inversa. PLoS ONE 2012, 7, e31810. [Google Scholar] [CrossRef] [PubMed]
- Hausser, J.; Zavolan, M. Identification and consequences of miRNA-target interactions—Beyond repression of gene expression. Nat. Rev. Genet. 2014, 15, 599–612. [Google Scholar] [CrossRef]
- Gurtan, A.M.; Sharp, P.A. The role of miRNAs in regulating gene expression networks. J. Mol. Biol. 2013, 425, 3582–3600. [Google Scholar] [CrossRef] [PubMed]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011, 39, 7223–7233. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.M.; Reeves, G.; Billman, G.E.; Sturmberg, J.P. Inflammation-Nature’s Way to Efficiently Respond to All Types of Challenges: Implications for Understanding and Managing “the Epidemic” of Chronic Diseases. Front. Med. 2018, 5, 316. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.M.; Rao, D.S.; Chaudhuri, A.A.; Baltimore, D. Physiological and pathological roles for microRNAs in the immune system. Nat. Rev. Immunol. 2010, 10, 111–122. [Google Scholar] [CrossRef]
- Ro, S.; Yan, W. Small RNA cloning. Methods Mol. Biol. 2010, 629, 273–285. [Google Scholar] [CrossRef]
- Reuter, J.S.; Mathews, D.H. RNAstructure: Software for RNA secondary structure prediction and analysis. BMC Bioinform. 2010, 11, 129. [Google Scholar] [CrossRef]
- Ward, J.R.; Heath, P.R.; Catto, J.W.; Whyte, M.K.B.; Milo, M.; Renshaw, S.A. Regulation of neutrophil senescence by microRNAs. PLoS ONE 2011, 6, e15810. [Google Scholar] [CrossRef] [PubMed]
- Pace, N.P.; Mintoff, D.; Borg, I. The Genomic Architecture of Hidradenitis Suppurativa—A Systematic Review. Front. Genet. 2022, 13, 861241. [Google Scholar] [CrossRef] [PubMed]
- Der Sarkissian, S.; Hessam, S.; Kirby, J.S.; Lowes, M.A.; Mintoff, D.; Naik, H.B.; Ring, H.C.; Chandran, N.S.; Frew, J.W. Identification of Biomarkers and Critical Evaluation of Biomarker Validation in Hidradenitis Suppurativa: A Systematic Review. JAMA Dermatol. 2022, 158, 300–313. [Google Scholar] [CrossRef] [PubMed]
- Hessam, S.; Sand, M.; Skrygan, M.; Gambichler, T.; Bechara, F.G. Expression of miRNA-155, miRNA-223, miRNA-31, miRNA-21, miRNA-125b, and miRNA-146a in the Inflammatory Pathway of Hidradenitis Suppurativa. Inflammation 2016, 40, 464–472. [Google Scholar] [CrossRef]
- Heyam, A.; Lagos, D.; Plevin, M. Dissecting the roles of TRBP and PACT in double-stranded RNA recognition and processing of noncoding RNAs. Wiley Interdiscip. Rev. RNA 2015, 6, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, F.; Prattichizzo, F.; Giuliani, A.; Matacchione, G.; Rippo, M.R.; Sabbatinelli, J.; Bonafè, M. miR-21 and miR-146a: The microRNAs of inflammaging and age-related diseases. Ageing Res. Rev. 2021, 70, 101374. [Google Scholar] [CrossRef]
- Luly, F.R.; Lévêque, M.; Licursi, V.; Cimino, G.; Martin-Chouly, C.; Théret, N.; Negri, R.; Cavinato, L.; Ascenzioni, F.; Del Porto, P. MiR-146a is over-expressed and controls IL-6 production in cystic fibrosis macrophages. Sci. Rep. 2019, 9, 16259. [Google Scholar] [CrossRef]
- Barutta, F.; Corbetta, B.; Bellini, S.; Guarrera, S.; Matullo, G.; Scandella, M.; Schalkwijk, C.; Stehouwer, C.D.; Chaturvedi, N.; Soedamah-Muthu, S.S.; et al. MicroRNA 146a is associated with diabetic complications in type 1 diabetic patients from the EURODIAB PCS. J. Transl. Med. 2021, 19, 475. [Google Scholar] [CrossRef]
- Giuliani, A.; Lattanzi, S.; Ramini, D.; Graciotti, L.; Danni, M.C.; Procopio, A.D.; Silvestrini, M.; Olivieri, F.; Sabbatinelli, J. Potential prognostic value of circulating inflamma-miR-146a-5p and miR-125a-5p in relapsing-remitting multiple sclerosis. Mult. Scler. Relat. Disord. 2021, 54, 103126. [Google Scholar] [CrossRef]
- Zhang, L.L.; Wu, X.X.; Wang, X.F.; Di, D.S.; Huang, Q.; Liu, R.S.; Shuai, Z.W.; Ye, D.Q.; Leng, R.X. Genetic variant in microRNA-146a gene is associated with risk of rheumatoid arthritis. Ann. Med. 2021, 53, 824–829. [Google Scholar] [CrossRef]
- Long, J.P.; Dong, L.F.; Chen, F.F.; Fan, Y.F. miR-146a-5p targets interleukin-1 receptor-associated kinase 1 to inhibit the growth, migration, and invasion of breast cancer cells. Oncol. Lett. 2019, 17, 1573–1580. [Google Scholar] [CrossRef]
- Zhan-Qiang, H.; Hai-Hua, Q.; Chi, Z.; Miao, W.; Cui, Z.; Zi-Yin, L.; Jing, H.; Yi-Wei, W. miR-146a aggravates cognitive impairment and Alzheimer disease-like pathology by triggering oxidative stress through MAPK signaling. Neurologia 2021. [Google Scholar] [CrossRef] [PubMed]
- Maffioletti, E.; Milanesi, E.; Ansari, A.; Zanetti, O.; Galluzzi, S.; Geroldi, C.; Gennarelli, M.; Bocchio-Chiavetto, L. miR-146a Plasma Levels Are Not Altered in Alzheimer’s Disease but Correlate with Age and Illness Severity. Front. Aging Neurosci. 2020, 11, 366. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, Y.; Liang, Q.; Yao, L.; Gu, S.; Bai, X. MiR-338-5p Promotes Inflammatory Response of Fibroblast-Like Synoviocytes in Rheumatoid Arthritis via Targeting SPRY1. J. Cell. Biochem. 2017, 118, 2295–2301. [Google Scholar] [CrossRef] [PubMed]
- Xing, Z.; Yu, L.; Li, X.; Su, X. Anticancer bioactive peptide-3 inhibits human gastric cancer growth by targeting miR-338-5p. Cell Biosci. 2016, 6, 53. [Google Scholar] [CrossRef]
- Zhuang, Y.; Dai, J.; Wang, Y.; Zhang, H.; Li, X.; Wang, C.; Cao, M.; Liu, Y.; Cai, H.; Zhang, D.; et al. MiR-338_suppresses fibrotic pathogenesis in pulmonary fibrosis through targeting L PA1. Am. J. Transl. Res. 2016, 8, 3197–3205. [Google Scholar]
- Chen, X.; Wei, L.; Zhao, S. MiR-338 inhibits the metastasis of lung cancer by targeting integrin beta3. Oncol. Rep. 2016, 36, 1467–1474. [Google Scholar] [CrossRef]
- Besse, A.; Sana, J.; Lakomy, R.; Kren, L.; Fadrus, P.; Smrcka, M.; Hermanova, M.; Jancalek, R.; Reguli, S.; Lipina, R.; et al. MiR-338-5p sensitizes glioblastoma cells to radiation through regulation of genes involved in DNA damage response. Tumor Biol. 2016, 37, 7719–7727. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HS Patients (n. 25) | Controls (n. 12) | p-Value | |
|---|---|---|---|
| Sex n. F/M (%) | 19/6 (76) | 5/7 (42) | |
| Age mean (SD) | 25.71 ± 13.31 | 33.25 ± 11.26 | 0.06 |
| BMI mean (SD) | 28.11± 6.19 | 25.72 ± 2.98 | 0.23 |
| Smokers’ status n. (%) | 51% | - | |
| Cholesterol (mg/dL) | 205.98 ± 38.57 | 155.72 ± 35.29 | 0.003 |
| Triglycerides (mg/dL) | 90.97 ± 27.16 | 85.63 ± 38.35 | 0.69 |
| Glycemia (mg/dL) | 96.57 ± 22.53 | 84.42 ± 5.38 | 0.18 |
| C-Reactive Protein (mg/L) | 10.57 ± 12.52 | - | |
| Hurley n. (%) | 15 (60) | - | |
| Hurley II n. (%) | 8 (32) | - | |
| Hurley III n. (%) | 2 (8) | - |
| Panther Pathway | p-Value | Molecules | miRNAs |
|---|---|---|---|
| Alzheimer’s disease–amyloid secretase pathway | 4.40 × 1011 | MAPK1, PRKACA, FURIN, ADAM17, PRKCE, PRKCQ, CACNB2, MAPK3, PRKCD, MAPK14, PAK1, PRKCB | hsa-miR-206, hsa-miR-146a-5p, hsa-miR-338-3p, hsa-miR-26a-5p, hsa-miR-338-5p |
| Alzheimer’s disease–presenilin pathway | 3.55 × 103 | FURIN, NOTCH2, ADAM17, NOTCH3, TRPC3, GSK3B | hsa-miR-338-3p, hsa-miR-206, hsa-miR-26a-5p |
| Axon guidance mediated by Slit/Robo | 3.22 × 103 | CXCR4, NET1, CDC42 | hsa-miR-146a-5p, hsa-miR-206, |
| Axon guidance mediated by netrin | 6.41 × 105 | NET1, PIK3R1, PIK3R3, VASP, CDC42, | hsa-miR-206, hsa-miR-338-5p, hsa-miR-26a-5p, |
| Axon guidance mediated by semaphorins | 2.47 × 102 | NRP1, PAK1 | hsa-miR-26a-5p, hsa-miR-338-3p, hsa-miR-338-5p, hsa-miR-206 |
| FAS signaling pathway | 3.13 × 10 | FADD | hsa-miR-146a-5p |
| Inflammation mediated by chemokine and cytokine signaling pathway | 5.72 × 1021 | PREX1, CAMK2G, CXCR4, ITPR1, CAMK2A, MAPK1, PTEN, NFAT5, PAK2, PRKCE, PRKACB, PTGS2, MAPK3, GNG2, IKBKB, PTAFR, ADRBK1, STAT3, GNG12, CDC42, PLCB1, PRKACA, STAT1, GNAI3, KRAS, NRAS, IL6, PAK1, ADCY6, PRKCB, | hsa-miR-206, hsa-miR-338-3p, hsa-miR-146a-5p, hsa-miR-26a-5p, hsa-miR-338-5p, hsa-miR-24-1-5p |
| Insulin/IGF pathway-mitogen activated protein kinase kinase/MAP kinase cascade | 5.00 × 10 | MAPK1, RPS6KA3, MAPK3, PTGIR, RPS6KA6, RPS6KA5, IRS1, RPS6KA2, FOS | hsa-miR-338-5p, hsa-miR-206, hsa-miR-26a-5p, hsa-miR-338-3p |
| Insulin/IGF pathway-protein kinase B signaling cascade | 9.03 × 106 | TSC1, PTEN, PIK3R1, PIK3R3, IRS1, GSK3B | hsa-miR-338-5p, hsa-miR-26a-5p |
| Interferon-γ signaling pathway | 3.30 × 105 | MAPK1, JAK1, MAPK3, MAPK14, STAT1 | hsa-miR-206, hsa-miR-338-5p, hsa-miR-146a-5p |
| Interleukin signaling pathway | 3.38 × 1013 | MAPK1, RPS6KA3, MAPK3, RPS6KA6, FRAP1, IKBKB, STAT3, STAT1, STAT5B, IRS1, RPS6KA2, FOS, NRAS, IL6, GSK3B | hsa-miR-206, hsa-miR-338-5p, hsa-miR-26a-5p, hsa-miR-24-1-5p, hsa-miR-146a-5p, hsa-miR-338-3p |
| Oxidative stress response | 3.85 × 103 | ATF2, MAPK14, STAT1, BCL2 | hsa-miR-26a-5p, hsa-miR-338-5p, hsa-miR-146a-5p |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Felice, B.; Montanino, C.; Mallardo, M.; Babino, G.; Mattera, E.; Ragozzino, G.; Argenziano, G.; Daniele, A.; Nigro, E. Circulating microRNAs in Hidradenitis Suppurativa. Genes 2022, 13, 1544. https://doi.org/10.3390/genes13091544
De Felice B, Montanino C, Mallardo M, Babino G, Mattera E, Ragozzino G, Argenziano G, Daniele A, Nigro E. Circulating microRNAs in Hidradenitis Suppurativa. Genes. 2022; 13(9):1544. https://doi.org/10.3390/genes13091544
Chicago/Turabian StyleDe Felice, Bruna, Concetta Montanino, Marta Mallardo, Graziella Babino, Edi Mattera, Giovanni Ragozzino, Giuseppe Argenziano, Aurora Daniele, and Ersilia Nigro. 2022. "Circulating microRNAs in Hidradenitis Suppurativa" Genes 13, no. 9: 1544. https://doi.org/10.3390/genes13091544
APA StyleDe Felice, B., Montanino, C., Mallardo, M., Babino, G., Mattera, E., Ragozzino, G., Argenziano, G., Daniele, A., & Nigro, E. (2022). Circulating microRNAs in Hidradenitis Suppurativa. Genes, 13(9), 1544. https://doi.org/10.3390/genes13091544